



Introduction
Walcottaspis vanhornei (Walcott, Reference Walcott1914) is a large but little-known dikelocephalid trilobite confined to a narrow outcrop belt of the St. Lawrence Formation in the Upper Mississippi Valley. Only four examples of any of its sclerites have previously been illustrated (Walcott, Reference Walcott1914, pl. 62, figs. 1–3; Ulrich and Resser, Reference Ulrich and Resser1930, pl. 20, figs. 3–5). This paper describes its morphology, ontogeny, and systematics and considers aspects of its paleoecology. It complements a forthcoming study of evolutionary patterns among dikelocephalid trilobites from this region. Study of this group is potentially instructive from an evolutionary perspective because the region contains a thin but relatively complete (Runkel, Reference Runkel1994; Runkel et al., Reference Runkel, Miller, McKay, Palmer and Taylor2007) record of late Cambrian depositional history formed close to the paleoshoreline. During that time, a condensed suite of nearshore facies was deposited in the Hollandale embayment (Mossler, Reference Mossler1992) within the northern part of the Mississippi Valley. The Hollandale embayment reflects an ancient rift basin located between the Wisconsin arch to the east and the putative “Transcontinental Arch” (Myrow et al., Reference Myrow, Taylor, Miller, Ethington, Ripperdan and Allen2003) to the west, two regions of positive relief that were reportedly intermittently exposed during the late Cambrian. This onshore environmental setting, comparable in some ways to modern coastal environments, may have offered opportunity for localized habitat fragmentation and thus enhanced opportunity for speciation, as seen among living marine organisms (e.g., Shen et al., Reference Shen, Jamandre, Hsu, Tzeng and Durand2011; Dolby et al., Reference Dolby, Ellingson, Findlay and Jacobs2018). Paleontological evidence suggests that nearshore marine sites were commonly those in which novel morphologies or ecologies first appeared (e.g., Jablonski et al., Reference Jablonski, Sepkoski, Bottjer and Sheehan1983; Jablonski and Bottjer, Reference Jablonski, Bottjer and Nikecki1990; Sepkoski, Reference Sepkoski1991; Jacobs and Lindberg, Reference Jacobs and Lindberg1998). Given this, detailed studies of morphological and ecological evolution in ancient nearshore sites have particular currency but are challenging because such environments are generally poorly preserved. The late Cambrian of the Upper Mississippi Valley provides an important and potentially instructive exception.
Geological setting
Localities
Specimens of Walcottaspis vanhornei are known from at least three localities (Fig. 1) from which most were obtained over one hundred years ago. One of these sites, Hokah (HH), was recollected by the late Gerald O. Gunderson and N.C.H in recent years. It is located at the intersection of Carlson Road and Hwy 16, ~ 4 km west of Hokah, Houston County, Minnesota (43°46′0.12″N, 91°25′26.4″W) and is apparently the site previously recorded as “Whiteman's Quarry on Mt Hope,” USGS Locality #346d (Hughes, Reference Hughes1993, p. 25). Previous records also mention another site at Hokah yielding W. vanhornei to be “near old railroad embankment” or “Rail Road Dam,” but this is unlikely to have been the HH site because that is at the top of a notable bluff, as are all outcrops of the St. Lawrence Formation in this area.

Figure 1. Location of Upper Mississippi Valley (UMV) region of the North American cratonic interior showing where lower Paleozoic strata are present. Localities yielding Walcottaspis vanhornei are shown with diamond-shaped symbols (i.e., Bluff Siding, Wisconsin; Winona and Hokah, Minnesota). Reno, Minnesota, is shown with a brown circle. Inset shows the location of UMV region (shaded area) with respect to the late Cambrian continental lithofacies belts of Palmer (Reference Palmer, Boettcher and Sloan1960). Depositional strike lines are based on Runkel et al. (Reference Runkel, Miller, McKay, Palmer and Taylor2007), and Eoconodontus notchpeakensis–Saukiella (En/Sauk) subzone boundary is based on Bell et al. (Reference Bell, Berg, Nelson and Rodgers1956), Nelson (Reference Nelson1956), and Miller et al. (Reference Miller, Evans, Loch, Ethington, Stitt, Holmer and Popov2003).
The specific positions of two other sites that have yielded reposited W. vanhornei specimens are unknown, but given the density of settlement in the area at the time they were collected, both localities will have been at most within a few kilometers of the places named. The Winona (WI; see Hughes, Reference Hughes1993), Winona County, Minnesota, site was reported as on the bluffs facing the Mississippi River, immediately west of the town and in the general vicinity of 44°3'N, 91°43'W (Walcott, Reference Walcott1914, p. 373). Bluff Sliding is in Buffalo County, Wisconsin, and lies on the eastern bank of the Mississippi River immediately opposite Winona. The St. Lawrence Formation crops out in the bluffs above the settlement, for example, in an overgrown roadcut at 44°5′8.82″N, 91°37′15.12″W, but the specific locality from which the specimens were collected is also unknown. The only other reported occurrences of W. vanhornei known to us are that of Raasch (Reference Raasch1951, p. 142), who reported that Charles Bell and Robert Berg made a find near Reno, Minnesota, and Ulrich and Resser's (Reference Ulrich and Resser1930, p. 66) report of “an imperfect cranidium” from Lodi, Wisconsin. No specimens from near Reno are available, but the settlement at Reno accords with an arc of outcrop defined by the other three localities and the Runkel et al. (Reference Runkel, Miller, McKay, Palmer and Taylor2007) prograding shoreface model (Fig. 1), and we consider this report to be credible. Although to our knowledge we have not seen the Lodi specimen to which Ulrich and Resser (Reference Ulrich and Resser1930) referred, occurrence of genuine W. vanhornei at Lodi is unlikely given its geographical position (and stratigraphic position as inferred from Runkel et al., Reference Runkel, Miller, McKay, Palmer and Taylor2007) and, as Walcottaspis cranidia strongly resemble those of D. minnesotensis Owen, Reference Owen1852, we consider this reported occurrence to be unsubstantiated.
Taphonomy
Sclerites of W. vanhornei became disarticulated, apparently completely, before recovery and are somewhat flattened/fractured composite molds. They occur in dolomitic siltstone layers within the heterolithic facies of the St. Lawrence Formation (Fig. 2) and are preserved in a manner similar to many examples of their close relative D. minnesotensis (see the following and Hughes, Reference Hughes1993). The W. vanhornei we have collected lie parallel to bedding (Figs. 3–6), and this appears to be their general condition (in contrast to some beds rich in Dikelocephalus, in which specimens may be inclined). The preservation of conjoined free cheeks along with apparently intact genal spines (Figs. 4.10, 4.11, 6.1) argues against significant reworking at the sediment–water interface. Early ontogenetic stages are missing, perhaps because their sclerites were insufficiently robust to be preserved in this environment or juvenile growth was spatially localized elsewhere (e.g., Beheregaray and Sunnucks, Reference Beheregaray and Sunnucks2001). Specimens may have derived from assemblages of individuals alive penecontemporaneously, or from post-exuvial trilobite assemblages, followed by some degree of amalgamation among temporally successive assemblages (Olszewski, Reference Olszewski1999). Thus, the “paleontological resolution” (Kowalewski and Bambach, Reference Kowalewski, Bambach and Harries2003), the span of time represented between those individuals alive most recently and those alive longest ago, is likely to have been no longer than years to decades at most. It may be that some beds exclusively represent exuviae and/or carcasses of contemporary individuals that have since been only slightly reworked taphonomically. Thus, these assemblages may offer a glimpse of the ambient local seafloor situation at high temporal resolution, as is increasingly being recognized among such deposits (e.g., Paola et al., Reference Paola, Ganti, Mohrig, Runkel and Straub2018).

Figure 2. Stratigraphic section of St. Lawrence Formation and adjacent units at locality Hokah (HH), Houston County, Minnesota. Left column depicts lithology and degree of induration with glauconite (G) and mudstone (black). Symbols on the right side of the log refer to the presence of dendroid graptolites (tuning-fork-like symbol), inarticulate brachiopods (‘∇’), and aglaspidids (half-moon-like symbol). The trilobite occurrences marked on the log include: Illaenurus quadratus (Iq), Dikelocephalus minnesotensis (Dm), Walcottaspis vanhornei (W), Tellerina (T). Mbr = member; Fmn = formation; f/sd facies = feldspathic facies. See Hughes and Hesselbo (Reference Hughes and Hesselbo1997) for further details.

Figure 3. (1–12) Walcottaspis vanhornei (Walcott, Reference Walcott1914) cranidia from the heterolithic facies of the St. Lawrence Formation at Hokah, Minnesota, in the Upper Mississippi Valley: (1) FMNH-UC23314c; (2) FMNH-PE39215; (3) FMNH-PE39214; (4) CMC-IP-89411i (formerly UMPC2443i); (5) CMC-IP-87565j (formerly UMPC2443j); (6) FMNH-UC23314a; (7) CMC-IP-89411h (formerly UMPC2443h); (8) CMC-IP-87565x (formerly UMPC2443x); (9) CMC-IP-95630 (formerly UMPC3689); (10) UWGM 7151 (formerly UW4006-491); (11) FMNH-UC14393a, paralectotype; (12) CMC-IP-87565t (formerly UMPC2443t). (13) Small holaspid cranidium, likely Dikelocephalus, from Trempealeau, Wisconsin, holotype of D. juvenalis Ulrich and Resser, Reference Ulrich and Resser1930, USNM-PAL-58601. (1–12) Scale bars = 20 mm; (13) scale bar = 10 mm.

Figure 4. (1–9) Walcottaspis vanhornei (Walcott, Reference Walcott1914) sclerites from the heterolithic facies of the St. Lawrence Formation at Hokah, Minnesota in the Upper Mississippi Valley: (1) detail of glabella of FMNH-UC14393a, paralectotype; (2) detail of posterolateral border, FMNH-UC14393a, paralectotype; (3) hypostome, FMNH-UC23314f; (4) counterpart of right librigena, FMNH-PE82161; (5) pixel inversion of (4); (6) two right librigena on same slab, FMNH-UC23314h; (7) detail of FMNH-UC14393b showing terrace ridges, lectotype; (8) axial part of thoracic segment, FMNH-PE82162. (9) Pixel inversion of counterpart thoracic segment, FMNH-UC14393c, paralectotype. (10) Conjoined free cheek from Bluff Siding, Wisconsin, MCZ-IP917 (arrow points to an example of a sinuous structure interpreted to reflect infestation); (11) counterpart of conjoined free cheek from Bluff Siding, Wisconsin, MCZ-IP917. Scale bars = 20 mm.
Westrop (Reference Westrop1986, p. 29) considered the effect of compaction on the cranidial morphology of Walcottaspis, then known only from one illustrated specimen, and suggested that Lochman's (in Harrington et al., Reference Harrington, Henningsmoen, Howell, Jaanuson, Lochman-Balk and Moore1959, p. 254–255) diagnosis of its low relief may reflect sediment compaction. Specimens of both D. minnesotensis and W. vanhornei from the heterolithic facies show similar patterns of flattening and compression-related cracking, with larger specimens such as FMNH-UC14393a (Fig. 3.11) showing an overall greater degree of cracking than smaller ones, as is typical among trilobites (Hughes, Reference Hughes and Harper1999). However, differences in the cranidial morphologies of these species cannot be assigned primarily to taphonomic effects. While all W. vanhornei cranidia show effaced S1 and S2 furrows, several such specimens preserve steep-sided, markedly inflated glabellae that have not suffered significant compaction (Fig. 3.1, 3.3, 3.6, 3.9). Accordingly, effacement of the glabellar furrows was an original biological feature and serves to distinguish Walcottaspis cranidia from those of D. minnesotensis, in which these furrows are notably incised.
As is also common in D. minnesotensis (see Hughes, Reference Hughes1993, p. 15), infestation resulted in shallow, flat-bottomed grooves etched into the interior surfaces of both the dorsal surface and the doublure of W. vanhornei. As the specimens are preserved as composite molds, these original grooves can also appear as ridges when cast on the underside of the dorsal exoskeleton. The organism responsible was presumably exploiting the interface between the internal surface of the exoskeleton and the soft tissues immediately adjacent. Grooves on the interiors of the dorsal and ventral surfaces are between 0.48 and 0.80 mm wide and do not mirror each other, suggesting independent origin on each surface. As in D. minnesotensis, these structures are particularly evident in the pygidium (Fig. 5.2, 5.3, 5.4, 5.7), where they, although sinuous or with angular junctions, tend to extend between the axis (where their author presumably gained entrance) and the lateral margin. A possibly similar groove occurs in a free cheek (Fig. 4.10). These structures were likely the result of either a metazoan parasite or a scavenger that was mining associated soft tissues and that partially eroded the exoskeleton as it did so. The similar morphologies and width ranges of these trails in both W. vanhornei and D. minnesotensis suggest the action of a similar author in each and may be evidence of a shared infester.

Figure 5. (1–3, 5, 7, 8) Walcottaspis vanhornei (Walcott, Reference Walcott1914) pygidia from the heterolithic facies of the St. Lawrence Formation at Hokah, Minnesota, in the Upper Mississippi Valley: (1) FMNH-PE39210; (2) CMC–IP 97568 (formerly UMPC6352); (3) CMC-IP 89411c (formerly UMPC2443c). (4) From Winona, Minnesota, USNM-PAL 72687A. (5) UWGM 7161 (formerly UW4006-501). (6) From Bluff Siding, Wisconsin, MCZ-IP910. (7) FMNH-UC14393b, lectotype; (8) CMC-IP 89411a (formerly UMPC2443a). Scale bars = 20 mm. Arrows in (4, 7) point to examples of sinuous structures interpreted to reflect infestation.

Figure 6. Reconstruction of holaspid Walcottaspis vanhornei. (1) Dorsal view of cranidium with median tubercle, free cheek with eye platform, and surface ornamentation; ventral view of conjoined free cheek with terrace ridges and hypostome with surface ornamentation; and dorsal view of pygidium showing petaloid facet, paradoublural line (dashed), and terrace ridges on the dorsal surface of the venter ventral side of pygidium. (2) Dorsal view of holaspid with estimated trunk proportions. The relative sizes of the cephalon and pygidium are drawn such that, upon putative enrollment, their marginal outlines match optimally in shape (see text for details).
Sedimentology
The St. Lawrence Formation is a fine-grained mixed carbonate mud–sandstone unit comprising three lithofacies that include rare stromatolitic dolomite, a laminated feldspathic sandstone facies containing notable flat pebble conglomerates, and a heterolithic facies in which very fine-grained feldspathic sandstone layers are interbedded with dolomitic mudstones (Hughes and Hesselbo, Reference Hughes and Hesselbo1997). All W. vanhornei specimens yet recovered are preserved in dolomitic mudstone layers of the heterolithic facies. At Hokah, specimens were collected toward the top of the St. Lawrence Formation from a 0.95 m thick buff-colored dolomitic siltstone bedset located about 6.4 m above the base of the section (Fig. 2). Walcottaspis vanhornei specimens are concentrated in a level about 15 cm above the base of this layer. A similar ~0.5 m dolomitic siltstone containing D. minnesotensis, but apparently no W. vanhornei, occurs about one meter above this level, with its base at about 8.2 m in the log (Figs. 2, 7).
Co-occurrent fossils and biostratigraphy
Both the Hokah bedsets containing the large species D. minnesotensis and W. vanhornei also yielded disarticulated sclerites of the trilobite Illaenurus quadratus Hall, Reference Hall1863 and linguiliform brachiopods. The aglaspidid Cyclopites vulgaris Raasch, Reference Raasch1939 co-occurs with W. vanhornei, and specimens belonging to the trilobite genus Tellerina occur along with D. minnesotensis (Fig. 7) in the upper layer, along with fragmentary aglaspidid remains (Fig. 2).

Figure 7. Dikelocephalus minnesotensis pygidium (UWGM7065) recovered from a dolomitic siltstone bed, the base of which is at about 8.2 m in the Hokah (HH) outcrop section, approximately one meter above the level containing the Walcottaspis vanhornei specimens collected in this study. This helps demonstrate that W. vanhornei occurs within the temporal range of D. minnesotensis. Scale bar = 20 mm.
Raasch (Reference Raasch1951, p. 148) suggested that W. vanhornei defines a distinct biostratigraphic level, the eleventh within his Saukia Zone. This interpretation places W. vanhornei as a stratigraphically late form in the area and is supported by recent sequence stratigraphic analysis. More recent work suggests that W. vanhornei was represented within a local, informally defined Saukiella trilobite subzone (Runkel et al., Reference Runkel, Miller, McKay, Palmer and Taylor2007, figs. 5, 6) that lies within the Eoconodontus conodont Zone and contains among the youngest St. Lawrence Formation rocks outcropping in the area. The appearance of Eoconodontus notchpeakensis Miller, Reference Miller1969 defines the first subzone of the Eoconodontus Zone. This species has been found to the east of Hokah but also in sites farther to the west but has not been recovered as yet from the Hokah section. The W. vanhornei-bearing bed at Hokah almost certainly lies within the Eoconodontus Zone, but it is not yet clear whether it is within the Eoconodontus notchpeakensis subzone or the overlying Cambrooistodus minutus subzone, the eponymous species of the latter being a form largely restricted to strongly carbonate-rich facies (A.C. Runkel, personal communication, 2022). Walcottaspis vanhornei thus occurs within the younger strata of the St. Lawrence Formation, but not the youngest yet known, which occur farther to the southwest.
Sequence stratigraphic setting
The heterolithic facies of the St. Lawrence Formation represents the toesets of progradational parasequences deposited early during the deposition of a falling-stage systems tract (Runkel et al., Reference Runkel, Miller, McKay, Palmer and Taylor2007) at ca. 490 Ma. Although recognizing individual parasequences within the St. Lawrence Formation can be challenging (perhaps because these deposits represent the toesets of the prograding shoreline where depositional and preservational events were relatively rare and haphazard), the beds containing W. vanhornei and D. minnesotensis at Hokah evidently belong to different parasequences and are separated by seven ~10 cm thick beds of parallel laminated feldspathic sandstone with burrowed tops, which may each represent deposition from a notable storm (Fig. 2).
During deposition of the highstand and falling-stage systems tracts, the shoreline in the Upper Mississippi Valley prograded basinward (i.e., toward the southwest) (Runkel et al., Reference Runkel, Miller, McKay, Palmer and Taylor2007). As the rate of relative sea-level rise declined, sediment supply from the land exceeded the generation of accommodation space, leading to deposition of regressive facies and the outward migration of the shoreface. The result was a series of parasequences whose lateral facies coarsened shoreward, the more offshore, distal toesets of which were characterized by carbonate muds of the heterolithic facies. Their proximal equivalent was fine-grained sandstones characteristic of the Norwalk Member of the Jordan Formation, but similar lithologies frequently also interbed with the finer-grained carbonates within the heterolithic facies of the St. Lawrence Formation, as stated. Such sandstone beds resulted from depositional events of sufficient magnitude to transport land-derived sediment into this part of the basin and, where concentrated, constitute the feldspathic sandstone facies of the St. Lawrence Formation. Flat pebble conglomerates are also common in these facies and, in some cases, indicate episodes of marked stratigraphic condensation (Hughes and Hesselbo, Reference Hughes and Hesselbo1997).
The geographical positions of the three verified localities from which known W. vanhornei specimens have been recovered fall almost exactly along the projected position of the prograding shoreline according to Runkel et al. (Reference Runkel, Miller, McKay, Palmer and Taylor2007, fig. 1; Fig. 8). In doing so, they provide additional evidence for the validity of this model. This distribution strongly suggests that the temporal range of W. vanhornei was notably short, as Raasch's (Reference Raasch1951) biostratigraphic scheme anticipated. To date, W. vanhornei has been localized to only a single parasequence at Hokah, and its distribution among localities does not exclude the possibility of occurrence within a single parasequence throughout the region (Fig. 8). This is a minimum, but given the narrow outcrop belt of its occurrence, W. vanhornei is unlikely to have been represented at the abundance seen in the bed at Hokah in more than a small number of parasequences, with five being a maximum estimate. On the basis of an estimate of the rate of deposition of the entire Croixan Series, Runkel et al. (Reference Runkel, Miller, McKay, Palmer and Taylor2007, fig. 11) suggested the average duration of a parasequence within the basin to range between 75,000 and 150,000 years. However, during the early falling-stage systems tract, parasequence duration was likely to have been at the short end of this range and perhaps closer to 50,000 years (A.C. Runkel, personal communication, 2021). The duration of the known fossil record of W. vanhornei thus likely represents an interval not much greater than about 250,000 years, and likely substantially shorter.

Figure 8. Spatial distribution of Walcottaspis vanhornei in the four localities from Figure 1, consistent with occurrence possibly restricted to the geographical range of a single parasequence of the En/Sauk subzone as defined by Runkel et al. (Reference Runkel, Miller, McKay, Palmer and Taylor2007).

Figure 9. Reconstruction of Walcottaspis vanhornei cranidium showing the set of 22 landmarks selected to optimally capture variation among the specimens available. Specimens had to preserve all four axial landmarks and at last one of each of the paired abaxial landmarks to be included in the dataset. Type 1 landmarks (Bookstein, Reference Bookstein1991) are shown as gray-shaded circles, Type 2 landmarks as open circles.
Although we have yet to recover trilobites within the St. Lawrence Formation at Hokah below the layer containing W. vanhornei (Fig. 2), regional relationships show that D. minnesotensis occurred widely throughout the region in both the heterolithic and feldspathic sandstone facies, including in areas where St. Lawrence Formation deposition predated that recorded at Hokah. The occurrence of D. minnesotensis both in the next fossil-bearing bed above those containing W. vanhornei (Figs. 2, 7) and at localities farther west, such as Rushford, Minnesota, confirms Raasch's (Reference Raasch1951) view that the range of W. vanhornei occupied only a small portion of the regional temporal range of D. minnesotensis and that D. minnesotensis evidently regained its local abundance in the heterolithic facies shortly after a time in which W. vanhornei apparently had sole local representation.
Published views on Walcottaspis vanhornei morphology and systematic affinities
Dikelocephalus vanhornei was introduced by Walcott (Reference Walcott1914, p. 373), who illustrated and described one counterpart cranidium, one thoracic segment, and one pygidium. He noted that although its cranidium was closely similar to that of the comparably large trilobite D. minnesotensis, its pygidium was strikingly different. The genus Walcottaspis was established by Ulrich and Resser (Reference Ulrich and Resser1930, p. 66–68), who refigured Walcott's part cranidium and pygidium, along with the illustration of one additional pygidium. Their justification for generic status was the markedly distinct pygidial structure including the termination of the pleural furrow abaxially to the axial furrow, the one fewer clearly defined axial segment, the absence of marginal spines, the transverse anterior pygidial margin, and its elliptical posterior margin. With respect to closest relatives, Walcott (Reference Walcott1914) compared W. vanhornei only with D. minnesotensis, but Ulrich and Resser (Reference Ulrich and Resser1930, p. 64) placed it with species of the genus Osceolia, also known only from the Upper Mississippi Valley, in their newly defined subfamily Osceolinae, from which Dikelocephalus was excluded. The basis for this grouping was that the pygidial interpleural furrows of Walcottaspis and Osceolia are partly or wholly obsolete and that the members of both genera have only four clearly defined pygidial axial rings, rather than the five that they considered characteristic of Dikelocephalus (Ulrich and Resser, Reference Ulrich and Resser1930, p. 65–66). They further supported this assertion by reference to unpublished specimens suggested to be allied to Osceolia from the “Devil's Lake Sandstone,” a nearshore facies variant of the Reno Member of the Tunnel City Group but which lack marginal pygidial spines. The name that Ulrich and Resser (Reference Ulrich and Resser1930, p. 68) gave to that material, “Osceolina diabolo,” is a nomen nudum, the whereabouts of its specimens is unclear, and no further material of this kind has subsequently been reported.
Lochman (Reference Lochman1956, fig. 2) suggested that Walcottaspis was derived from an unspecified species or species lineage within the genus Briscoia, which she considered to be explicitly polyphyletic and derived directly, in her view, from two different species of the idahoiid Wilbernia (Fig. 10). She did not provide character-based support for the particular links invoked but wrote of certain “irreversible trends in the evolution of Dikelocephalidae” (Lochman, Reference Lochman1956, p. 449), some of which, such as the squaring of the anterior glabella and the obsolescence of the anterior border furrow, are present in W. vanhornei. Others, such as increased eye length and shortened posterolateral border relative to putative ancestors, are not. Given Lochman's (Reference Lochman1956) recognition of explicitly polyphyletic genera without specification of putative phylogenetically informative characters within these, the reasoning for her association of W. vanhornei with a species within Briscoia is unclear, as is that justifying her derivation of Osceolia and two branches of a polyphyletic Dikelocephalus from other unspecified Briscoia species.

Figure 10. Wilbernia sp. from the “Hudson Member” of the Lone Rock Formation in the Upper Mississippi Valley from the Winona (Minnesota)/Fountain City (Wisconsin) region showing notable pygidial homeomorphy with W. vanhornei. (1) UWGM7069, cranidium. (2) UWGM7070, latex of counterpart pygidium. Scale bars = 20 mm.
Westrop's (Reference Westrop1986, p. 29) consideration of the effect of compaction (see the preceding) compared W. vanhornei directly with St. Lawrence Formation Dikelocephalus, and no other taxa.
A report of two cranidia assigned to Walcottaspis sp. from substantially older Cambrian rocks in the Himalaya (Shah et al., Reference Shah, Parcha and Raina1991, p. 91, pl. 1j, n) was based on misidentification. The Himalayan specimens, although effaced, differ in having a trapezoidal glabella, strong eye ridges, and in one case, an anterior border. These authors reported the specimens to be of Guzhangian age, some 7.5–6.0 Myr older than W. vanhornei, although in our opinion these forms are likely still older. Their features compare better to Wuliuan forms such as Paramecephalus (see Zhang et al., Reference Zhang, Lu, Chu, Qian, Lin, Zhou, Zhang and Yuan1980, pl. 117, 118, figs. 1–6; Jell and Hughes, Reference Jell and Hughes1997, pl. 12; Luo et al., Reference Luo, Hu, Hou, Gao, Zhan and Li2009, pl. 13, figs. 3, 4) or Xingrenaspis dardapurensis (Jell and Hughes, Reference Jell and Hughes1997, pl. 18, figs. 4–11).
Hughes (Reference Hughes1994, p. 53) noted that the pygidial axis of W. vanhornei displays fewer rings than that of D. minnesotensis and suggested that the difference in their morphology might be partly explained if holaspid D. minnesotensis retained in its pygidium a segment homologous to the one released into the thorax in holaspid W. vanhornei.
Phylogenetic position
Herein we have used primarily parsimony-based phylogenetic analysis, supplemented by a Bayesian maximum likelihood approach, to assess phylogenetic relationships between the various taxa of dikelocephalids bearing an extended cephalic doublure. The cladistic analysis of Dikelocephalidae conducted here uses a branch and bound search of 38 characters divided into a total of 98 character states across 16 ingroup taxa and one outgroup (Table 1; Supplementary Data 1). As some characters varied along a continuous scale, we also conducted a separate parsimony-based analysis treating them as such, rather than with user-imposed, discretized character state divisions as used in the first analysis. Eleven out of a total of 38 characters were continuous and were measured using the most complete sclerites of each species considered (see table in the Supplementary Data 1 for the character states of continuous characters). For both the discretized dataset and the mixed continuous character and discrete character dataset, the outgroup was Prosaukia misa (Hall, Reference Hall1863), chosen for its close similarity with dikelocephalids in having a quadrate glabella and elliptical pygidium but not showing the extended doublure. The species used in the phylogenetic analysis are listed in Supplementary Data 1, with references to their published figures and unpublished images from our image library.
Table 1. Matrix of character states for selected taxa. See Supplementary Data 1 for a descriptive list of characters, character states, and taxa used in this phylogenetic analysis. Columns correspond to character state designations, and rows contain taxa names.

The parsimony analyses were run using Tree Analysis Using New Technology (TNT; Goloboff et al., Reference Goloboff, Farris and Nixon2008), and Winclada (Nixon, Reference Nixon2002) was used to map transitions in discrete character states (Fig. 11.1, 11.2) and to calculate support indices (Supplementary Data 2, figs. 1, 2). For the discretized character-state analysis, an implicit enumeration search in TNT resulted in four most parsimonious trees (of which the strict consensus tree is depicted in Fig. 11.1) and a tree length of 141, consistency index of 39, and retention index of 33. Although overall tree resolution is poor, relationships between most well-represented dikelocephalids from the Upper Mississippi Valley are supported by unambiguous synapomorphies and suggest common ancestry among Dikelocephalus species and W. vanhornei, with W. vanhornei as sister taxon to D. minnesotensis. Bootstrap resampling was calculated by the GC method for 1,000 replicates with the implicit enumeration search option (Supplementary Data 2, fig. 1). Bremer support (Bremer, Reference Bremer1988, Reference Bremer1994) was calculated by default TBR branch swapping, setting the suboptimal limit to six steps. Both these metrics suggest that relationships of W. vanhornei reported in the preceding receive the strongest support within the analysis. Maximum likelihood analysis of the same dataset using MrBayes yields results consistent with this parsimony-based analysis (Supplementary Data 6), specifically that D. freeburgensis Feniak in Bell et al., Reference Bell, Feniak and Kurtz1952 is the sister taxon to a clade bearing W. vanhornei and D. minnesotensis, and that these relationships have posterior probability support comparable to that of other morphological character-based analyses.

Figure 11. Parsimony-based phylogeny of dikelocephalids using Prosaukia misa as the outgroup. (1) Strict consensus tree of the four most parsimonious tree found for this character matrix using all characters divided into discretized character states. (2) Single tree found for the character matrix using a mixture of discretized and continuous characters. Numbers on top of node branches show the character number, and numbers on bottom show the state for that character. Filled circles indicate unambiguous derived character states while open circles indicate convergent derived character states. Refer to Table 1 for codings, Supplementary Data 1 for list of characters, character states, and selected taxa; refer to Supplementary Data 2 for Bremer support and bootstrap resampling results.
For the combined discrete and continuous character-based analysis, TNT implicit enumeration search resulted in a single tree. The 27 discrete characters from this dataset were mapped onto the most parsimonious tree found by TNT (Fig. 11.2), with tree length of 72, consistency index of 48, and retention index of 50. Overall tree resolution is improved and maintains shared ancestry among Dikelocephalus species and W. vanhornei, with W. vanhornei as sister taxon to D. minnesotensis. Again, Bremer support and bootstrap resampling show these relationships to receive relatively high levels of support (Supplementary Data 2, fig. 2). This analysis confirms that these phylogenetic conclusions are not the result of arbitrary character state divisions imposed during the discretization of continuous variables.
The results of both the parsimony-based phylogenetic analysis and of maximum likelihood analyses conducted to date suggest W. vanhornei to be sister taxon to Dikelocephalus minnesotensis, whose stratigraphic range encompasses that of W. vanhornei (Fig. 11) and support Walcott's (Reference Walcott1914) original comparison between these taxa. The older Dikelocephalus form from the transgressive systems tract in the Reno Member, Tunnel City Group, Dikelocephalus freeburgensis, is sister taxon to the clade comprising D. minnesotensis and Walcottaspis vanhornei. Osceolia species lie outside the clade bearing Dikelocephalus and Walcottaspis, and this result predicts that Osceolia or its closest relatives will occur in the stratigraphic record before the appearance of the Dikelocephalus–Walcottaspis clade. Neither Ulrich and Resser's (Reference Ulrich and Resser1930) nor Lochman's (Reference Lochman1956) proposed relationships of W. vanhornei are supported, and Briscoia remains either an association of plesiomorphic species with unresolved relationships (Fig. 11.1) or polyphyletic (Fig. 11.2). Most Briscoia species are based on small numbers of specimens and occurrences widely scattered across Laurentia. A more extensive phylogenetic analysis of dikelocephalids and their relatives is forthcoming but beyond the scope of this paper.
Ulrich and Resser (Reference Ulrich and Resser1930) justified Walcottaspis among dikelocephalids largely on the basis of its phenetically distinctive pygidium (see the preceding), and this opinion has not been challenged by any subsequent worker to date. One implication of our phylogenetic analyses is that, if the genus Walcottaspis is maintained, the genus Dikelocephalus is paraphyletic (Fig. 11). We note this implied paraphyly but defer further consideration until we complete a comprehensive investigation of dikelocephalid morphometry and systematics.
Shape variation
Although specimens assigned to Walcottaspis vanhornei share definitive features, morphological variation is evident among its members. Here we use semilandmark- and landmark-based geometric morphometric analysis to explore such variation. Specimens were imaged in dorsal view with the palpebral lobe surface mounted horizontally and the axial pygidial furrow mounted horizontally. A set of landmarks were selected: 22 in the cranidium (Fig. 9) and 40 landmarks and 216 semilandmarks in the pygidium, with semilandmarks used to capture information about the pleural and interpleural furrows (Fig. 12.1, 12.2). A pygidial analysis with only the 14 landmarks present on the axial lobe and the axial posterior margin was also conducted (Fig. 12.3). Except where prohibited, specimens were coated with ammonium chloride sublimate before being photographed. The image library was assembled intermittently between 1986 and 2021 and mostly using a Nikon SRL camera with images captured either as 35 mm black-and-white negatives or directly as digital images. Most were taken within the specimen's host institution. Negatives were digitally scanned with a Polaroid SprintScan 35 plus or an Epson Perfection V700 scanner. Cartesian X and Y coordinates of morphological landmarks for each specimen of cranidia and pygidia were recorded using the freely available software tpsDig (http://www.sbmorphometrics.org/soft-dataacq.html) and ImageJ (https://imagej.nih.gov/ij/download.html). Morphometric analysis of the landmark dataset was conducted using RStudio interface in R (https://www.rstudio.com). In R, the packages used for statistical analysis were geomorph (Adams et al., Reference Adams, Collyer, Kaliontzopoulou and Baken2021; Baken et al., Reference Baken, Collyer, Kaliontzopoulou and Adams2021) for most of the morphometric functions and prWarp (Mitteroecker et al., Reference Mitteroecker, Bartsch, Erkinger, Grunstra, Le Maître and Bookstein2020; Grunstra et al., Reference Grunstra, Bartsch, Le Maître and Mitteroecker2021) for partial warp analysis. For fitting linear regression models, Residual Randomization Permutation Package (Collyer and Adams, Reference Collyer and Adams2018, Reference Collyer and Adams2021) was used as it has a high statistical power to identify patterns in data (Anderson and Braak, Reference Anderson and Braak2003). Preservation in dolomitic siltstone resulted in specimen compaction, which inflates variance due to taphonomic, as opposed to biological, reasons. While we are confident that such variance does not overwhelm the original biological signal, it conflates it in such a way that we cannot confidently interpret the biological significance of very small differences among specimen shape. Accordingly, rather than some of the more sophisticated estimates of measurement error used in recent analyses (see Hughes et al., Reference Hughes, Adrain, Holmes, Hong and Hopkins2021), we used a simple estimate of measurement error involving multiple remounting, reimaging, and redigitizing of three cranidia (Supplementary Data 5). The result of this analysis is that measurement error accounted for 4.84% of the total cranidial variance and thus was not a major contributor to the within-group or intergroup variance observed (see Supplementary Data 5). Bookstein (Reference Bookstein1991, p. 63–66) defined “type 1” landmarks when three surfaces intersect (“discrete juxtapositions of tissues”) and “type 2” landmarks when two surfaces intersect (“maxima of curvature”), noting that arguments for positional homology are stronger for type 1 landmarks than for type 2. This study utilizes both these landmark types (Figs. 9, 12), but because W. vanhornei is somewhat effaced, the majority are type 2. Although we preferred to use type 1 landmarks where possible, measurement error in shape is broadly comparable to those of trilobites with a higher proportion of type 1 landmarks (e.g., Hong et al., Reference Hong, Hughes and Sheets2014), a finding echoed by a recent comparative analysis among landmark categories (Wärmländer et al., Reference Wärmländer, Garvin, Guyomarc'h, Petaros and Sholts2019). The high proportion of type 2 landmarks used herein does not appear to have notably inflated observed variance.

Figure 12. Reconstruction of Walcottaspis vanhornei pygidium showing the landmarks used to assess morphological variation in a set of 10 specimens. (1) Landmark scheme comprising 40 landmarks capturing features of the axis, the pygidial margin, and extent of pleural and interpleural furrows. (2) Landmark scheme of 256 landmarks, including the 40 landmarks mentioned in (1) combined with 216 semilandmarks used to trace the positions of the pleural and interpleural furrows. (3) Landmark scheme of 14 landmarks capturing only the axial features of the pygidium. Type 1 landmarks are shown as gray-shaded circles, Type 2 landmarks as open circles. Eight additional Type 1 landmarks mark confluence of the pleural and interpleural furrows with the axial furrow. They are not shown in (1) and (2) because they have near overlap with those landmarks at the intersection of the axial and inter-ring furrows.
Results
Cranidial shape changes
The occipital-glabellar length of available W. vanhornei cranidia ranges from 1.40 mm to 4.30 mm, and across this span, analysis of cranidial shape change was based on 22 cranidial landmarks superimposed using general Procrustes superimposition of shape configurations of 17 cranidial specimens (Fig. 13).

Figure 13. Procrustes superimposition of 22 landmarks for holaspid cranidia of W. vanhornei, (N = 17).
A principal component analysis (PCA) of the partial warp scores (derived from the thin-plate spline analysis of Procrustes aligned landmarks based on the mean shape of the whole sample) shows that 68.44% of the shape variance is captured by the first three relative warps: Relative Warp 1 (RW 1), RW 2, and RW 3, respectively, explain 38.34%, 18.73%, and 11.37% of the total shape variance (Fig. 14.1). Each of the other 13 RWs accounts for less than 10.00% of the total variance, and they are not considered further (Supplementary Data 3). The deformation grids for the principal components were obtained by comparing minimum with maximum scores on each RW. RW 1 depicts an increase in relative length of the frontal area (sag.) and an overall contraction of the proportions of the glabella and of the eye (tr.) compared with other structures (Fig. 15.1). RW 2 depicts change in curvature of the frontal area, extension (sag.) of the L2 glabellar lobe, and contraction (sag.) of the L1 glabellar lobe (Fig. 15.2). RW 3 reflects widening of the fixigena (tr.) and of the frontal area (tr.) (Fig. 15.3). RW 1 and RW 2 display an interesting relationship in that specimens that score high on one axis score low on the other, and also show possible grouping (Fig. 14.1).

Figure 14. Bivariate scatter plots of first two principal components showing Walcottaspis vanhornei cranidia and pygidia categorized according to occurrence in the three localities. (1) Plot of principal components 1 and 2 for cranidial dataset showing 17 specimens from Hokah, Minnesota (blue inverted triangle). (2) Plot of principal components 1 and 2 for pygidial dataset showing eight specimens from Hokah, Minnesota (blue inverted triangle), one specimen from Winona, Minnesota (black circle), and one specimen from Bluff Siding, Wisconsin (yellow square).
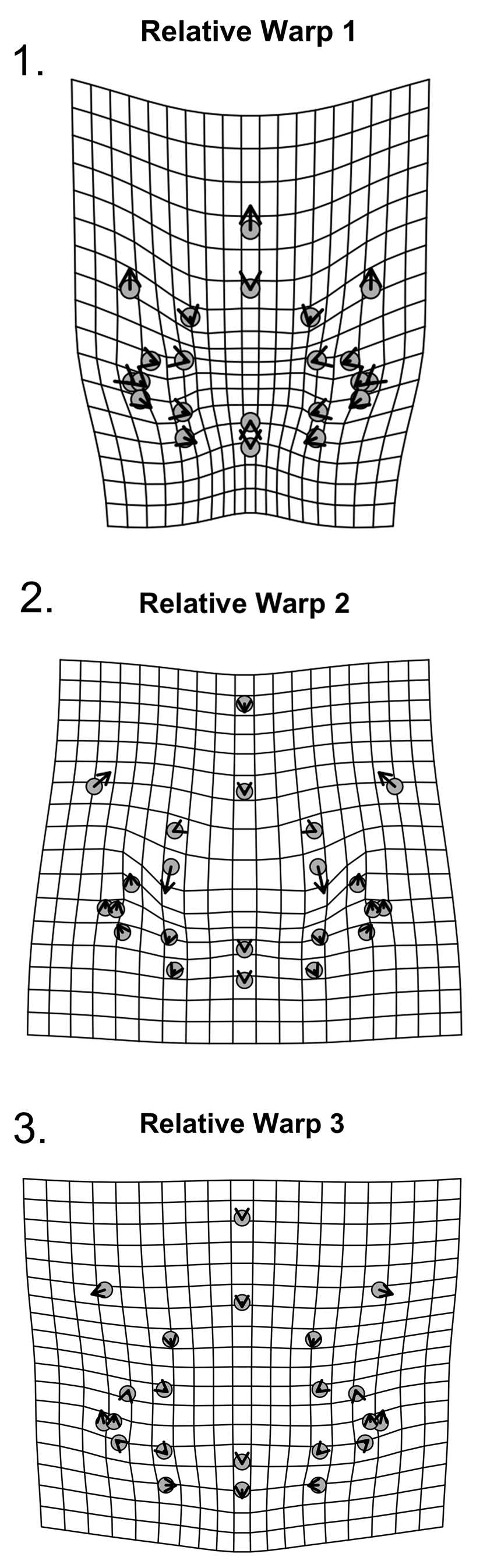
Figure 15. Thin-plate spline deformation grid of relative warps for 22 landmarks for holaspid cranidia of W. vanhornei (N = 17). (1) Shape variation related to RW1. (2) Shape variation related to RW2. (3) Shape variation related to RW3.
To investigate whether any of these patterns reflect ontogenetic change, the partial Procrustes distance of individual specimens from the mean of the smallest two cranidia specimens in the dataset was calculated and plotted against the natural log of centroid size as estimated by generalized Procrustes analysis (Fig. 16). A significant positive relationship exists between partial Procrustes distance and ln CCS (the natural log of centroid size, slope = 0.0486, p = 0.002267, r = 0.7241). Regression of partial warp scores from the mean shape of the smallest two cranidia specimens against ln CCS is significant (p = 0.003996 for 1,000 bootstraps) and suggests that 16.39% of overall shape variance captures ontogenetic variation. A deformation plot was computed by regression of Procrustes aligned landmark data onto the ln CCS and using the predicted shapes from the regression model. Size-related shape change (Fig. 17) includes a relative decrease in frontal area length, particularly sagittally, accompanied by slight rearward migration of its widest point, slight negative allometry and abaxial migration of the palpebral lobe, and minor relative shortening of the occipital ring. According to sum squared calculation of predicted values from this regression, 28.64% of the cranidial shape variation is ontogenetic.

Figure 16. Partial Procrustes distance from the reference (mean shape of the smallest two cranidia) of 22 cranidial landmarks of W. vanhornei (N = 17).
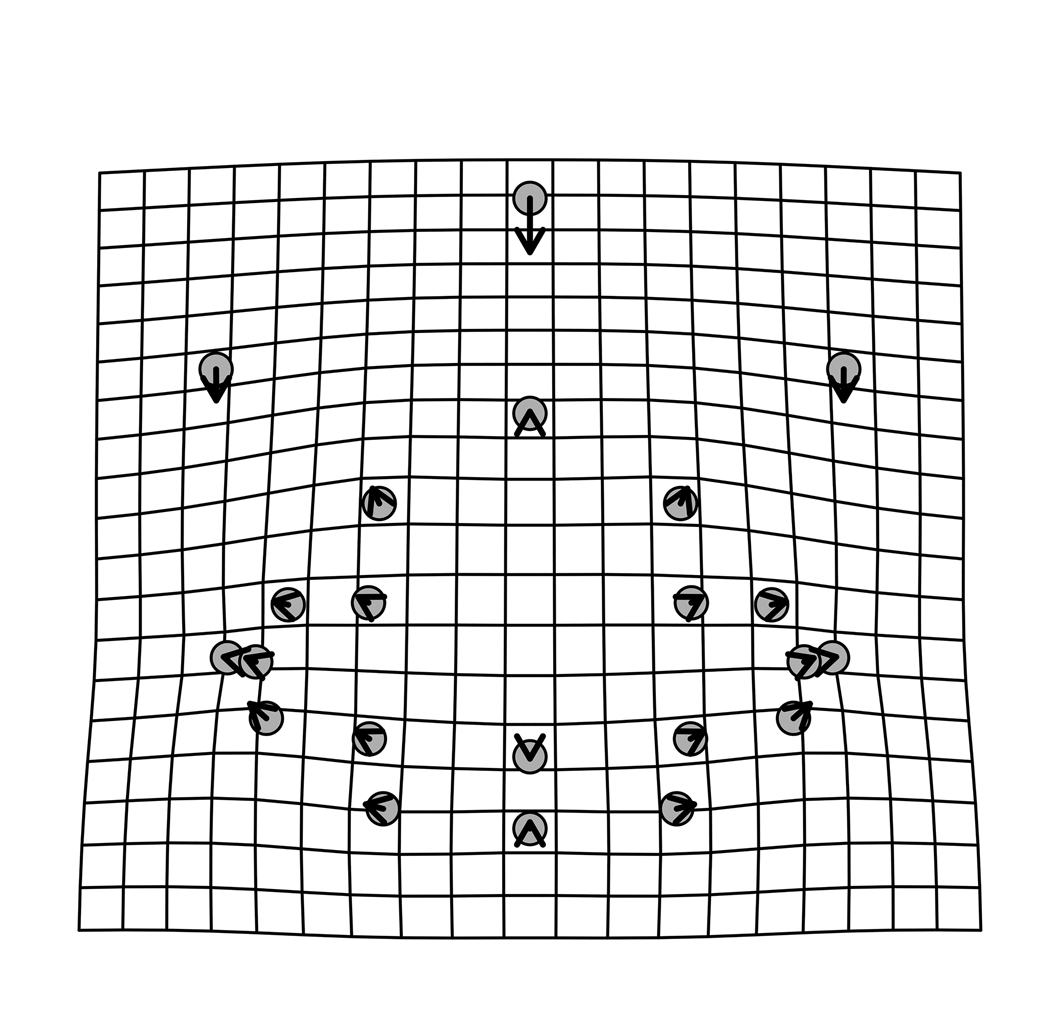
Figure 17. Thin-plate spline deformation grid of shape changes with growth for the 22 cranidial landmarks of W. vanhornei (N = 17).
Pygidial shape variation
The size range for the pygidium is from 2.00 mm to 5.50 mm in terms of total pygidial length (sag.).
We conducted three analyses of variation in pygidial shape for a total of 10 pygidia. One was a landmark-based analysis of landmarks on the axis and pygidial margin that also included landmarks representing both the adaxial and abaxial extents of the pleural and interpleural furrows. The second was similar but also included semilandmarks interpolated along the course of each of these furrows. A third analysis was limited to the axial landmarks only. In all cases, the smallest well-preserved pygidium specimen was used as the sole ontogenetic reference in this analysis because its centroid size was far smaller than that of all other specimens such that the normal procedure of averaging several specimens to produce a small-sized reference form was not feasible. In no case did we find evidence of significant allometry among the pygidia, although this result may reflect the fact that only 10 specimens were part of the analysis. Nevertheless, the analysis did highlight notable patterns of morphological variation among these pygidia. Given no evident support for ontogeny-related variation, these can be summarized together. The main results are: (1) The principal morphological variation among pygidia is related to the proportions of the pygidial terminal piece, the length of the postaxial region, and the location of the widest point on the pygidium. Together, these summarize the proportion of the pygidium occupied by the axis. (2) Despite some arbitrariness in exact placement of the adaxial extents of pleural and interpleural furrows due to differential flattening, their positions and courses are notably stable among W. vanhornei, and especially so in the better-defined, more-anterior furrows. (3) In specimens in which the terminal piece is expanded, the third axial ring is notably contracted sagittally.
Pygidial shape variation based on 40 landmarks.—These include landmarks marking the abaxial and adaxial margins of six furrow left-right pairs in addition to the landmarks on the axis and pygidial margin (Fig. 12.1).
A PCA of the partial warp scores shows that 71.60% of the shape variance is captured by the first three relative warps: RW 1, RW 2, and RW 3, respectively, explain 32.15%, 21.14%, and 18.31% of the total shape variance (Fig. 14.2). Each of the other six principal components contains less than 15.00% of total variance (Supplementary Data 4). RW 1 captures mostly variation in the position of the abaxial extent of the last interpleural furrow, and this may be influenced by differential flattening but also suggests some variation in the ratio of axial to postaxial length (Fig. 18.1). Other than that, RW 1 also captures variation in the relative width of the axis. RW 2 shows variation in the proportions of the axial terminal piece with respect to the third axial ring and in the relative length of the postaxial field (Fig. 18.2). RW 3 shows variation in the position of pygidial maximum width and in the relative length of the postaxial field (Fig. 18.3).
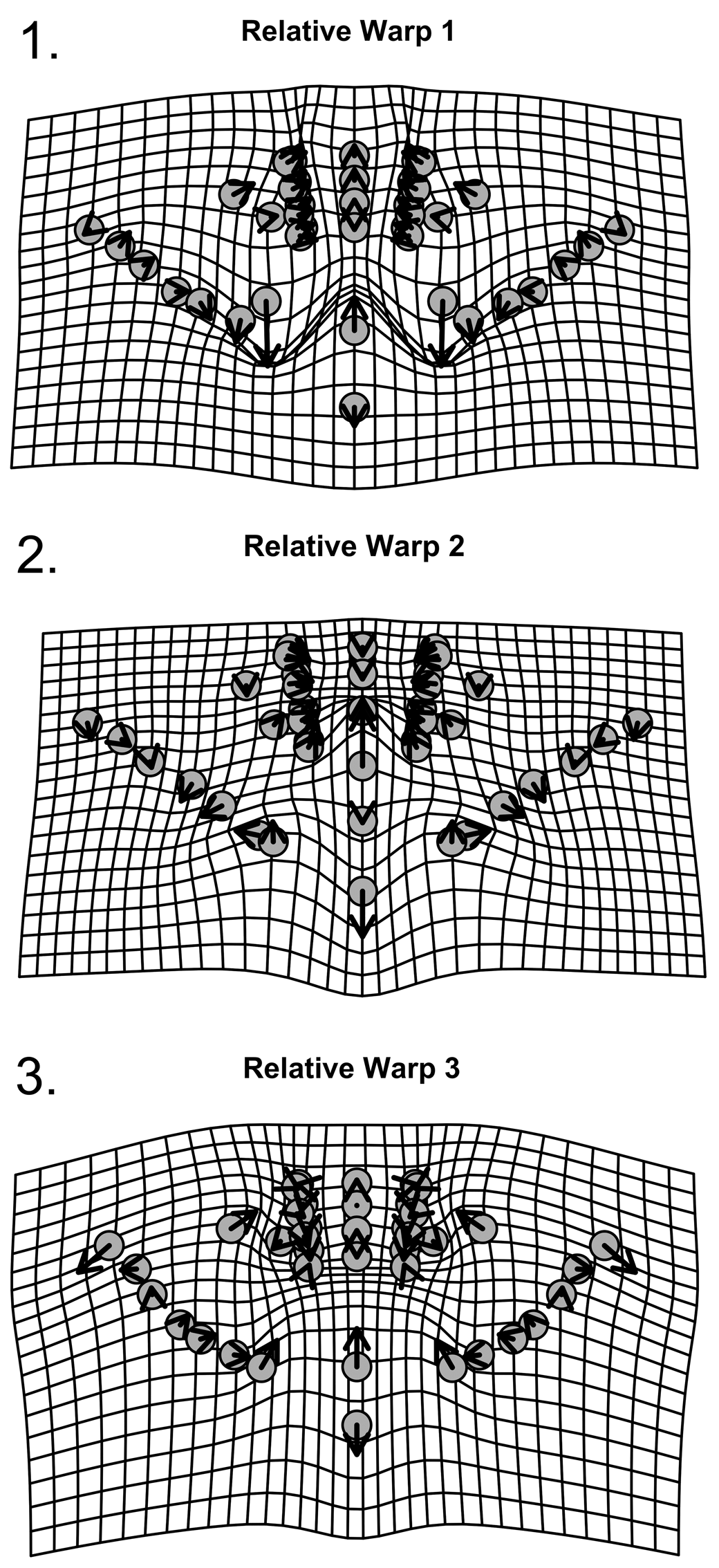
Figure 18. Thin-plate spline deformation grid of relative warps for 40 landmarks for pygidia of W. vanhornei (N = 10). (1) Shape variation related to RW1. (2) Shape variation related to RW2. (3) Shape variation related to RW3.
The relationship between partial Procrustes distance and ln CCS is not significant (slope = 0.02274, p = 0.72000, r = 0.14170) (Fig. 19). Regression of partial warp scores against ln CCS is not significant at p = 0.3726 (at 1,000 bootstraps) but captures 12.23% of the total variance. Accordingly, while limitations of the sample preclude exclusion of any role for ontogeny in generating observed variance among holaspid W. vanhornei pygidia, its influence did not dominate observed variation.

Figure 19. Partial Procrustes distance from the reference (mean shape of the smallest pygidium) of 40 pygidial landmarks of W. vanhornei (N = 10).
Analysis of pygidial semilandmarks.—The traces of furrows on the pleural region of Walcottaspis pygidium invite investigation via semilandmarks (18 semilandmarks distributed along each of 6 left-right paired pleural furrows, Fig. 12.2).
The PCA of general Procrustes aligned landmarks shows that 75.36% of the shape variance is captured by the first three RWs. RW 1 depicts 37.83% of the shape variance in the form of expansion of the postaxial region and change in the extent of the posteriormost pleural furrows, which is likely influenced partly by degree of compaction (Fig. 20.1). RW 2 contains 22.76% of the shape variance and expresses minor variation in positions of the pleural and interpleural furrows (Fig. 20.2). RW 3 (14.77% of shape variance) depicts expansion of the postaxial region and contraction of the third axial ring (Fig. 20.3). Each of the six additional RWs contains less than 10.00% of the shape variance (Supplementary Data 4).
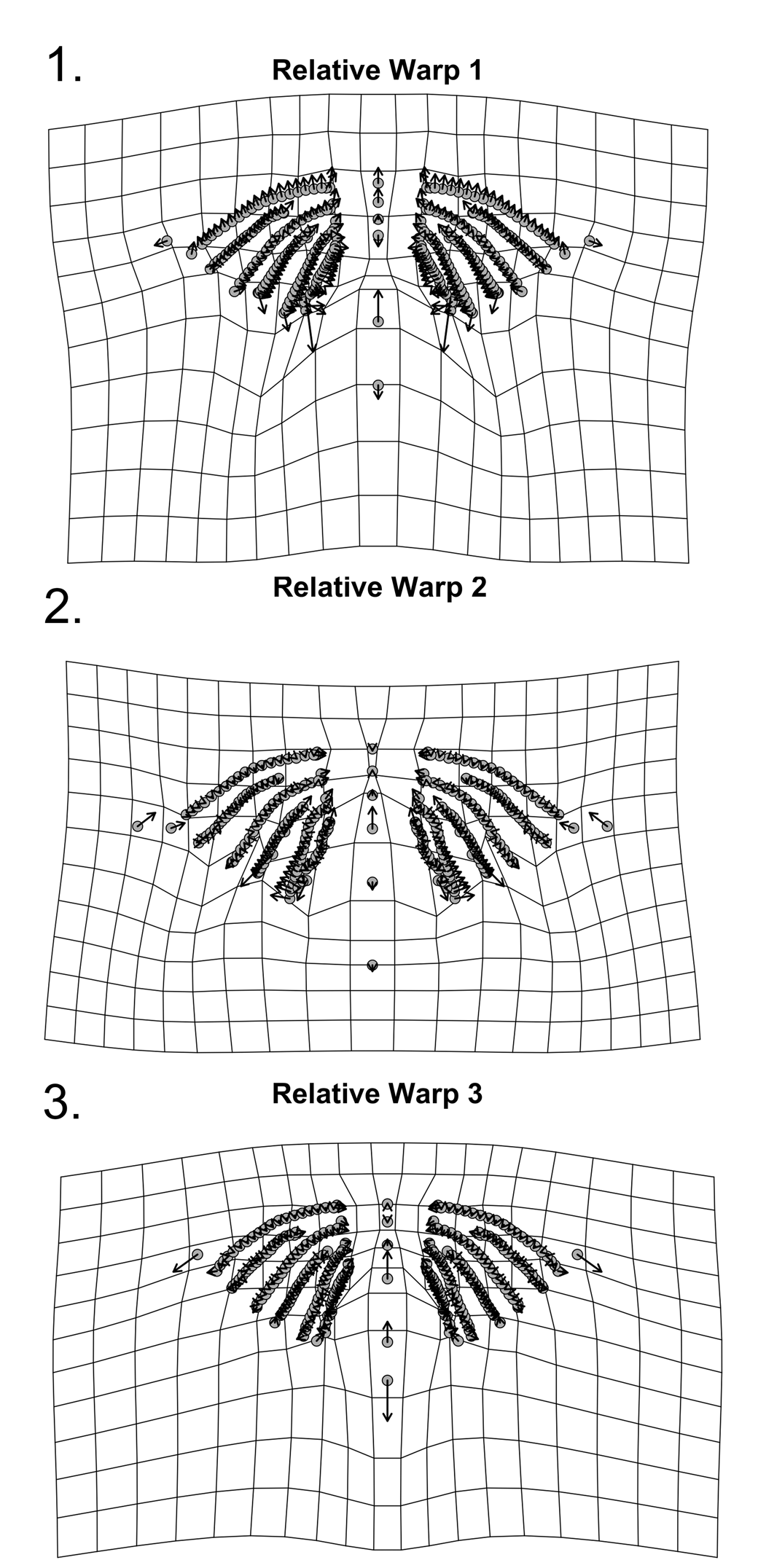
Figure 20. Thin-plate spline deformation grid of relative warps for 256 landmarks for pygidia of W. vanhornei (N = 10). (1) Shape variation related to RW1. (2) Shape variation related to RW2. (3) Shape variation related to RW3.
A nonsignificant negative relationship exists between partial Procrustes distance and ln CCS (slope = –0.02518, p = 0.2753, r = 0.4083) (Fig. 21). The regression of partial warp scores from the reference smallest specimen shape against ln CCS is also not significant (p = 0.4456 at 1,000 bootstraps).

Figure 21. Partial Procrustes distance from the reference (mean shape of the smallest pygidium) of 256 pygidial landmarks of W. vanhornei (N = 10).
Axial pygidial landmarks.—A subset of the previous data comprising 14 landmarks along the pygidial axis was chosen to explore variation therein (Fig. 12.3). The 10-specimen dataset was superimposed by generalized Procrustes analysis (Fig. 22).

Figure 22. Procrustes superimposition of 14 landmarks for pygidia including axis and posterior margin of W. vanhornei (N = 10).
The PCA of Procrustes aligned shape configurations shows that first two RWs capture almost the entire shape variance (84.38%): RW 1 captures more than half of the shape variance (54.61%,) and RW 2 explains 29.77% of the variance. Each of the remaining six RWs captures less than 15.00% of the shape variance in the dataset (Supplementary Data 4). RW 1 shows contraction of the length of the third axial segment while the posterior margin is extended and an overall contraction (tr.) of the axis is captured (Fig. 23.1). By contrast, RW 2 depicts expansion of the terminal axial piece, which decreases the relative length of the postaxial region (Fig. 23.2). The contraction of the third axial segment is consistently shown by both relative warps.

Figure 23. Thin-plate spline deformation grid of relative warps for 14 landmarks for pygidia of W. vanhornei (N = 10). (1) Shape variation related to RW1. (2) Shape variation related to RW2.
The relationship between partial Procrustes distance and ln CCS is not significant at p = 0.183 (slope = 0.2455, r = 0.488) (Fig. 24). When partial warp scores from the reference shape of the smallest specimen are regressed against ln CCS, the regression is not significant (p = 0.1598 at 1,000 bootstraps)

Figure 24. Partial Procrustes distance from the reference (mean shape of the smallest pygidium) of 14 pygidial landmarks of W. vanhornei (N = 10).
Overall, despite the low specimen abundance, W. vanhornei shows notable intraspecific variation in both the cranidium and pygidium, with most of this occurring in samples derived from a single locality, Hokah (HH), and there apparently from a single bed (Figs. 2, 14). The cranidium, known from more specimens, shows evident ontogenetic variation, something not confirmed for the pygidium. Pygidial morphological variation is nonetheless marked, particularly with respect to the length of the postaxial region, and again, this varies markedly within the collection made from the layer at about 6.6 m above the section base at Hokah (Fig. 14.2).
Materials
All specimens of W. vanhornei came from the heterolithic facies of the St. Lawrence Formation and are from Hokah, Minnesota, unless otherwise specified.
Repositories and institutional abbreviations
Types, figures, and other specimens are reposited in the following institutions: University of Minnesota Paleontological Collection (UMPC, now part of the Cincinnati Museum Center [CMCIP] collection); Museum of Comparative Zoology, Harvard (MCZ); Field Museum of Natural History, Chicago (FMNH); University of Wisconsin, Madison (UWGM); United States National Museum of Natural History (USNM), and Yale Peabody Museum (YMP).
Systematic paleontology
In the following, we employ the synonymy list notations of Matthews (Reference Matthews1973).
Class Trilobita Walch, Reference Walch1773
Subclass Libristoma Fortey, Reference Fortey1990
Order Asaphida Salter, Reference Salter1864; emended Fortey, Reference Fortey1990
Superfamily Dikelocephalacea Miller, Reference Miller1889 (emended Ludvigsen, Westrop, and Kindle, Reference Ludvigsen, Westrop and Kindle1989)
Family Dikelocephalidae Miller, Reference Miller1889; emended Ludvigsen, Westrop, and Kindle, Reference Ludvigsen, Westrop and Kindle1989)
Remarks
While the higher-level systematic relationship among members of this family and with its relatives will be discussed by us elsewhere, here we reject Ulrich and Resser's (Reference Ulrich and Resser1930) concept of Osceolinae because our phylogenetic analysis does not support a sister-taxon relationship between W. vanhornei and species of Osceolia. We consider W. vanhornei to be part of a clade containing those dikelocephalids with an extended cephalic doublure (see the preceding) and thus not part of the paraphyletic subfamily Dikelocephalinae as conceived of by Ludvigsen et al. (Reference Ludvigsen, Westrop and Kindle1989, p. 28), which contains taxa traditionally considered to be saukiid.
Genus Walcottaspis Ulrich and Resser, Reference Ulrich and Resser1930
Type species
Walcottaspis vanhornei (Walcott, Reference Walcott1914) from the heterolithic facies of the St. Lawrence Formation.
Diagnosis
As for type species by monotypy.
Walcottaspis vanhornei (Walcott, Reference Walcott1914)
Figures 3, 5, 6
- v*Reference Walcott1914
Dikelocephalus vanhornei Walcott, p. 373, pl. 62, figs. 1–3.
- v*Reference Ulrich and Resser1930
Walcottaspis vanhornei Ulrich and Resser, p. 65, pl. 20. figs. 3–5
- Reference Twenhofel, Raasch and Thwaites1935
Walcottaspis vanhornei Twenhofel, Raasch, and Thwaites, p. 1712
- Reference Richter and Richter1940
Walcottaspis vanhornei Richter and Richter, p. 24, pl. 25, fig. 4.
- Reference Raasch1951
Walcottaspis vanhornei Raasch, p. 141, 150.
- Reference Harrington, Henningsmoen, Howell, Jaanuson, Lochman-Balk and Moore1959
Walcottaspis vanhornei Lochman in Harrington et al., p. 254, fig. 191, 1a.
- non v*Reference Shah, Parcha and Raina1991
Walcottaspis sp. Shah et al., p. 91, pl. 1, figs. j, n.
Lectotype
Selected herein: pygidium FMNH-UC14393-b from heterolithic facies of St. Lawrence Formation, Hokah (HH), Minnesota (Walcott, Reference Walcott1914, pl. 62, fig. 3).
Paralectotypes
Cranidium (FMNH-UC14393-a) and thoracic segment (FMNH-UC14393-c) (Walcott, Reference Walcott1914, pl. 62, figs. 1, 2); paralectotypes from the same bed as lectotype at Hokah (HH), Minnesota.
Diagnosis
The S1 glabellar furrow isolated, transversely weakly incised, effaced in largest specimens. The S2 and additional glabellar furrows absent. Fixigena wide relative to other derived dikelocephalids. Pygidium with three clearly incised axial rings and long (sag.) terminal piece. Anterior pygidial margin fulcrate, transverse adaxially; posterior margin elliptical. Interpleural furrows of first two segments abruptly obsolete within pleural platform before approaching axial furrow. Postaxial region of variable length, but short (sag.).
Occurrence
Eoconodontus Zone, heterolithic facies of the St. Lawrence Formation within narrow outcrop belt including Hokah, Winona, and Reno, Minnesota, and Bluff Siding, Wisconsin.
Description
Mature exoskeletons large, largest likely longer than 50 cm sagittally. Cephalon convex, outline semicircular, pygidium lateral margin shape matches that of anterior cephalon.
Glabella quadrate in smaller holaspids, trapezoidal in largest holaspids, inflated, maximum width just posterior to S1 furrow, expressed as slight bulge in L1. Glabellar anterior margin rounded. SO deepest adaxially, isolated in some specimens, shallowing transversely to yield slit-like furrow appearance. S1 isolated, variably incised but deepest axially where it forms an open U shape, running obliquely slightly forward abaxially. S2 obsolete. Median occipital tubercle present. Low, semicircular to transverse ridge shortly anterior to posterior margin of occipital ring. Axial furrow sharply incised. Frontal area long (sag.), broad (tr.), not less than 35% of occipital-glabellar length, flat to downsloping. The occipital lobe width is not less than 46% of the frontal area width. Anterior margin rounded to slightly angular axially, border absent. Frontal area width (tr) wider than maximum cranidial width between palpebral lobes (tr.) in smaller specimens, equal in larger. Opisthoparian dorsal cephalic suture marginal sagittally, laterally sweeps posteriorly at an angle of 20–41° to sagittal axis. Fixigenae wide for family, narrowest preocularly, shortly anterior to S1 where wider (tr.) than occipital lobe length (sag.). Intraocular fixed cheek inflated against palpebral furrow, sloping downward toward axial furrow (tr.). Palpebral lobes relatively short (exsag.) and narrow (tr.), midpoint opposite L1 bulge, relatively longer in smaller specimens. Palpebral furrow crescentic, deeply and evenly incised, eye ridge obsolete but ocular platform inflated to height of palpebral lobe distally. Posterior border area spatulate, terminating in blunt point abaxially, abaxial posterior portion with transverse ridge that parallels course of posterior border furrow. Posterior border furrow evenly incised, anterior face steeper than posterior, running transversely and posteriorly from point where SO projects to meet axial furrow.
Free cheeks arcuate, conjoined. Broad, undivided genal field, gently convex near the margin, rising steeply concavely toward flat ocular platform. Posterior border furrow sigmoidal, curving posteriorly into base of narrow, long genal spine. Ocular incisure arcuate, bounded adaxially by shallow arcuate furrow and raised marginal flange forming eye socle. Terrace ridges on ocular platform inosculate. Doublure wide, extending about nine-tenths of distance from margin to ocular incisure, with flexure in its anterior portion that accommodates the anterior edge of hypostome. Doublure curves upward adaxially, remaining close beneath dorsal surface. Terrace ridges on doublure straight.
Hypostome subquadrangular, wider (tr.) than long (sag.); median body large, oval, inflated; anterior lobe convex, round; posterior lobe short (sag.), wide (tr.). Median furrow pit-like abaxially, shallow axially; maculae oval, small; pit-like furrows are developed at posterolateral margin of posterior lobe; border furrow weakly incised; lateral border triangular, widest (tr.) opposite midpoint of posterior lobe. Posterior border lip-like, gently arched anteriorly in axial portion. Terrace ridges on lateral border straight to sinuous, cuestaform, steep slopes facing adaxially.
Axial region of thoracic segments convex, articulating furrow transverse. Margins of thoracic pleurae transverse adaxially from axial furrow, curving posteriorly abaxial of fulcrum; anterior facet long (exsag.) abaxially of fulcrum; pleural furrow diagonal, firmly incised adaxially, reaching almost to tip of pleura; propleura and opisthopleura of equal length (exsag.) at fulcrum; pleural spine tip blunt. Fulcrum no more abaxial than one-third (tr.) of pleural width.
Pygidium anterior margin transverse adaxially, extending posterolaterally abaxial to fulcrum; posterior margin entire, semielliptical. Axis convex, 60–72% of pygidial length (sag.) with anterior articulating half-ring, three distinct axial rings defined by narrow and well-incised inter-ring furrows, long (sag.) terminal piece. Ridge arches anteriorly from posterior border of first axial ring, occupying medial three-quarters of ring width (tr.) and extending across one-half of ring length (sag.) Each axial ring with pair of swollen nodes located anterior of segment midline (exsag.) near axial furrow; two additional pairs of the lobes extend onto terminal piece in lectotype, which also shows one additional incomplete rachial furrow. Terminal piece may be inflated, posterior margin blunt, rounded to bullet-shaped. Pleural furrows entire, extending to near margin in anterior segments; interpleural furrows of first two segments abruptly obsolete on pleural platform approaching axial furrow; third interpleural furrow connects to axis at posterior of third axial ring. Postaxial region short (sag.) but of variable length. Petaloid terraces developed on articulating facet of first pleura. Doublure extends to pleural platform lying close beneath dorsal surface, with faint paradoublural line, terrace ridges on it straight, relief high, steeper slopes facing outward; some 25 terraces from top of pleural platform to margin.
Other material
All from Hokah, Minnesota, unless otherwise stated. Twenty-one cranidia (FMNH-PE-39216-c, FMNH-PE-39214, FMNH-PE-39215, FMNH-UC-23314-a, FMNH-UC-23314-b, FMNH-UC-23314-c, FMNH-UC-23314-d, CMC-IP-87565x, CMC-IP-89411h, CMC-IP-89411i, CMC-IP-87565j, CMC-IP-89411m, CMC-IP-87565n, CMC-IP-87565t, CMC-IP-95630, UWGM 7150, UWGM 7151, UWGM 7152, UWGM 7155, UWGM 7156, YMP73152), 19 pygidia (FMNH-PE-39216-a, FMNH-PE-39216-b, FMNH-PE-39210, FMNH-PE-39312, FMNH-UC-23314-e, FMNH-UC-14393-b, MCZ-IP910 from Bluff Siding, Wisconsin, CMC-IP-89411a, CMC-IP-89411b, CMC-IP-89411c, CMC-IP-87565d, CMC-IP97568, USNM-PAL-72687A from Winona, Minnesota, USNM-PAL-72687B from Winona, Minnesota, UWGM 7157, UWGM 7158, UWGM 7159, UWGM 7160, UWGM 7161), five disarticulated free-cheek specimens (FMNH-UC-23314-h2, FMNH-UC-23314-h, FMNH-UC-23314-h1, FMNH-UC-23314-i, CMC-IP89411y), three hypostomes (FMNH-UC-23314-f, FMNH-UC-23314-f, FMNH-UC-23314-g), one conjoined free cheek (MCZ-IP-917 from Bluff Siding, Wisconsin), and two thoracic segment specimens (FMNH-PE-82162, FMNH-UC-14393-c). Other previously figured material includes one pygidium (USNM-PAL-72687A) from Winona, Minnesota (Ulrich and Resser, Reference Ulrich and Resser1930, pl. 20, fig. 6; Fig. 5.4) and one plastotype cranidium (USNM-PAL 58608-a) and one plastotype pygidium (USNM-PAL 58608-b) cast from Walcott's (Reference Walcott1914) figured specimens (FMNH-UC-14393-a and FMNH-UC-14393-b; Ulrich and Resser, Reference Ulrich and Resser1930, pl. 20, figs. 3, 4).
Remarks
The proportions of the posterior trunk in our reconstruction are based on a pygidial size in which the marginal outline best matches the shape of the anterior cephalic doublure, which it does closely and that may also suggest spheroidal enrollment in this trilobite. Proportioning the pygidium in this way and assuming 12 thoracic segments in maturity, as known in D. minnesotensis, yields an overall body plan that seems realistic (Fig. 6.2). However, the number of thoracic segments in holaspid W. vanhornei is unknown. Given that W. vanhornei has one fewer clearly defined holaspid pygidial segment than D. minnesotensis, one possibility is that the total number of trunk segments was common to both species and that the evolution of W. vanhornei from a minnesotensis-like ancestor reflected release of the leading pygidial segment into the mature thorax (see Hughes, Reference Hughes1994, p. 53). If the pygidial spines in D. minnesotensis were related to this leading segment, then the striking morphological difference between their pygidia might be explained without invoking major body repatterning because the two species may have had quite similar trunk forms, their principal difference being where the pygidial boundary was situated. Although this suggestion is speculative, it illustrates how the trilobite trunk region, with its variations not only in the form and degree of integration between segments but also in their overall number and allocation to the mature thorax or pygidium, offered more “degrees of freedom” to vary than the cephalon and may explain, in part, why the cranidia of W. vanhornei and D. minnesotensis remain much more similar than their pygidia. However, in the posterior of the thorax of D. minnesotensis, segments lost the fulcrum and became markedly more siculate, whereas in W. vanhornei, the fulcrum was maintained throughout the trunk as it is present in both the holaspid posterior cephalic and anterior pygidial margins. This suggests a lower gradient of thoracic segment shape change in W. vanhornei than in D. minnesotensis. A number of determinants of trunk structure were evidently adjusted during the evolution of W. vanhornei.
Walcott's (Reference Walcott1914, pl. 62, fig. 3) illustration of the lectotype pygidium of W. vanhornei was “retouched,” an ink-based outlining technique used at will to emphasize subtle features. His figure suggests that the terminal piece bears possibly as many as four transverse furrows in addition to those defining the three clear axial segments. These additional furrows are not visible in the image of the same specimen provided by Ulrich and Resser (Reference Ulrich and Resser1930, pl. 20, fig. 4), but our illustrations (Figs. 4.1, 5.7) confirm some transverse structures within the terminal piece. Recognition of an incompletely expressed fourth segment is warranted on the basis of the nodes and furrow expressed, and there may be a fifth set of nodes, but we do not see evidence for all the four additional segments that Walcott (Reference Walcott1914, fig. 3) depicted. Furthermore, furrows expressed in the pleural region relate only to the first three axial segments. Such imprecision in segment counts is not surprising. The terminal piece housed the zone of teloblastic cells from which new segments derived, and trilobites varied in the ways in which new trunk segments were expressed with, for example, mismatches known between the numbers of ventral appendages and segments expressed in the exoskeleton (Hughes, Reference Hughes2003). However, there is no evidence of a change in the number of well-expressed segments during the part of the holaspid ontogeny of W. vanhornei available for analysis: it is three in all specimens, as opposed to the four segments clearly expressed in both axis and pleural region in most D. minnesotensis, a species that also has additional, indistinctly expressed segments in its terminal piece. Accordingly, we consider the two species to differ in the number of firmly expressed pygidial segments and this difference between them to be not simply variation in the prominence with which the most posterior segments are expressed.
The distinctive elliptical pygidial shape of W. vanhornei is homeomorphic with that of several other trilobites. Examples from the same region include Briscoia pertransversa Lochman (Reference Lochman1956) from the Prosaukia Zone of the Tunnel City Group and likewise Wilbernia sp. occurring in the “Hudson Member” of the Lone Rock Formation in lower stratigraphic units (Fig. 10). Both these other examples also bear three clearly expressed segments in the axis and matching pleural and interpleural furrows that mimic those of the thorax (i.e., all trunk segments are homonomous in structure; Hughes, Reference Hughes2003). Such pygidia must link articulating thoracic segments to the terminal piece across a short interval of three pygidial segments. If the postaxial region is short, then an elliptical shape for the pygidial posterior shape is required to produce the necessary pleural width at the anterior of the pygidium to allow integration with the thorax. Accordingly, it is not unexpected that pygidial shape and furrow structure arose iteratively in various groups at that time. However, W. vanhornei is clearly distinguished from such homeomorphs by its unique termination of the anterior interpleural furrows and by the features that place it as sister taxon to D. minnesotensis, which bore a more segment-rich pygidium.
Acknowledgments
We thank J.D. Cundiff, C.A. Eaton, M. Florence, B.R. Hunda, C.C. Labandeira, P. Mayer, C.E. Schwalbach, and R. Slaughter for access to collections in their care and fondly recall the help of M. Carmen, F.J. Collier, B.R. Erickson, C.A. Foster, R. Johnson, R. Pody, W. Rice, P.M. Sheehan, R.E. Sloan, and K. Westphal with collections access in the 1980s. Fieldwork was undertaken with the help of G.O. Gunderson and R.E. Meyer and funded by a UK Natural Environmental Research studentship to N.C.H., advised by D.E.G. Briggs, and more recently by awards to S.S. by the American Association of Petroleum Geologists, the Paleontological Society, the Indo-US Science and Technology Forum, and the University of California – Riverside Mike Murphy fund. Insightful reviews by L. Laibl, J.F. Taylor, and associate editor B.S. Lieberman much improved the work. We especially thank M.J. Hopkins of the American Museum of Natural History for running the Bayesian analysis and E. Vargas-Perra, A.C. Runkel, and H.D.S. Sheets for essential discussion of setting and analysis.
Declaration of competing interests
The authors declare none.
Data availability statement
Data available from the Dryad Digital Repository: https://doi.org/10.5061/dryad.h70rxwdpd.


























 Open access
Open access