



Introduction
The 77,950 km2 Ouadi Rimé–Ouadi Achim Reserve (Réserve de l'Ouadi Rimé–Ouadi Achim) in central Chad is one of only a few protected areas, and by far the largest, representing Africa's vast Sahelian grassland biome (Brugiere & Scholte, Reference Brugière and Scholte2013; Protected Planet, 2020). When legally established as an IUCN category IV reserve in 1969, the objective was to protect significant populations of threatened Sahelo–Saharan endemic species (Newby, Reference Newby1978) such as the scimitar-horned oryx Oryx dammah (categorized on the IUCN Red List as Extinct in the Wild; IUCN/SSC Antelope Specialist Group, 2016c), dorcas gazelle Gazella dorcas (Vulnerable; IUCN/SSC Antelope Specialist Group, 2017), dama gazelle Nanger dama (Critically Endangered; IUCN/SSC Antelope Specialist Group, 2016b) and addax Addax nasomaculatus (Critically Endangered; IUCN/SSC Antelope Specialist Group, 2016a). The Reserve also supported the North African ostrich Struthio camelus, Arabian Ardeotis arabs, Nubian Ardeotis nuba and Denham's Neotis denhami bustards, and small populations of the West African cheetah Acinonyx jubatus and African wild dog Lycaon pictus. Traditional transhumant livestock management, originally low impact and highly seasonal, has always been permitted in the Reserve, but hunting and tree cutting are banned. The populations of oryx, addax and ostrich were extirpated during a period of civil unrest in the late 1970s and early 1980s (Newby, Reference Newby1978, Reference Newby1980, Reference Newby, Dixon and Jones1988). The scimitar-horned oryx was first categorized as Extinct in the Wild on the IUCN Red List in 2000. Since the 1970s, as elsewhere (De Leeuw & Tothill, Reference De Leeuw and Tothill1990), development and anthropogenic pressures such as density of people and livestock, water use, grazing pressure, disturbance, vehicle traffic and grass fires have increased in the Reserve.
Following the decision of the Government of Chad to support the restoration of the scimitar-horned oryx and other wildlife populations (Bemadjin et al., Reference Bemadjim, Newby, Desbiez, Lees and Miller2012), we assessed numbers and spatial interactions between dorcas and dama gazelles and livestock in the Reserve. This study is based on transect surveys conducted to assess the feasibility of reintroducing the oryx to the area, and continued during the initial stages of this reintroduction. We report on trends in densities and distribution of the gazelles and livestock, based on data from eight large-scale, vehicle-based line transect surveys conducted in the core of the Reserve during 2011–2019. Our aim was to determine how the presence of livestock, artificial water sources and bush fires affects the distribution of wild ungulates. With oryx (Duthie et al., Reference Duthie2018), addax and ostrich reintroductions underway and steps taken to support the dama gazelle in the Reserve, we use evidence from the surveys to recommend management actions to ensure that both conservation and livestock sector objectives are achieved.
Study area
The study area is in the central Ouadi Rimé–Ouadi Achim Reserve, Chad (Fig. 1). The nearest towns are Arada (80 km to the east) and Biltine (117 km to the east-south-east). The habitat is Sahelian grassland on flat grey soils overlain in a few places by vegetated fixed dune formations. Altitude descends from 400 m in the east to 374 m in the west of the 70-km wide study area. The shallow gradient on flat land creates diffuse run-off along broad seasonal drainage lines (locally called ouadis). There are no significant hills or rocky outcrops, but vegetated low dunes form a belt of undulating sandy substrates across the centre of the study site.

Fig. 1 Location of the study area in the Ouadi Rimé-Ouadi Achim Reserve, Chad, and details of the line transect and reconnaissance zones in the study area. We used eight core line transect routes in 2015–2019 (solid lines) and four reconnaissance transect routes in 2017 (dashed lines). An additional seven line transects were interleaved simultaneously in parallel to the primary route in 2019 only (dotted line). Natural water hole depressions that have been artificially enlarged (hafiris), major drainage lines (ouadis) and main access routes to the release site of the scimitar-horned oryx Oryx dammah are also shown.
The mean temperature is 35 °C (range 21–47 °C) in the hottest month, May, and 22 °C (9–37 °C) in the coolest month, January. In the driest month, April, relative humidity averages 9% (1.7–27.5%), and 72% (27.8–99.9%) in the most humid month, August. The annual passage of the Intertropical Convergence Zone creates a single wet season during July–September, with occasional dust storms as winds change from north-easterly, bringing humid air from the south-west. Mean annual rainfall at the study site during 2017–2020 was 190 mm (139–273 mm). Typically, there are no natural water sources in the study area for 9 months of the year, but following rainstorms, which are often local in distribution, small temporary water holes last for a few days or weeks. Within the ouadis, longer-lasting chains of shallow, temporary lakes (called rahads) may form, which typically measure 50–100 m across. In the east of the study area some of these have been artificially enlarged to create so-called hafiris, providing water for increased numbers of livestock and locally prolonging the period of grazing pressure and human occupation further into the dry season (Fig. 1).
The grasslands are dominated by annual grasses, of which Aristida mutabilis Trin. & Rupr., Aristida funiculata Trin. & Rupr., Dactyloctenium aegyptium (L.) Willd., Schoenefeldia gracilis Kunth., Cenchrus biflorus Roxb. and Brachiaria sp. (Trin.) Griseb are important for ungulates. On sandy soil the perennial Panicum turgidum Forssk. is also important, but now appears to be less abundant than formerly (Gillet, Reference Gillet1961; JN, pers. obs., 2007–2020). Following rain, the legume Indigofera colutea (Burm f.) Merrill (selected by gazelles and oryx) forms dense local patches, and stands of the bushy perennial Chrozophora brocchiana (Vis) Schweinf. are characteristic of drainage depressions. The bitter melon Citrullus colocynthis (L.) Schrad. is widespread, and the water content of its leaves and fruit is important for wildlife and livestock. Following the short growing season, from late September the grasses and herbs desiccate to form dense, dry hay and the land becomes vulnerable to fire.
Shade is provided mainly by desert date trees Balanites aegyptiaca (L.) Delile, growing as widely scattered individuals or in small thickets, and less commonly by Acacia tortilis (Forssk.) Hayne, which often indicate seasonal water collection points. On some soil types scattered stands of shrubbier Capparis decidua (Forssk.) Edgew. and Boscia senegalensis (Pers.) Lam. predominate.
Methods
Surveys
The aim of the eight surveys conducted during 2011–2019 was to establish baseline estimates of densities and relative distribution of wild and domestic ungulates in the central Reserve. The survey area is a small part of an open system across which dorcas and dama gazelles and livestock move freely. The term ‘population’ thus refers here only to the animals using the study area at the time of a survey.
Surveys used standard line transect methods (Buckland et al., Reference Buckland, Anderson, Burnham, Laake, Borchers and Thomas2001; Young et al., Reference Young, Murray, Strindberg, Buuveibaatar and Berger2010; Baamrane, Reference Baamrane, Znari, Loggers, El Mecht and Naimi2013; Le Moullec et al., Reference Le Moullec, Pedersen, Yoccoz, Aanes, Tufto and Hansen2017). The survey zone covered c. 3,500 km2 (3% of the Reserve) and was selected as an area of relatively low human activity, with no local development of water wells, and a representative mix of vegetated fixed dune and floodplain habitats in the core of the Reserve. This included the only area where the Critically Endangered dama gazelle was reported to be regularly observed. Transect orientation and the central survey area were consistent throughout the study, but transect layout varied slightly during the initial exploratory phases in 2011–2013, before the design was fixed once longer-term funding was assured (Table 1). During 2015–2019, surveys used a standard core framework of seven fixed transects, spaced 10 km apart (Fig. 1). A continuous line transect design improved efficiency. Transects were orientated to traverse low dune habitats running east to west across the survey zone and extended into floodplain habitats either side. In the latter four surveys, an east–west transect was added to the south within the floodplain of Ouadi Haddat (Fig. 1).
Table 1 Summary statistics and results from eight line transect surveys of the central zone of the Ouadi Rimé–Ouadi Achim Reserve, Chad, 2011–2019.

1CV, coefficient of variation.
2Tropical livestock unit, equivalent of mature individual weighing 250 kg (see text for details).
In the 2017 wet and dry seasons, we extended the systematic assessment of wild and domestic ungulate distribution by surveying an additional 40 × 50 km contiguous reconnaissance survey block to the north of the line transect study zone (Fig. 1). In the reconnaissance block, data were collected as counts only, by two teams in separate vehicles working on parallel transects 10 km apart. Species encounter rates (per 10 × 10 km grid cell) of the reconnaissance survey block and line transect zone were combined to generate distribution maps of wild ungulates and livestock. In the final line transect survey (2019), an additional set of transects was inserted between the established transects to create 5-km transect spacing, driven by two teams working in parallel, to increase sampling effort to 15 transects and improve precision of counts and density estimates (Bårdsen & Fox, Reference Bårdsen and Fox2006; Strindberg et al., Reference Strindberg, Buckland, Thomas, Buckland, Anderson, Burnham, Laake, Borchers and Thomas2010).
Because dama gazelles were encountered only a total of 13 times during all surveys, we also analysed the distribution of 78 additional, opportunistic observations of this species collected during 2–3 month periods of continuous field work in the 2016 wet season and the following dry season in 2017. In these periods we recorded all observed wildlife and livestock, and all routes driven, and mapped perimeters of bushfires. We then overlaid locations of dama gazelle observations and fires on livestock encounter rate heat maps, using a 10 × 10 km grid in Surfer 11.6.1159 (Golden Software, Golden, USA).
On line and reconnaissance transects, survey routes prepared with GPS software (Mapsource 6.16.3 and Basecamp 4.7.2; both Garmin Ltd., Olathe, USA) were displayed on dashboard mounted units. In the open terrain, the vehicle remained mostly well within 25 m of the planned transect line, with an average speed of 12–15 km/h.
We recorded wildlife and livestock observations as groups using Cybertracker 3 (Cybertracker Conservation, 2019). Dorcas and dama gazelles often move away on being sighted. To fulfil distance sampling assumptions, we selected landmark reference points (isolated trees, small shrubs, grass tussocks or bare patches of ground) near the central position of herds when first seen. We then measured the distance from the transect line to these landmarks with a laser rangefinder once the vehicle was positioned perpendicular to the landmark. At right angle turns, observations originating in the external quadrant of the turn were excluded and observations originating in the interior quadrant were not double counted.
Livestock (camels, sheep, goats, cattle, horses and donkeys) were counted (or estimated for large, dense herds) individually by species, and distances measured to the central location of the assembly. We converted these counts to tropical livestock units (equivalent to a mature animal weighing 250 kg), using correction factors: 1.25 for camels, 0.7 for cattle (in herds), 0.1 for sheep, 0.08 for goats, 0.5 for donkeys and 1 for horses (Le Houerou & Hoste, Reference Le Houerou and Hoste1977; FAO, 2011), to provide standardized measures. We used 16 kg as the unit weight for the dorcas gazelle (Yom-Tov et al., Reference Yom-Tov, Mendelssohn and Groves1995).
We obtained satellite data on locations of fires within the survey area from ESDS (2020). As a proxy for the impact of fires, we summed the number of detected fires in each 10 × 10 km grid cell over 1–60 days and 61–180 days prior to the start date of each survey.
We required 3–5 days to complete each survey, operating during 07.30–17.00 each day, with a break during 12.00–14.00. Detectability of animals may thus have varied over the course of a survey as animal activity changes with time of day, cloud cover and temperature, but all surveys were conducted with as much standardization as was feasible.
Data analysis
We calculated estimates for numbers and densities of dorcas gazelles and livestock using Distance 6.0 (Research Unit for Wildlife Population Assessment, University of St Andrews, St Andrews, UK). Uniform, half-normal and hazard rate key functions were compared, with cosine, simple polynomial and hermite polynomial adjustment terms constrained, to ensure the detection function decreased monotonically. We used size-bias regression to account for larger groups being more likely to be sighted at longer distances than smaller groups, and maximized sample size by using a global detection function across all surveys, with truncation at 400 m for the smaller dorcas gazelle and 490 m for livestock. We used Akaike's information criterion (AIC; Sakamoto et al., Reference Sakamoto, Ishiguro and Kitagawa1986) to select the best models, and confirmed an acceptable fit to the data using a χ 2 test.
We assessed the trend of the dorcas gazelle population for 2013–2019, when the sample design was standardized, by regressing the natural log of the density estimates against survey year in R 3.4.4 (R Core Team, 2019). We used density rather than estimated numbers because of variations in survey area (Table 1). We performed a Bayesian analysis (Crome et al., Reference Crome, Thomas and Moore1996) to estimate the probability of decline or increase, assuming a flat prior and treating the scaled likelihood curve as the posterior probability.
We examined the distribution of livestock and dorcas gazelles in relation to artificial water sources by calculating minimum linear distance from the centre of each 10 × 10 km transect grid cell to the nearest artificial water point using the NNJoin package in QGIS 3.12 (QGIS, 2020). There were three types of artificial water sources (in order of increasing cost to use them): hafiris (natural water hole depressions, artificially enlarged to prolong surface water availability), cement-lined wells (hand-drawn using livestock) and boreholes (with mechanized pumps). We modelled dorcas gazelle encounter rate in each grid cell against livestock encounter rate, distance to nearest water point of each type, season, survey and fire detections in the recent and longer-term period prior to a survey, using multiple regression in R 3.4.4 (R Core Team, 2019). Livestock encounter rate was modelled against distance to nearest water point of each type, season, survey, and recent and longer-term fire detections. We used AIC for model selection. To examine sampling efficiency, we used observed encounter rates to predict sample effort required (total transect line length) in relation to targeted level of precision, measured as the coefficient of variation (CV; Buckland et al., Reference Buckland, Anderson, Burnham, Laake, Borchers and Thomas2001).
Meteorological data for the study site were obtained from a HOBO 21-USB weather station established at the oryx release site base camp in 2017, as no other data close to the study area were available.
Results
Dorcas gazelle numbers and trend
Estimated numbers of dorcas gazelles in the line transect study zone ranged from c. 7,700 to c. 18,000 individuals; associated mean densities were 3.5–7.0 dorcas/km2 (Fig. 2a). Precision (CV) averaged 18% (11–29%) across all surveys (Table 1). The large differences between maximum and minimum numbers correspond with the movement patterns of the species in this area, with some seasonality in reproduction, although calves can be found in all months. Average dorcas density during the wet season was marginally lower than during the dry season (Table 1). Bayesian analysis showed an 81% probability of decline over 2011–2019 (Fig. 3a), influenced by high densities in the smaller scale 2011 surveys. Analysis across the latter six standardized surveys (2015–2019) showed a positive trend, with 95% probability of increasing numbers (Fig. 3b).

Fig. 2 Estimated densities of (a) dorcas gazelles Gazella dorcas and (b) livestock (TLU, tropical livestock units) in the study area during 2011–2019. The vertical lines represent the 95% CI. Note the order of magnitude difference in y-axis scale and unit body weights (c. 16 kg/dorcas individual vs 250 kg/tropical livestock unit).

Fig. 3 Bayesian posterior probability distributions of trend in numbers of dorcas gazelles using the line transect sample zones across (a) all eight surveys (2011–2019) and (b) the latter six surveys (2013–2019), when the transect layout was standardized.
Livestock numbers
Mean estimates for livestock numbers ranged between c. 2,000 and c. 68,000 tropical livestock units, with low precision (CV 30–74%; Fig. 2b, Table 1). The low precision was associated with large variation in encounter rate and group size between transects, and prevents analysis of trend in livestock numbers. The average livestock density during the wet season was c. 16 tropical livestock units/km2, more than twice that of the dry season (Table 1). Livestock made up > 90% of ungulate biomass (range 93–99%) present in the study zone in every survey except the first, which was a dry season survey when unusually low livestock numbers were recorded.
Dorcas gazelle distribution in relation to livestock, artificial water supply, season and fire
The multiple regression model that included livestock encounter rate and season as explanatory variables had the lowest AIC value and showed a significant negative association (P < 0.05) between dorcas and livestock encounter rates, with little variation between seasons (Figs 4 & 5). Although there was no significant relationship between dorcas gazelle density and fire frequency during either 1–60 days or 61–180 days prior to the surveys at the 10 × 10 km grid scale, there was a positive relationship (P < 0.05) between dorcas gazelle density and fire frequency over the full interval of 1–180 days prior to each survey at the scale of the whole study area.

Fig. 4 Relationship between dorcas gazelle and livestock encounter rates per 10 × 10 km grid cell across all surveys in (a) the wet and (b) the dry season.

Fig. 5 Comparative distribution of (a) dorcas gazelle individuals/km and (b) combined tropical livestock units/km, derived for each 10 × 10 km grid cell across line transect and reconnaissance transect survey routes in September 2017.
Livestock distribution in relation to artificial water supply, season and fire
Livestock encounter rate was positively associated with the distribution of seasonal hafiri waterholes (P << 0.05), particularly in the wet season when hafiris were full (Fig. 6). Livestock distribution was not significantly associated with fire detections at the 10 × 10 km grid scale during the 60-days or 61–180 days prior to the survey. Across the entire study area, livestock density did not have a significant relationship (P = 0.09) with fire detection frequency.
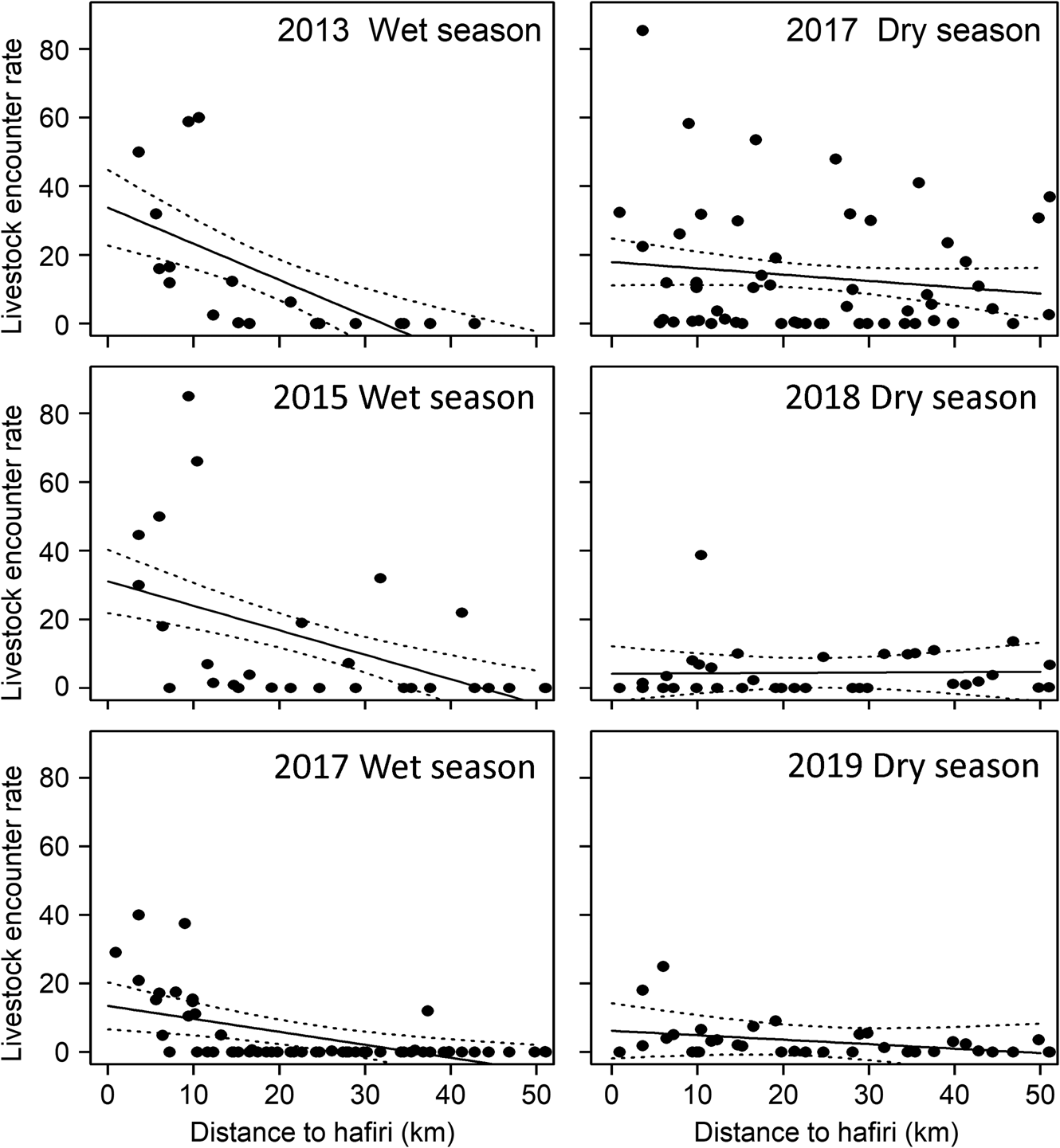
Fig. 6 Livestock encounter rate with respect to distance to hafiri, by season and year.
Dama gazelle distribution in relation to livestock and fire
The maximum opportunistic single-day count for dama gazelles across 2011–2019 was 32. Only two sightings of dama gazelles outside the study area have been reported in this period. Distribution of dama gazelles during the wet season of 2016 compared to the dry season of 2017 suggests they moved to avoid both livestock and burnt areas (Fig. 7).
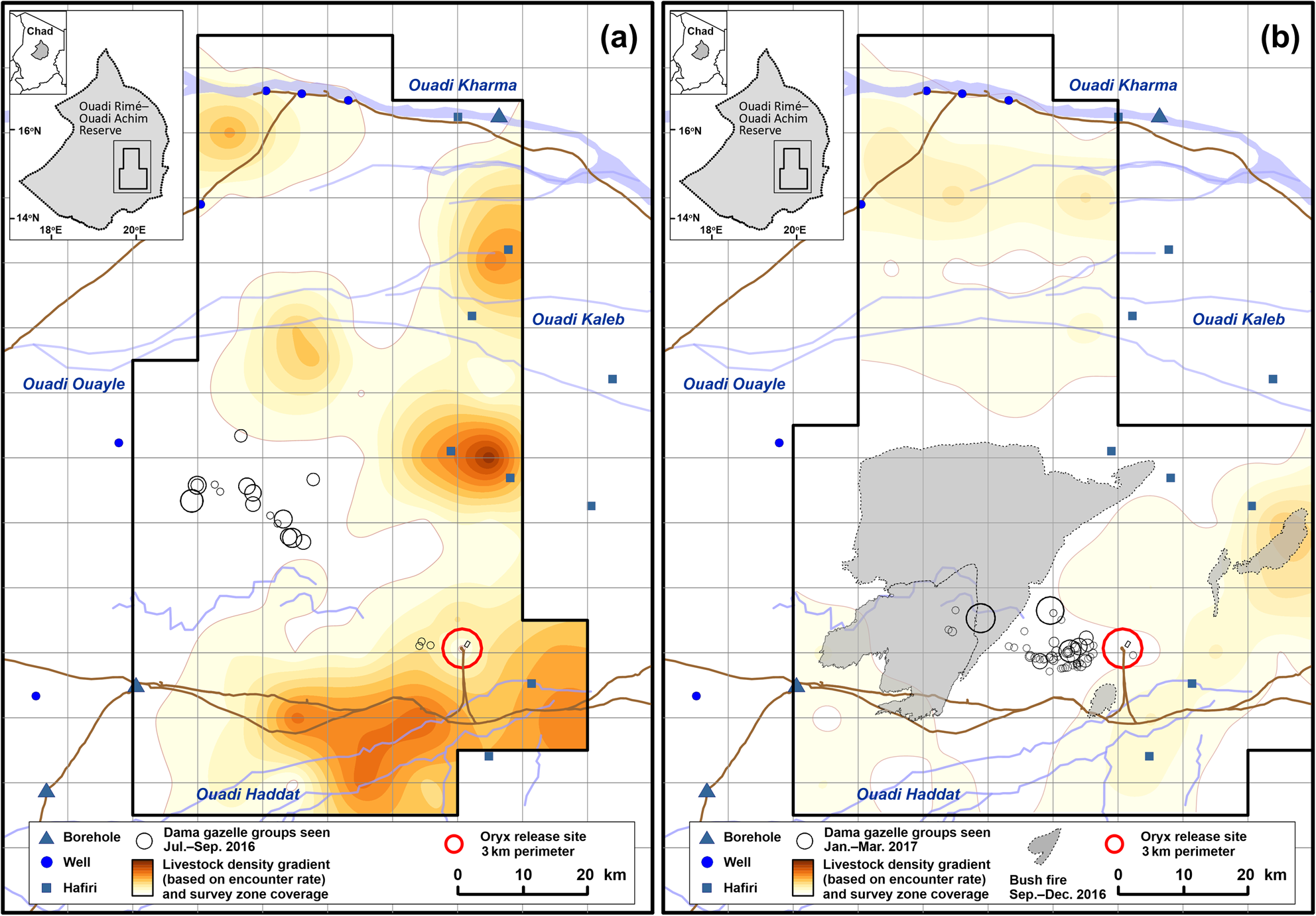
Fig. 7 Location of all dama gazelle Nanger dama sightings (circles proportional to group size; range 1–17 individuals) in relation to livestock distribution (a) during July–September 2016 (wet season) and (b) during January–March 2017 (dry season), showing reduced livestock encounter rates in the dry season, avoidance of livestock by dama gazelles in both seasons and avoidance of burnt ground by dama gazelles in the dry season.
Sample effort and precision
The mean encounter rate across the eight surveys indicated a survey effort of c. 470 km is needed to achieve a CV of 10% for dorcas gazelles. The estimated effort to achieve a CV of 20% for livestock groups is 483 km.
Discussion
National policy for the management of the Ouadi Rimé–Ouadi Achim Reserve seeks to restore natural biodiversity (Bemadjim et al., Reference Bemadjim, Newby, Desbiez, Lees and Miller2012) and improve conditions and living standards for livestock pastoralists (Bailey, Reference Bailey2020; World Bank Group, 2020). This first systematic study of wild and domestic ungulate numbers and relative distribution in the central Sahelian zone of Chad provides an evidence base to assist decision-making on land-use management to help meet these objectives (see also Newby, Reference Newby2014).
Our findings suggest that the line transect survey design and effort has achieved reasonable precision in estimated dorcas gazelle numbers (CV ≤ 20%), but only the increased effort applied in our final survey (575 km) can be expected to achieve this for livestock numbers. This is because of the high variability in livestock herd sizes and species composition, depending on season and management, and varying transhumance destinations, depending on grazing conditions.
A key result of our study is that the central area of the Reserve still supports the dorcas gazelle at high densities and almost certainly holds the largest protected population globally (see also Lafontaine et al., Reference Lafontaine, Beudels-Jamar, Devillers and Wacher2006), with no indication of persistent decline. Numbers in the study area were increasing during 2013–2019. It is not known whether this represents an increase in overall population size or increasing concentration because of environmental factors, and/or increased protection from activities related to the reintroduction of the scimitar-horned oryx. We did not examine the status of the dorcas gazelle in the wider Reserve, but it is known that significant numbers are present. The species ranges widely across the whole Reserve and can survive for long periods independently of water (Yom-Tov et al., Reference Yom-Tov, Mendelssohn and Groves1995; Scholte & Hashim, Reference Scholte, Hashim, Kingdon and Hoffman2013a). For example, in September 2017, the population in the line transect survey zone was estimated to be 5,700 (95% CI 3,100–10,700), an apparent reduction of nearly 10,000 individuals relative to the estimate for the same zone 7 months earlier (Table 1). However, the mean encounter rate in the adjacent reconnaissance survey block to the north in September 2017 was four times higher than in the line transect zone, indicating a high likelihood that many individuals had moved 30–50 km north (Fig. 5). Large assemblies of dorcas gazelles were at that time attracted to extensive green pastures of Boerhavia sp., I. colutea and other green vegetation in the reconnaissance survey zone, while avoiding high concentrations of livestock towards the east. Although dorcas gazelle distribution was independent of artificial water sources, the species uses natural water sources that form temporarily after rainfall. In a localized study zone of the Negev desert, dorcas gazelles were associated with permanent water points (Henley et al., Reference Henley, Ward and Schmidt2007). But free-living dorcas gazelles in Africa frequently experience prolonged absence of water, and routinely so in the 9-month dry season in Chad. The absence of association between dorcas gazelles and the array of artificial water points infers that habitat quality in the Reserve is still sufficient to allow dorcas gazelles to meet their water requirements across the seasonal cycle without having to rely on artificial water sources.
Extensive coverage during the survey period confirmed that dama gazelles are present in low numbers in a relatively restricted area. Nevertheless, previous research indicates this is the largest of the four remaining wild populations (Scholte & Hashim, Reference Scholte, Hashim, Kingdon and Hoffman2013b; Mungall, Reference Mungall2018; IUCN/SSC, 2019). Data collected over two seasons suggests dama gazelles avoid livestock in a similar way to dorcas gazelles, but may be even more sensitive to disturbance, commonly exhibiting much longer flight distances (often > 500 m; SCF, unpubl. data, 2020). Evidence suggests that anthropogenic influences such as livestock presence, fire and occasional hunting by motorbike affect the Critically Endangered dama gazelle more heavily than the dorcas gazelle, indicating that land-use planning and law enforcement will play a critical role in determining the viability of this small but globally important population.
Unexpectedly, our findings on the local impact of fires were inconclusive. The frequency and extent of grass fires, removing grazing biomass at the start of the long dry season, has become a major management concern for both conservation and pastoralism interests. We did not detect a significant effect of fire frequency at the scale of the 10 × 10 km grid cells on wild or domestic ungulates. However, a positive association between dorcas gazelles and fire detections at the scale of the entire study area over the previous 6 months requires further investigation to determine if this is consistent over time and whether it relates to effects of fire on habitat and food plant condition, or the relative reduction of livestock. Ground surveys of actual area burnt and food plant condition over time in burnt areas may provide more accurate data to assess fire impact.
Our observations of livestock confirm that the biomass of livestock far exceeds that of wild ungulates within the Reserve in all seasons, with transhumant management creating strong seasonal variation. Negative spatial relationships between wild and domestic ungulates have been recorded in other situations (Foufopolos et al., Reference Foufopoulos, Altizer, Dobson and Vandermeer2002; Acebes et al., Reference Acebes, Traba and Malo2012; Gordon, Reference Gordon2018). We could not determine the causal mechanism for spatial separation between dorcas gazelles and livestock in this study, but disturbance and predation are likely factors. In the Reserve, predation probably affects ungulates in two ways, with jackals Canis lupaster and striped hyaenas Hyaena hyaena having an impact on livestock, and human activities and domestic dogs (associated with many nomad camps) affecting wild ungulates. Competition for resources, especially grazing and undisturbed space, is likely to be important, and disease dynamics may also play a role. High livestock densities support increased populations of biting arthropods that can act as disease vectors, and livestock concentrations at artificial water points facilitate disease transmission. Transhumance also contributes to the spread of diseases and their vectors, and limits opportunities for veterinary management (Apolloni et al., Reference Apolloni, Corniaux, Coste, Lancelot, Touré, Kardjadj, Diallo and Lancelot2019).
Evidence that high livestock densities around water or other concentration points (piospheres) are causing significant environmental harm is not always clear (Turner, Reference Turner2000). In the Reserve, during the dry season the presence of a well can be perceived at distances of up to 5–10 km by progressive thinning of ground cover, increasing density of dry livestock dung, tracks and signs of trampling, and increased dust in the air. Contrary to this, previous research measuring primary production around well sites in the Ferlo of Senegal based on satellite data reported no loss of vegetation biomass around wells, which may be associated with persistence of unpalatable vegetation (Hanan et al., Reference Hanan, Prevost, Diouf and Diallo1991; Rasmussen et al., Reference Rasmussen, Brandt, Tong, Hiernaux, Diouf and Assouma2018). Negative effects such as soil impaction and reduced water infiltration have been detected at higher livestock grazing densities (Hiernaux et al., Reference Hiernaux, Bielders, Valentin, Bationo and Fernández-Rivera1999). Site-specific features such as soil type, vegetation community and particularly the effective livestock stocking rates and management patterns around focal points determine the impact of livestock grazing on the natural habitat. In the Ouadi Rimé–Ouadi Achim Reserve, our findings show that high stocking rates, associated with artificial water points, lead to spatial exclusion of wild ungulates.
This has implications for conservation planning. Assuming that despite the low precision, our estimates of livestock numbers are nevertheless broadly representative, the livestock numbers in the Reserve are typical for the region. The mean observed stocking rate across all surveys was 0.12 tropical livestock units/ha, with a maximum of 0.26 units/ha in some wet seasons (Table 1). This is comparable to rates of 0.15–0.20 units/ha reported from other (unprotected) areas of the Sahel (De Leeuw & Tothill, Reference De Leeuw and Tothill1990; Miehe, Reference Miehe1997; Hein & Weikard, Reference Hein and Weikard2008). In the Reserve, the pastoralist economy has access to c. 95% of all ungulate biomass and associated primary productivity. Distribution of dorcas gazelle populations (and likely other wildlife) is limited by this anthropogenic disturbance and pressure on available resources. An agro-economic case study in the Sahelian Ferlo of Senegal estimated an optimal livestock stocking rate (to maximize profit margins) of 0.09 units/ha; slightly below the estimates for environmentally sustainable and actual observed stocking rates (Hein & Weikard, Reference Hein and Weikard2008; Weikard & Hein, Reference Weikard and Hein2011). The observed mean stocking rate in the Reserve (Table 1) exceeds this economically optimal stocking rate and is at or above the estimated ecologically sustainable maximum. Therefore, managing stocking rates not to exceed current levels, and ideally slightly reduce them, could be beneficial to the pastoralist economy as well as wildlife conservation.
Spacing and management of water points are critical to managing stocking rates. The waterpoints that most affect livestock distribution in the study area are hafiris (Fig. 1). It is likely that outside the study area, boreholes and wells have a similar influence in other parts of the Reserve, where these water sources are also critical for livestock.
Water resource development is a key issue in the Sahel, often associated with disruption of traditional well ownership and creating social tensions (Cotula, Reference Cotula2006). In view of the observed livestock stocking rates in the Reserve, we believe that an effective conservation zone could be established by ensuring no further development of water resources or use of mobile bladder reservoirs takes place within a designated 8,000 km2 core area (c. 10% of the Reserve), focused on the survey blocks in this study. The size of this core area is scaled to match ecological conditions for livestock and wildlife in a historically arid and drought-prone region, and considers the impact of climate change (Fremantle et al., Reference Fremantle, Wacher, Newby and Pettorelli2013). Management within the conservation zone should aim to avoid further increase in livestock density and control fire, both of which would benefit pastoralist communities and support biodiversity conservation. Improved livestock management is also needed in the remaining 90% of the Reserve. The support and active involvement of local communities are vital for this stratified approach to succeed. To reduce potential opposition to what may be perceived as the creation of a large zone from which local people are excluded, the conservation plan should specify that livestock grazing may continue in the core zone, but no further water resource development or settlement should take place. The objectives of this management regime are to support biodiversity conservation and habitat sustainability, manage fire risk, improve veterinary care and promote the role of the conservation core zone as a grazing reserve supporting the much larger surrounding area.
To track the effect of future conservation and livestock management strategies, ongoing ground surveys with monitored law enforcement and modern aerial survey techniques (Lamprey et al., Reference Lamprey, Pope, Ngene, Norton-Griffiths, Frederick, Okita-Ouma and Douglas-Hamilton2020) will be needed. Satellite tracking of dorcas gazelles could confirm the movement patterns and requirements of this species implied by our study. This would be particularly valuable if coordinated with formal study of livestock management practice and movements. Ongoing productivity and veterinary health assessments would confirm progress in the livestock sector.
Our finding that the Ouadi Rimé–Ouadi Achim Reserve holds globally important populations of the dorcas and dama gazelles, in addition to the ongoing reintroduction of oryx (Duthie, Reference Duthie2018; Mertes et al., Reference Mertes, Stabach, Songer, Wacher, Newby, Chuven, Al Dhaheri, Leimgruber and Monfort2019) addax and ostrich populations (Sahara Conservation Fund, 2019, 2020), highlight its unique value in supporting Sahelian biodiversity. This study provides evidence for management steps needed to achieve national conservation and development goals in a complex social environment and ecosystem. Our recommendations aim to ensure that at least 10% of the Reserve continues as a protected area of IUCN category IV (IUCN, 1994) or higher and is managed as such, with management strategy and actions distinct from a sustainable development area (Locke & Deardon, Reference Locke and Dearden2005).
Acknowledgements
We thank the Government of Chad, its Ministry of Environment and Fisheries and the Direction de la Faune et des Aires Protégées for close collaboration and support throughout our fieldwork; the Private Department of HH Sheikh Mohammed bin Zayed for funding the Pan-Sahara Wildlife Survey that supported the initial fieldwork; the Environment Agency Abu Dhabi for support and leadership in establishing the oryx reintroduction project; the Société de Voyages Sahariens for logistical support during the early surveys; the rangers and staff of the oryx project basecamp for their support; and the reviewers for their critiques.
Author contributions
Survey design: TW, JN; survey observers: JN, KA, HA, SZB, FNB; data collection: TW, JN, KA, HA, MHH, SZB, FNB; data analysis: RA, TW; writing: TW, RA, JN.
Conflicts of interest
None.
Ethical standards
This research abided by the Oryx guidelines on ethical standards. All work was conducted with the permission and direct participation of the government authorities of Chad. Survey teams included local participants, ensuring effective communication with local people during informal encounters. No animals were handled during this study.









 Open access
Open access