


Introduction
Although subsistence hunting is still a critical survival strategy for many forest dwellers throughout Amazonia, for many wildlife species this practice is unsustainable under current social and economic conditions in most Amazonian settings (Redford & Robinson, Reference Redford and Robinson1987; Alvard, Reference Alvard1993; Alvard et al., Reference Alvard, Robinson, Redford and Kaplan1997; Bodmer et al., Reference Bodmer, Eisenberg and Redford1997; Mena et al., Reference Mena, Stallings, Regalado, Cueva, Robinson and Bennett2000; Peres, Reference Peres2000a; Souza-Mazurek et al., Reference Souza-Mazurek, Pedrinho, Feliciano, Hilário, Gerôncio and Marcelo2000; Zapata-Ríos, Reference Zapata-Ríos2001). Sustainable hunting implies that harvest rates of wildlife populations are at a level that meets the consumption needs of local people without reducing wildlife populations to a level at which they are in danger of local extirpation (Bennett & Robinson, Reference Bennett, Robinson, Robinson and Bennett2000). In this context, evaluation and monitoring of hunting levels is needed to avoid local extinction of important game species and to ensure the long-term cultural survival of indigenous Amazonians.
The first step towards achieving sustainability of subsistence hunting is to identify and characterize the species most frequently hunted and current extraction rates. To evaluate the balance between production and harvest this characterization needs to be combined with information on carrying capacity of the area and species’ biology. A major limitation, however, is the scarcity of the appropriate biological data and the difficulty of collecting this information in the field. This problem has been approached by the development of simple algorithms that do not require detailed biological information on any given species, thus providing coarse estimates of sustainability (Robinson & Redford, Reference Robinson, Redford, Robinson and Redford1991, Reference Robinson and Redford1994; Slade et al., Reference Slade, Gomulkievicz and Alexander1998; Robinson, Reference Robinson, Robinson and Bennett2000; Stephens et al., Reference Stephens, Frey-Roos, Arnold and Sutherland2002; Bodmer, Reference Bodmer and Polanco-Ochoa2003; Rowcliffe et al., Reference Rowcliffe, Cowlishaw and Long2003; Bodmer & Robinson, Reference Bodmer, Robinson, Silvius, Bodmer and Fragoso2004). These models, although useful, are not free of flaws and limitations, and they allow the detection of overharvest but not of sustainable hunting (Milner-Gulland & Akçakaya, Reference Milner-Gulland and Akçakaya2001).
In the southern Ecuadorian Amazon hunting is an integral part of the subsistence and culture of the Shuar group. Previously known as Jívaros, the Shuar people have lived for several centuries in south-eastern Ecuador on the lower slopes of the Andes and the Amazonian lowlands (Stirling, Reference Stirling1938; Steel, Reference Steel1999). Until recently the small population size and traditional hunting methods of this indigenous group had little impact on wildlife populations (Harner, Reference Harner1972; Descola, Reference Descola1994, Reference Descola1996). However, like many other areas in the Amazon, the Shuar territory is undergoing rapid socio-economic changes that increase the pressure on wildlife and habitats. Although there have been several anthropological studies of this indigenous group (Karsten, Reference Karsten1935; Stirling, Reference Stirling1938; Ghinassi, Reference Ghinassi1939; Harner, Reference Harner1972; Descola, Reference Descola1994; Taylor & Landázuri, Reference Taylor and Landázuri1994; Steel, Reference Steel1999), little is known about their patterns of wildlife use. Here we document Shuar hunting patterns, and assess their impact on large mammals using three sustainability algorithms: the production model (Robinson & Redford, Reference Robinson, Redford, Robinson and Redford1991), the stock-recruitment model and the harvest model (Bodmer, Reference Bodmer and Polanco-Ochoa2003; Bodmer & Robinson, Reference Bodmer, Robinson, Silvius, Bodmer and Fragoso2004).
Study area
The study site, an area of 600 km2 known as Miasal, is on the western margin of the Amazon basin at altitudes of 250–1,200 m, at the foot of the steep Kutukú mountain range in south-eastern Ecuador (Morona-Santiago province). The region, located on the easternmost border of one of the world's biodiversity hotspots (the Tropical Andes), has been recognized for its biodiversity (Mittermeier et al., Reference Mittermeier, Myers, Thomsen, da Fonseca and Olivieri1998; Myers et al., Reference Myers, Mittermeier, Mittermeier, da Fonseca and Kent2000). Miasal is defined by the watershed of the Mangosiza river, a white-water river that originates in the highlands of Kutukú (c. 2,500 m) draining the area to the south into the Morona river (Fig. 1). The predominant vegetation types are upland moist tropical forest and pre-montane tropical wet forest (Sierra, Reference Sierra1999). Climate is wet and warm, with an annual precipitation of 4,500 mm and an average monthly temperature of 24°C (Winckell et al., Reference Winckell, Zebrowski and Sourdat1997).
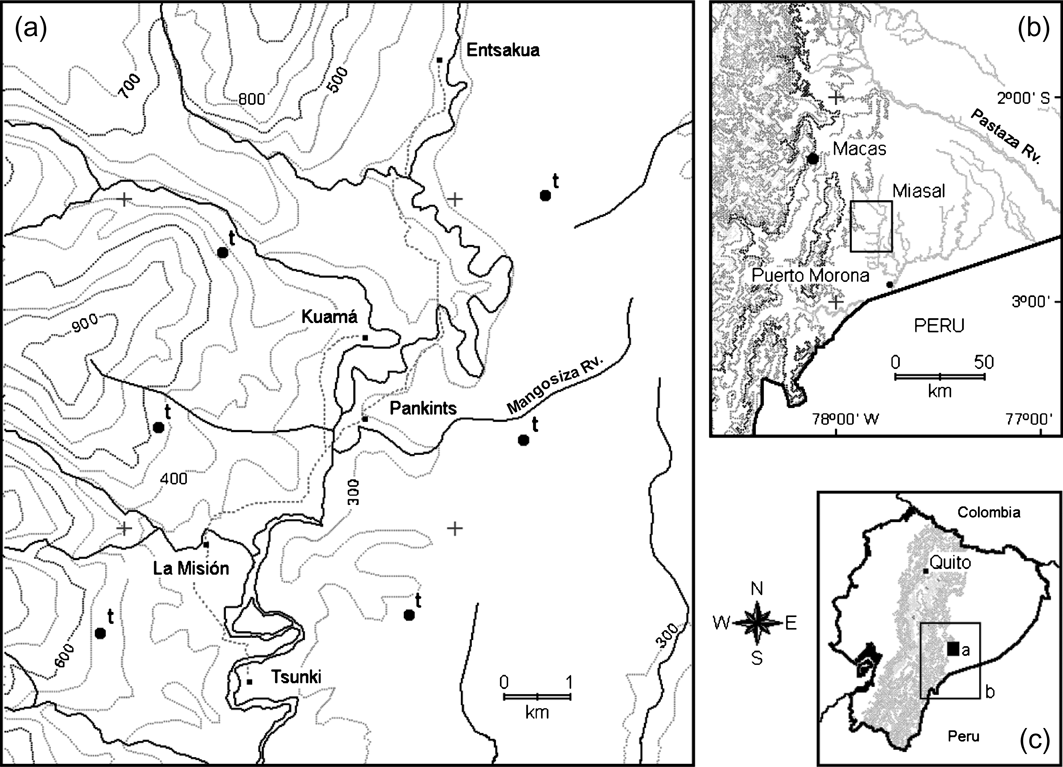
Fig. 1 (a) The study area, Miasal, on the eastern slopes of the Kutukú mountain range, in south-eastern Ecuador, showing the location of the four Shuar communities, La Misión, and the six line transects (t), (b) the regional context (Morona-Santiago province), and (c) the location of the province in Ecuador.
The Shuar are historically famous for being fierce warriors and for shrinking the heads of their enemies (Up de Graff, Reference Up de Graff1923; Steel, Reference Steel1999). With a population of c. 80,000 people this ethnic group constitutes Amazonian Ecuador's second largest indigenous group (Moya, Reference Moya1998). In Miasal the population is c. 1,000 people (c. 1.7 km-2) living in four communities or centros (the smallest administrative unit of the Shuar Federation). The four centros (Entsakua, Kuamá, Pankints and Tsunki) are located along the Mangosiza river, and were established c. 30 years ago when four Shuar families moved to the area looking for new hunting grounds. The centros are close to each other and function as one community, with La Misión as the main central settlement. La Misión is a Salesian missionary post where a church, high school, small first-aid health centre and several sport fields have been built. People from the four centros gather every weekend at La Misión to socialize and sometimes trade wild meat and agricultural produce.
Miasal is isolated from urban population centres. Depending on water level, it can be reached via the Mangosiza river in 6 h to 1.5 days from Puerto Morona by motorized canoe. From Macas, the capital city of Morona-Santiago province, it can be reached after a 45-minute flight in a small plane but the costs are high (c. USD 80 per person). Shuar also use footpaths, one parallel to the river that take 2 days to reach Puerto Morona, and another, to the west, that takes 5 days to reach Macas, crossing the Kutukú mountain range. Although, some ecotourism activities occur in the area, the economy is largely based on subsistence agriculture (plantain and manioc), and a high percentage of protein intake still comes from wild meat, mainly large mammals.
Methods
This study was carried out from September 2001 to January 2003. During the first 2 months a series of introductory meetings were organized in the four communities to obtain permission to work in the territory of Miasal and to engage the local hunters to participate and support the project. Hunting patterns and impact of hunting activities were documented during 12 complete months using interviews, direct observations, self-monitoring records (Townsend, Reference Townsend, Fang, Montenegro and Bodmer1999; Noss et al., Reference Noss, Cuéllar and Cuéllar2003, Reference Noss, Cuéllar and Cuéllar2004; Zapata-Ríos & Jorgenson, Reference Zapata-Ríos, Jorgenson, Campos-Rozo and Ulloa2003), participatory mapping (Chapin & Threlkeld, Reference Chapin and Threlkeld2001; Sheil et al., Reference Sheil, Puri, Basuki, van Heist, Rukmiyati and Sardjono2002) and mammal surveys (Sutherland, Reference Sutherland1996; Wilson et al., Reference Wilson, Cole, Nichols, Rudran and Foster1996; Rabinowitz, Reference Rabinowitz1997). The last 3 months of the study were dedicated to return the research results to the communities, design wildlife management strategies, and sign agreements with the communities for the implementation of wildlife management plans.
An inventory of species harvested in the area was carried out using several techniques. In addition to accompanying hunters for direct observation of hunting techniques and prey harvested, a series of bi-weekly structured interviews were conducted with hunters. Interviews, in Spanish, included questions on species hunted and local names, and characteristics of the prey (sex, age, reproductive status, weight). Respondents were also asked to identify on a map the approximate location where the hunt took place, the vegetation type, the time of the hunting event and weapons used. To involve hunters in this research and to reach the more distant households, we developed a hunter self-monitoring form (written in both Spanish and Shuar). The self-monitoring forms, a simplified and graphical version of the structured interviews, asked for the same information. The self-monitoring form also included a sketch map for hunters to note the approximate location of the hunt. These forms were presented to the hunters in a series of workshops to ensure the questions were clear and to obtain the input of the participants. Every hunter that agreed to participate received a spring scale for measuring the weight of prey. In addition to completing the forms, hunters were asked to provide skulls, feathers or bones of hunted animals to assess the reliability of the information provided by the self-monitoring and interviews. The data from the interviews and self-monitoring forms were used to estimate actual harvest rates. Data from these sources also allowed us to characterize hunting patterns, estimate the hunting catchment area, and describe hunter preferences.
To estimate the density of prey populations in the catchment area we established six transects of 2,000 m, located on both sides of the Mangosiza river on the hunting grounds of the four Shuar communities (Fig. 1). Each transect was surveyed weekly during 6.00–08.00 and 18.00–20.00. We walked slowly (c. 1 km h-1), looking for mammals and mammal tracks and signs. Censuses were not conducted on rainy days (Peres, Reference Peres1999). The species, number of individuals, sighting or radial distance (observer to animal distance) and angle, and distance along the transect were recorded for mammals observed. Population densities were calculated using Hayne's estimator, a robust estimator that relies on radial distance observations (Hayne, Reference Hayne1949; Hayes & Buckland, Reference Hayes and Buckland1983; Krebs, Reference Krebs1999). In the case of social species, individual density was estimated by multiplying group density by the average group size. Density estimates and information gathered in the interviews and with the self-monitoring forms were used for estimating the sustainability of hunting with three models (Table 1): the production model (Robinson & Redford, Reference Robinson, Redford, Robinson and Redford1991), the stock-recruitment model and the harvest model (Bodmer, Reference Bodmer and Polanco-Ochoa2003; Bodmer & Robinson, Reference Bodmer, Robinson, Silvius, Bodmer and Fragoso2004).
Table 1 The three sustainability models used to estimate the impact of hunting, and the sources of data.

Hunters’ preferences were analysed using simple linear regression and χ2 goodness-of-fit tests. A simple linear regression model was used to test whether prey availability predicts species' harvest levels (Rao, Reference Rao1998), evaluating the null hypothesis that hunters are selecting prey species at random (higher numbers will be extracted of more abundant species). The χ2 goodness-of-fit-test evaluated the null hypothesis of no difference between proportions of use by the hunters and prey availability. In addition, 90% adjusted Wald confidence intervals (ŵ) for the expected proportions of use (p exp) were calculated for each individual species to determine whether a species is preferred (Agresti & Coull, Reference Agresti and Coull1998; Agresti, Reference Agresti2007). Where the observed proportion (p obs) of use did not lie within the interval (p exp ± ŵ), differences between expected and observed use of species were identified as significantly different.
The three algorithms require population parameters (Table 1) and current harvest rates. The production and harvest models assess sustainability by comparing actual extraction rates with estimated production thresholds. In the production model (Robinson & Redford, Reference Robinson, Redford, Robinson and Redford1991; Robinson, Reference Robinson, Robinson and Bennett2000) we compared the harvest rates (E) for each species obtained from the interviews and self-monitoring forms to the theoretical maximum sustainable harvest (H) estimated using the density data from the line-transects and the intrinsic rate of natural increase (Robinson & Redford, Reference Robinson and Redford1986). In the harvest model we also compared the harvest rates (E) obtained from the interviews and self-monitoring forms to a production estimate based on estimated fecundity rates (litter size and gestations per year) and population densities (Bodmer, Reference Bodmer and Polanco-Ochoa2003; Bodmer & Robinson, Reference Bodmer, Robinson, Silvius, Bodmer and Fragoso2004).
The stock-recruitment model (Bodmer, Reference Bodmer and Polanco-Ochoa2003; Bodmer & Robinson, Reference Bodmer, Robinson, Silvius, Bodmer and Fragoso2004) does not use an estimate of production as a standard for comparison. It is based on a density-dependent population model that uses maximum sustainable yield (MSY) and an estimate of carrying capacity to assess the status of wildlife populations in hunted areas. The model (Bodmer & Robinson, Reference Bodmer, Robinson, Silvius, Bodmer and Fragoso2004) assesses hunting sustainability for different population sizes depending on the distance between population size and carrying capacity (K). Population size (as a percentage of K) was obtained from the density estimates, and K was assumed to be the population size in unhunted areas. Because obtaining data from unhunted areas was logistically infeasible, these estimates were obtained from the literature (Mena et al., Reference Mena, Regalado, Cueva, Mena, Soldi, Alarcón, Chiriboga and Suárez1997). These density estimates (from the Waorani territory in the Ecuadorian Amazon, 150 km north of Miasal) were obtained and analysed implementing the same methodology used in this study, in an area with similar ecological characteristics (Sierra, Reference Sierra1999). The combined use of the three models increases the reliability of our assessment of the sustainability of the wildlife harvest.
Results
The Shuar of Miasal are devoted hunters, using traditional and introduced techniques. Hunting tools include guns, blowguns, sticks, machetes and dogs. There is a short supply of ammunition for guns in the area, and prices are highly variable. When ammunition is scarce Shuar hunters use seeds of the palm Aiphanes schultzeana (Bennett et al., Reference Bennett, Baker and Gómez2002), known as ampakai-kamancha. Seventy-nine percent of all hunting events we recorded involved the use of non-traditional devices, particularly guns. There were no Shuar hunters in the area who remembered how to craft blowguns and prepare the poison for darts (curare). They purchase these items from Achuar traders (indigenous people from the Ecuadorian-Peruvian border) and sometimes use them. The average age of male hunters was 30 years (n = 94, range 13–54). Women also participate in hunting activities, especially hunting around the house and in the kitchen garden. Because of the community's isolation, commercial hunting does not occur, although a minimum level of trade takes place among members of the communities, providing subsistence level income used for medicines and school supplies for children.
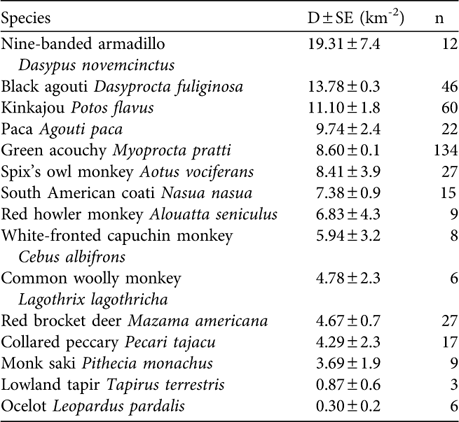
The total sampling effort along the six transects was 384 km. Density estimates were calculated for 15 species that are frequently hunted (Table 2). A total of 401 individuals of the 15 species were recorded, i.e. a sighting rate of 1.04 sightings km-1. Sightings per species ranged from three (lowland tapir Tapirus terrestris) to 134 (green acouchy Myoprocta pratti). These numbers represent a wide range in sampling effort to obtain a detection event (2.8 km for M. pratti, and 128 km for T. terrestris). The minimum recommended sample size for obtaining robust density estimates of 20 detection events per species (Peres, Reference Peres1999) was only obtained for six of the 15 species.
Table 2 Density (D) estimates for the 15 species of mammal that were frequently hunted in Miasal.
In the initial meetings 176 hunters were identified. After an initial effort to accompany Shuar hunters (38 trips with 12 hunters) this methodology was discarded as an objective means of gathering hunting data because our presence was affecting the behaviour of the hunters (e.g. showing off their skills with blowguns by aiming at small species such as hummingbirds and cockroaches). Other techniques were more successful. A total of 284 interviews were conducted with 30 hunters on a bi-weekly basis and a total of 2,397 self-monitoring forms were gathered during the 12 months. The self-monitoring forms were completed at least once by 160 hunters (91% of the hunters in the study area), and 119 hunters (68%) periodically reported their activities. A total of 1,989 items such as skulls, bones, feathers, and skins of hunted animals accompanied the self-monitoring forms. These items verified 83% of the forms. With the consent of the hunters the items that were considered of biological importance were submitted to the Ecuadorian Museum of Natural Sciences in Quito.
Although the Shuar harvest a wide range of species (52), including insects, amphibians, reptiles, birds and mammals (fish were not included in the assessment), we only report information about mammals. Data from interviews and self-monitoring forms demonstrated that medium and large mammals (> 1 kg) were the preferred prey, comprising 61% of the total biomass harvested. A total of 3,181 individuals, belonging to 21 mammal species, were hunted during the study, with collared peccary Pecari tajacu (7,177 kg) being the most important (Table 3). Other important species were red brocket deer Mazama americana, common woolly monkey Lagothrix lagotricha, T. terrestris, paca Agouti paca and nine-banded armadillo Dasypus novemcinctus (Table 3). Based on participatory mapping the hunting catchment area was c. 243 km2 (40% of the total territory of the four communities). A total of 825 hunting events (26% of 3,181) ground-truthed using a global positioning system confirmed the map sketches obtained from self-monitoring forms and generated during participatory mapping exercises (Fig. 2). Data from participatory mapping, interviews, and self-monitoring forms also suggest that large-bodied species (> 5 kg) have been extirpated (red howler monkey Alouatta seniculus, L. lagotricha and T. terrestris) or have been substantially reduced (M. americana, P. tajacu and A. paca) within a 3 km radius of the communities (Fig. 2). Hunters travel > 3 km from the communities to capture these species.
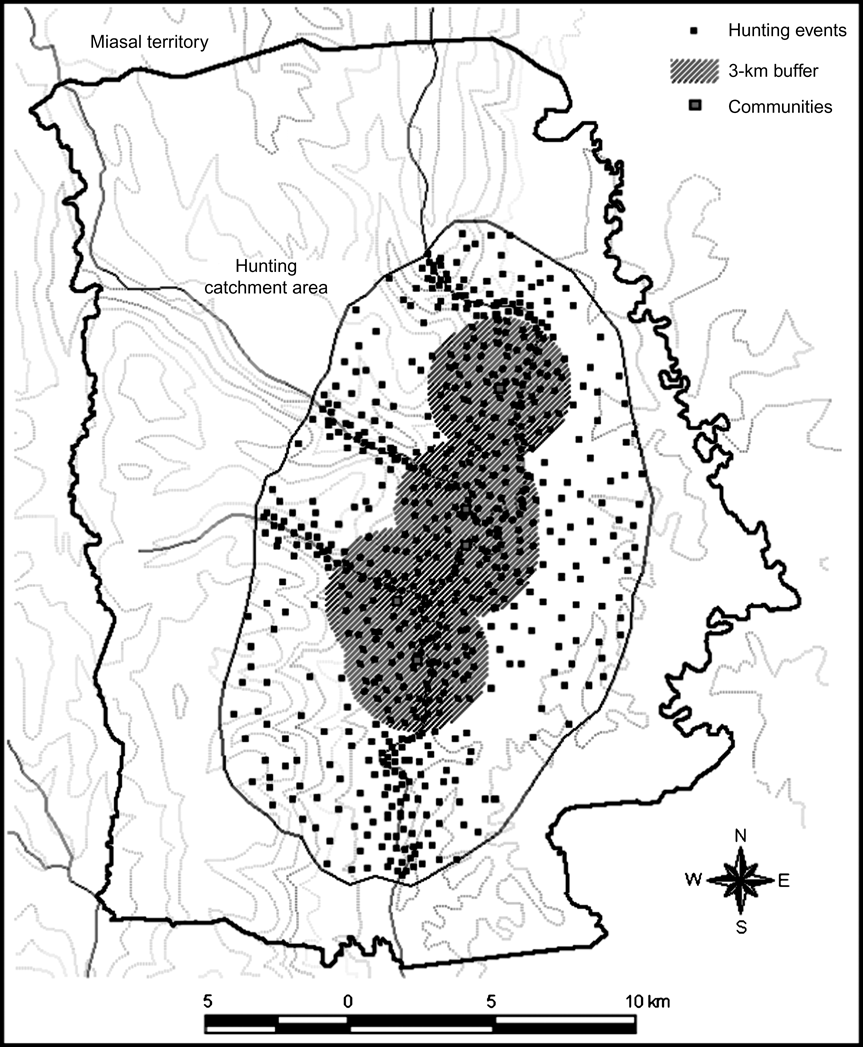
Fig. 2 Miasal (Fig. 1) showing the hunting catchment area (243 km2; 40% of the total territory of the four communities). The black symbols indicate the location of the 825 hunting events that were ground-truthed with a global positioning system. An area surrounding the communities (within a 3 km radius) where several large-bodied species have been extirpated or become rare is shown (see text for details).
Table 3 Total number of individuals (n) and biomass (mean per individual, total and per km2 per year) of 21 species of mammal extracted from the 243 km2 hunting catchment area in November 2001–October 2002.
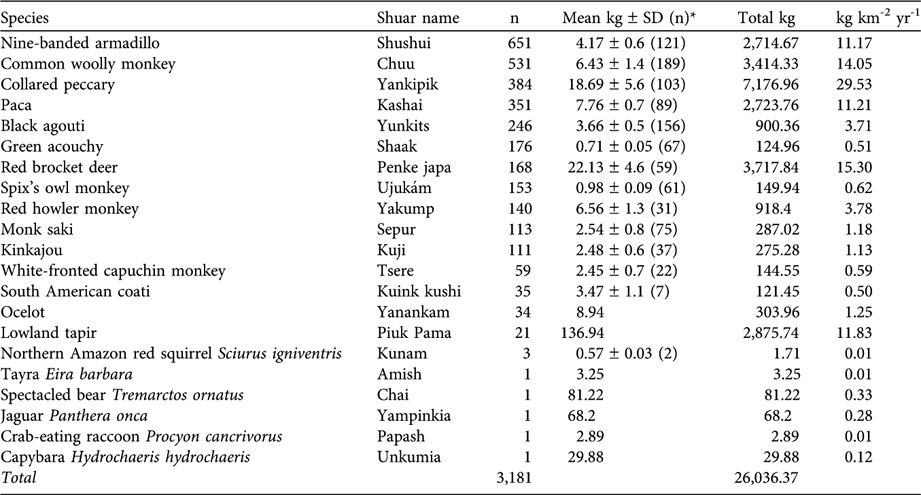
* Weights reported were obtained from hunted prey recorded in the field, except for T. ornatus, L. pardalis, P. onca and T. terrestris, data for which were obtained from Seymour (Reference Seymour1989), Padilla & Dowler (Reference Padilla and Dowler1994), Murray & Gardner (Reference Murray and Gardner1997), Eisenberg & Redford (Reference Eisenberg and Redford1999) and Tirira (Reference Tirira2007)
The goodness-of-fit test (χ 2 = 71.41, P < 0.0001) indicates that the proportions of species hunted differed significantly from the proportion of species available (Table 4). The adjusted Wald confidence intervals show that L. lagotricha and P. tajacu were hunted significantly more than expected according to availability (Table 4). The linear regression also suggests that Shuar hunters prefer to hunt large-bodied mammals such as L. lagotricha (n = 531) and P. tajacu (n = 384), which occur in low densities, than more abundant species such as A. paca (n = 351) and black agouti Dasyprocta fuliginosa (n = 241; Fig. 3).

Fig. 3 Numbers of 15 species of prey most commonly hunted versus their density (Table 2), and linear regressions for all species (solid line, r 2 = 30.68%, P = 0.03) and excluding the two outliers (Lagothrix lagotricha and Pecari tajacu; dashed line, r 2 = 68.38%, P = 0.0007). The regressions suggest that prey species are not hunted according to their availability (see text for details).
Table 4 Preferences of Shuar hunters (for the 15 species of mammal of which > 3 individuals were hunted) analysed using a χ2 goodness-of-fit test and 90% adjusted Wald confidence intervals (ŵ) for the expected proportions of use (p exp) of prey species. Where the observed proportions of use (p obs) do not lie within the interval p exp ± ŵ, differences between expected and observed use of species are considered significantly different (in bold).

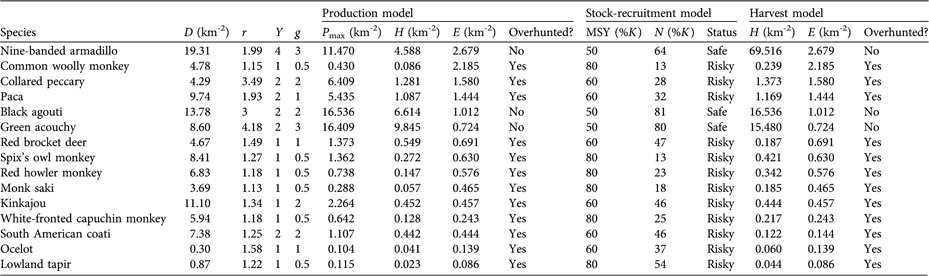
Of the 21 mammal species hunted 15 had sufficient data to be included in the sustainability assessment. The results of the three algorithms are consistent, and all suggest that only three species are not overhunted (D. novemcinctus, D. fuliginosa and M. pratti; Table 5).
Table 5 Results of the sustainability assessment, using the production model (Robinson & Redford, Reference Robinson, Redford, Robinson and Redford1991), the stock-recruitment model and the harvest model (Bodmer, Reference Bodmer and Polanco-Ochoa2003; Bodmer & Robinson, Reference Bodmer, Robinson, Silvius, Bodmer and Fragoso2004). See Table 1 for model details and parameters.
Discussion
Although line transects have been successfully used to survey mammals in several Neotropical localities (Carrillo et al., Reference Carrillo, Wong and Cuarón2000; Peres, Reference Peres, Robinson and Bennett2000b; Cullen et al., Reference Cullen, Bodmer, Valladares-Pádua, Ballou, Silvius, Bodmer and Fragoso2004) the technique is not free of limitations. The major shortcoming is that large sample sizes are required to apply the estimation models. Recommended sample sizes are 20–80 sightings (Peres, Reference Peres1999; Buckland et al., Reference Buckland, Anderson, Burnham, Laake, Borchers and Thomas2001). In many tropical forest localities, however, to obtain this number of detection events requires a sampling effort of hundreds or even thousands of km (Peres, Reference Peres1999; Carrillo et al., Reference Carrillo, Wong and Cuarón2000). Although we only obtained the minimum recommended sample size of 20 for six of the 15 species our density estimates lie within the same range as in other hunted Neotropical areas (Hill & Padwe, Reference Hill, Padwe, Robinson and Bennett2000; Mena et al., Reference Mena, Stallings, Regalado, Cueva, Robinson and Bennett2000; Peres, Reference Peres, Robinson and Bennett2000b; Cullen et al., Reference Cullen, Bodmer, Valladares-Pádua, Ballou, Silvius, Bodmer and Fragoso2004).
Shuar hunting as practised today in Miasal is not sustainable. Extraction levels of 80% of large mammals (12 species out of 15 included in the sustainability assessment) are well above sustainable harvest levels. The main reason for overharvesting of these species is the change of traditional hunting practices and their replacement with more effective hunting methods in the context of the rapid socio-economic changes that have increased demand and depleted wildlife populations (Bennett & Robinson, Reference Bennett, Robinson, Robinson and Bennett2000; Robinson & Bennett, Reference Robinson and Bennett2004). For example, human population growth in the study area (from four to > 100 families in the last 30 years) has caused an unprecedented demand that has increased hunting pressure (INEC, 2002). Human population density is high (Robinson & Bennett, Reference Robinson, Bennett, Robinson and Bennett2000) for a population that depends on wildlife for its major source of protein (c. 4.1 people km-2 in the hunting catchment area, c. 1.7 people km-2 for the total area of the communities). As expected, our results (Table 5) confirm that mammal species with higher intrinsic rates of natural increase (r), higher densities (N), and higher fecundity rates (Y and g) are more resilient to hunting pressure than those species with lower levels of the same parameters (Bodmer & Robinson, Reference Bodmer, Robinson, Silvius, Bodmer and Fragoso2004). Under these circumstances, unless urgent wildlife management strategies are implemented (e.g. hunting quotas, hunting rotations, access to alternative protein sources, and changes in consumer behaviour), some species will probably go locally extinct in the near future.
Shuar hunters show preferences for large-bodied species according to optimal foraging principles (Emlen, Reference Emlen1966; MacArthur & Pianka, Reference MacArthur and Pianka1966), valuing different species based on their body mass-to-capture cost ratio, thus maximizing hunting benefits in terms of economy, energy and time allocated (Bodmer, Reference Bodmer1995; Fitzgibbon et al., Reference Fitzgibbon, Mogaka, Fanshawe, Robinson and Bennett2000; Jerozolimski & Peres, Reference Jerozolimski and Peres2003; Hilaluddin et al., Reference Hilaluddin, Jandrotia and McGowan2004). In the study area, however, large-bodied species (> 5 kg) such as A. seniculus, L. lagotricha, T. terrestris, M. americana, P. tajacu, and A. paca have been depleted in the immediate vicinity of the four communities, and hunters travel > 3 km to find them. Shuar hunters, therefore, are reaching a threshold where they will no longer be able to be selective. Local depletion in the areas that surround the four communities (Fig. 2) may explain why the Shuar are exploiting a range of medium- and small-sized species (Fig. 3) to obtain higher returns from hunting activities, a common response to overhunting (Mena et al., Reference Mena, Stallings, Regalado, Cueva, Robinson and Bennett2000; Jerozolimski & Peres, Reference Jerozolimski and Peres2003). This phenomenon has reached its extreme in other areas of Ecuador where small rodents and marsupials (e.g. Proechimys semispinosus, Oryzomys spp. and Didelphis marsupialis) are the main source of protein for local communities (Suárez et al., Reference Suárez, Stallings and Suárez1995). Biological and ecological differences between species (population density, life span, reproductive rates) render some more susceptible to overharvesting than others (Bodmer, Reference Bodmer1995; Bodmer et al., Reference Bodmer, Eisenberg and Redford1997). Large primates (e.g. A. seniculus, L. lagotricha, Ateles belzebuth), tapirs and other large-bodied species are more vulnerable to the negative impacts of hunting than small-bodied species such as acouchies (M. pratti), agouties (D. fuliginosa) and armadillos (D.novemcinctus; Table 5).
The hunting assessment presented here represents only 12 months of a dynamic and ongoing process of wildlife extraction. Sustainability indices cannot show that a harvest is sustainable, only that it is unsustainable. Therefore, we cannot be sure that the three species (D. novemcinctus, D. fuliginosa and M. pratti) whose extraction levels suggest they are not currently overhunted will continue to be so in the long-term. The hunting grounds of the communities (243 km2) are surrounded by a larger area (c. 350 km2) that is part of the legal territory of the communities. This larger territory may still be an important wildlife source area, which could explain why overhunted species have not yet been extirpated within the catchment area after several decades of harvesting (Joshi & Gadgil, Reference Joshi and Gadgil1991; Novaro et al., Reference Novaro, Redford and Bodmer2000). Anthropogenic pressures are increasing as surrounding communities are starting to use this source area as their hunting grounds. Current patterns of wildlife exploitation can only be expected to intensify with current rates of population growth throughout Shuar territory (c. 6.5% per year; unpubl. data of the Shuar Federation, 2004). Research elsewhere in the Neotropics also reports that current patterns of wildlife use are unsustainable (Peres, Reference Peres, Robinson and Bennett2000b; Naughton-Treves et al., Reference Naughton-Treves, Mena, Treves, Alvarez and Radeloff2003; Altrichter, Reference Altrichter2005; de Thoisy et al., Reference de Thoisy, Renoux and Julliot2005; Franzen, Reference Franzen2006). This regional pattern is going to cause local and regional extinctions of preferred species and people will no longer be able to obtain a sufficient protein supply from hunting. Ultimately, the impact of this overhunting will have negative ecological and socio-economic repercussions.
Although many Shuar hunters recognize that protein sources from wildlife populations are steadily diminishing, the immediate need to conserve wildlife populations to ensure its future availability is not obvious to hunters who still enjoy what they perceive to be an inexhaustible source of meat. In this context, management of Shuar hunting practices to control harvest levels is complex. Support for this project was obtained because the Shuar were interested in developing a locally-managed ecotourism operation and were worried about the decline of wildlife species in the areas surrounding their communities, areas that are frequented by the tourists that visit the area. The results of these analyses were presented to the communities on several occasions during and at the end of the study. After a series of eight 1-day participatory meetings the four communities signed, in January 2003, an agreement for the implementation of a series of wildlife management strategies. During these meetings the communities discussed pragmatic strategies, including establishing hunting zones, prohibiting hunting by outsiders, implementing offtake quotas (by sex, age and season), and prohibiting hunting of highly vulnerable species such as T. terrestris.
Since the agreement was signed the members of the four Shuar communities have been implementing the wildlife management strategies and a wildlife monitoring programme with limited external support. Periodical patrols of their territory have lowered, but not eliminated, hunting by outsiders. The majority of the hunters are actively involved in this community-based initiative and comply with the community regulations, although some continue to overharvest wildlife species and are under strong social pressure to stop this practice. However, several internal conflicts unrelated to hunting are currently threatening the long-term implementation of the wildlife management plan. This community-owned process is still promising but the results need to be consolidated to guarantee the long-term sustainability of hunting by the Shuar of these four communities.
Acknowledgements
Funding was provided by USAID through CARE International's Program of Sustainability and Regional Union. We are grateful to Ecuador's Ministry of the Environment for granting permission to undertake this study (026-IC-FAU/DBAP/MA). The Shuar Federation granted permission for working in Shuar territory. The Shuar community of Miasal (Entsakua, Kuamá, Pankints and Tsunki) provided great support and hospitality throughout the study. It is impossible here to name all of the Shuar hunters who assisted in the field but we extend our warm thanks to all of them. We are greatly indebted to Juan Carlos Jintiach and Patricia Arcos and her family for their continuous logistical support. The effort of the Arcos family to conserve their territory is meritorious and remarkable. Servicio Aéreo Misional provided us safe and reliable transportation by small plane to Miasal. Juan Francisco Rivadeneira of the Ecuadorian Museum of Natural Sciences received and catalogued the specimens provided by the Shuar hunters. Edison Araguillin carried out most of the mammal surveys and Carlos Morales helped gather hunting data. Víctor Utreras, Karen Kainer, Brian Child and Ari Martinez provided useful comments on earlier drafts, and the insightful comments of David S. Wilkie and two anonymous reviewers greatly improved this manuscript.
Biographical sketches
Galo Zapata-Ríos’s interests include the ecology and conservation of Neotropical mammals, in particular the effects of human activities on wildlife and issues concerning development, deforestation, hunting and human-wildlife conflict. He is currently studying the ecology and distribution of carnivores in the human-dominated landscapes of the Ecuadorian Andes. Carlos Urgilés's interests include mammalogy and wildlife management. He is currently working in a community-based wildlife management programme in the Ecuadorian Amazon, implemented by the Instituto para la Conservación y Capacitación Ambiental, a local NGO. Esteban Suárez started his career working with Afroecuadorian and Chachi indigenous groups in north-western Ecuador, where he worked with local hunters characterizing their use of small mammals as food. He has also worked with campesino communities in the cloud forests of north-western Ecuador where, in addition to research activities, he collaborated with the local communities in several management and conservation education activities.








