


Introduction
How and why pottery became such a common and widespread form of material culture is a key question for archaeologists around the world. In north-eastern North America, early pottery was first used by hunter-gatherers slightly before 3000 BP, at the start of the Early Woodland Period (Taché & Hart Reference Taché and Hart2013). Based on our previous lipid analysis of the contents of over 130 Early Woodland Vinette 1 pottery vessels from north-eastern North America (Taché & Craig Reference Taché and Craig2015), we suggested that pottery initially developed in this region for use in the processing of freshwater and marine organisms at episodic social gatherings during periods of high resource abundance (Taché & Craig Reference Taché and Craig2015). Recently, Skibo et al. (Reference Skibo, Malainey and Kooiman2016) have challenged this hypothesis, reporting an absence of aquatic-derived lipids in 12 sherds from three sites in the Great Lakes region. The authors consequently argue that our proposed model for early pottery use cannot be applied to the wider north-eastern part of North America, including the Great Lakes. While this may be true, the purpose of the present article is to demonstrate that the discrepancies between these studies are probably due to the use of different analytical procedures and criteria for residue identification, rather than any real differences in how pottery was used across the region.
Why fatty acid distributions are inappropriate for categorising archaeological residues
The argument presented in Skibo et al. (Reference Skibo, Malainey and Kooiman2016) is based on the analysis of a small sample of sherds from the Schultz (n = 5), Naomikong Point (n = 4) and Grand Island (n = 3) sites in the Great Lakes region using gas chromatography (GC) and gas chromatography mass spectrometry (GCMS). The approach taken was to match relative proportions of fatty acids—one of the most common lipid classes preserved in archaeological pottery—with ‘reference ranges’ from modern samples, which had been subjected to heating in order to simulate the effects of oxidative degradation over time (Malainey et al. Reference Malainey, Przybylski and Sherriff1999). The authors conclude that the absence of long-chain fatty acids and the ratio of C18:1 to C18:0 fatty acids are typical of plants and terrestrial animals. None of the samples appear to be derived from fish, which, according to their criteria, have 15–27.5 per cent of the total fatty acid content comprised of C18:1.
We argue that the approach used by Skibo et al. (Reference Skibo, Malainey and Kooiman2016) for categorising residues is inadequate because it relies on the assumption that laboratory degradation (30 days at 75°C) accurately simulates degradation in different depositional environments over millennial timescales. The alteration of lipids in pottery following use and deposition is a complex process, as demonstrated by degradation experiments and analysis of archaeological potsherds from different archaeological contexts. While lipids absorbed during use can be traced back to the original contents of a vessel, variation in the modes of use (e.g. time, temperature, oxidative conditions), the nature of the ceramics (e.g. porosity, clay matrix) and a range of reactions in the burial environment alter structure and distributions of lipids, complicating direct comparison with experimental analogues (Evershed et al. Reference Evershed, Stott, Raven, Dudd, Charters and Leyden1995, Reference Evershed, Copley, Dickson and Hansel2008; Regert et al. Reference Regert, Bland, Dudd, Bergen and Evershed1998; Evershed Reference Evershed2008).
Generally, (poly)unsaturated fatty acids are more liable to oxidative degradation than saturated compounds and are therefore preferentially lost (Regert et al. Reference Regert, Bland, Dudd, Bergen and Evershed1998). Shorter-chain fatty acids are more soluble that longer-chain compounds and will be preferentially lost to percolating groundwater. Triacylglycerol distributions are also altered through hydrolysis, with the preferential loss of lower molecular weight components (Dudd et al. Reference Dudd, Regert and Evershed1998). Fatty acids derived from the soil bacteria can also make a contribution to the overall distribution in the potsherd (Dudd et al. Reference Dudd, Regert and Evershed1998). Without knowing the extent of these complex and varied post-depositional degradation processes, it is wholly unrealistic to expect that organic residues can be reliably classified on the basis of their fatty acid distributions. The method of extraction may also affect the distribution of fatty acid liberated from the sample (Regert et al. Reference Regert, Bland, Dudd, Bergen and Evershed1998; Correa-Ascencio & Evershed Reference Correa-Ascencio and Evershed2014; Deviese et al. Reference Deviese, Van Ham-Meert, Hare, Lundy, Hommel, Bazaliiskii and Orton2018).
To circumvent the issues, Taché and Craig (Reference Taché and Craig2015) adopted two strategies that have been widely applied over the last 20 years, and which have arguably transformed the field of archaeological residue analysis. The first is to identify chemical biomarkers—molecules or suites of molecules that can be traced to their source with a high degree of confidence, taking into account degradation. For example, ω-(o-alkylphenyl)alkanoic acids (APAAs) formed during the protracted heating of unsaturated fatty acids are stable in the burial environment, and their distribution can be used to identify broad, precursor foodstuffs, such as aquatic and plant oils (Hansel et al. Reference Hansel, Copley, Madureira and Evershed2004; Evershed et al. Reference Evershed, Copley, Dickson and Hansel2008; Cramp & Evershed Reference Cramp, Evershed and Turekian2014). As the isomerisation reaction needed to form these unusual compounds only occurs at high temperatures and is assisted by the presence of a clay matrix, they can be reliably traced to foodstuffs heated during vessel use.
Secondly, carbon isotopic values of individual fatty acids can be measured to identify the origins of the lipids; these values are maintained, despite extensive exposure to the burial environment (Evershed et al. Reference Evershed, Mottram, Dudd, Charters, Stott, Lawrence, Gibson, Conner, Blinkhorn and Reeves1997; Evershed Reference Evershed2008). This approach has been widely used to identify marine foods in pottery, which generally have fatty acids enriched in the heavier carbon isotope (13C) compared with terrestrial and freshwater products (e.g. Copley et al. Reference Copley, Hansel, Sadr and Evershed2004; Craig et al. Reference Craig, Steele, Fischer, Hartz, Andersen, Donohoe, Glykou, Saul, Jones, Koch and Heron2011; Cramp et al. Reference Cramp, Evershed, Lavento, Halinen, Mannermaa, Oinonen, Kettunen, Perola, Onkamo and Heyd2014; Lucquin et al. Reference Lucquin, Gibbs, Uchiyama, Saul, Ajimoto, Eley, Radini, Heron, Shoda, Nishida and Lundy2016a; Oras et al. Reference Oras, Lõugas, Tõrv, Kriiska and Craig2017), and to distinguish between ruminant and non-ruminant fats (Craig et al. Reference Craig, Allen, Thompson, Stevens, Steele and Heron2012). Skibo et al. (Reference Skibo, Malainey and Kooiman2016) did not report the presence or absence of aquatic biomarkers, or employ compound-specific isotope analysis. As the former are best identified as methyl esters on a polar chromatography column and with the mass spectrometer in single ion monitoring (SIM) mode for increased sensitivity (see Cramp & Evershed Reference Cramp, Evershed and Turekian2014), it is unlikely that these would be observable in the experimental conditions used by Skibo et al. (Reference Skibo, Malainey and Kooiman2016).
Applying fatty acid distribution criteria to vessels with aquatic biomarkers
The discrepancy between Skibo et al.’s (Reference Skibo, Malainey and Kooiman2016) and Taché and Craig's (Reference Taché and Craig2015) approaches is exemplified if we apply Malainey et al.’s (Reference Malainey, Przybylski and Sherriff1999) criteria to the Vinette 1 vessels that produced the full set of aquatic biomarkers (see the online supplementary material (OSM)). When using Malainey et al.’s (Reference Malainey, Przybylski and Sherriff1999) criteria, of the 47 ‘aquatic’ samples, 68 per cent (n = 32) would have been interpreted as being from ‘large herbivores’, 21 per cent (n =10) from ‘plants (except corn)’ and six per cent (n = 3) from ‘plant with large herbivores’; two samples do not fit any of the criteria. Importantly, the highest relative concentration of C18:1 detected in the 110 interpretable Vinette 1 vessels is 10.27 per cent, significantly less than the 15–27.5 per cent interval ascribed to aquatic resources (Malainey et al. Reference Malainey, Przybylski and Sherriff1999), and without any relationship to the position of the residue on the vessel (contrary to the suggestion in Malainey Reference Malainey, Barnard and Eerkens2007). Setting such a high relative concentration of C18:1 as the criterion for identifying aquatic lipids in archaeological vessels inevitably leads to erroneous conclusions. These include Skibo et al.’s (Reference Skibo, Malainey and Kooiman2016: 1235) interpretation that Early Woodland ceramics were seldom or never used to prepare aquatic resources, or Malainey et al.’s (Reference Malainey, Przybylski and Sherriff2001) proposal that ‘fish avoidance’ characterised hunter-gatherer populations of the Northern Plains of North America.
The higher susceptibility of C18:1 to post-depositional oxidation (Regert et al. Reference Regert, Bland, Dudd, Bergen and Evershed1998) probably explains its diminished abundance in residues derived from aquatic sources. Even exceptionally well-preserved archaeological residues contain much lower proportions of mono- and polyunsaturated fatty acids than predicted based on extrapolations from modern reference samples artificially ‘aged’ by heating (Malainey Reference Malainey, Barnard and Eerkens2007). A 350-year-old Late Thule rim sherd with excellent lipid preservation (7362μg g−1) and clear association with a maritime economy on the Alaska Peninsula (Dumond Reference Dumond2003), for example, produced a relative C18:1 concentration of 15.41 per cent—barely above the minimum threshold for fish outlined by Malainey et al. (Reference Malainey, Przybylski and Sherriff1999). Yet, the enrichment of 13C in the fatty acids, along with the presence of three isoprenoid acids and the full range of APAAs C18−22, leave no doubt that the vessel was used exclusively for the processing of marine resources (see the OSM: NAK-8).
Differences in the overall distributions of 12 fatty acids (C12:0, C14:0, C15:0, C16:1, C16:0, C17:0, C18:1, C18:0, C20:1, C20:0, C24:1, C24:0) in samples with and without aquatic biomarkers from north-eastern North America (n = 110) and Alaska (NAK-8, above) were investigated using principal component analysis (PCA) based on a variance-covariance matrix and the PAST3 software package (version 3.16 for Windows) (Figure 1; also see the OSM). Analysis of the 111 × 12 data matrix shows that the first two principal components (PCs) account for 73.9 per cent and 18.4 per cent of the total variance in the dataset, respectively. Despite slight differences between groups in relative concentrations of C18:0, C12:0 and C14:0 fatty acids, there is little overall correspondence between fatty acid distributions and the presence or absence of aquatic biomarkers. Thus, the claim that fatty acid distributions are inappropriate for categorising archaeological residues is further supported.
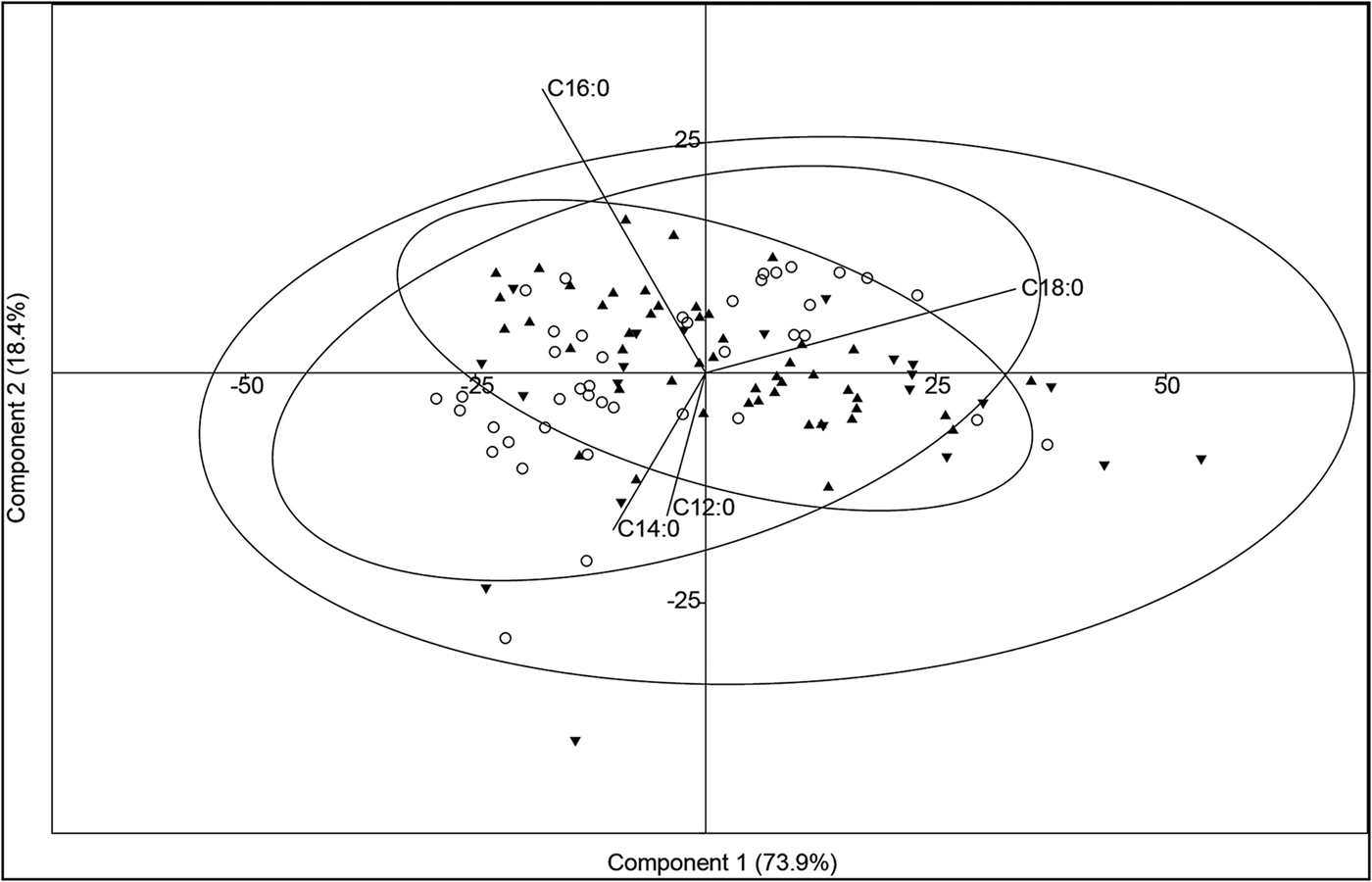
Figure 1. Principal component analysis (PCA) scatter plot of the first two principal components (PCs) for residues with the full set of aquatic biomarkers (filled triangles), the partial set of aquatic biomarkers (filled inverted triangles) and without aquatic biomarker (empty circles). The three ellipses represent group membership, assuming the 95 per cent confidence limit (2σ) as the cut-off distance.
Applying the isotope and lipid biomarker approach to pottery from the Great Lakes
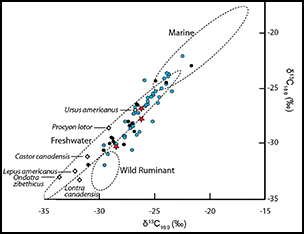
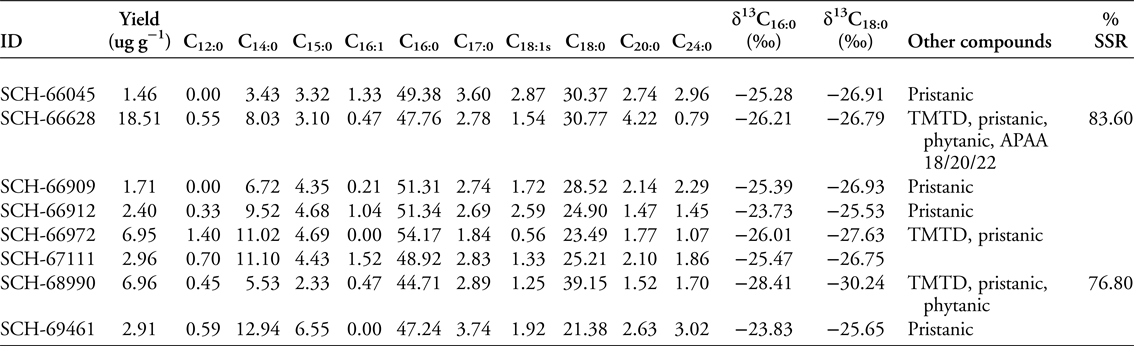
To test whether the Great Lakes pottery was used for processing aquatic resources, we used the protocols described in Cramp and Evershed (Reference Cramp, Evershed and Turekian2014) to analyse lipids in eight ceramic samples from the Schultz site. These were different potsherds to those reported by Skibo et al. (Reference Skibo, Malainey and Kooiman2016). Lipids were generally poorly preserved, with only three samples producing interpretable quantities (i.e. >5μg g−1 ceramic). Significantly, we found that the sample with the highest lipid concentration yielded APAAs C18−22 and three isoprenoid fatty acids (TMTD, phytanic acid and pristanic acid; Figure 2 & Table 1)—satisfying the criteria for aquatic lipid identification (Evershed et al. Reference Evershed, Copley, Dickson and Hansel2008; Cramp & Evershed Reference Cramp, Evershed and Turekian2014). Dihydroxy fatty acids, which are also used to identify aquatic resources (Cramp & Evershed Reference Cramp, Evershed and Turekian2014), could not be detected, probably due to the overall poor lipid preservation. In contrast, diastereomers of phytanic acid in two of the Schultz samples showed a clear dominance of the SSR isomer (>75 per cent; Table 1). This is typical of an aquatic, rather than a terrestrial, source (Lucquin et al. Reference Lucquin, Colonese, Farrell and Craig2016b). Finally, carbon isotope analysis of fatty acids from the three samples produced values consistent with freshwater fish (Table 1 & Figure 3; Cramp & Evershed Reference Cramp, Evershed and Turekian2014; Taché & Craig Reference Taché and Craig2015). In the present study, isotope values alone cannot be used to identify aquatic resources, given that they are also consistent with a range of other non-ruminant terrestrial products. These three samples, however, are clearly outside the range of isotope values obtained from ruminant animals, despite levels of C18:0 exceeding 27.5 per cent in two samples. According to Malainey et al’s (Reference Malainey, Przybylski and Sherriff1999) criteria, these levels should result from the preparation of ‘large herbivores’, which in the case of Early Woodland north-eastern North America can only be ruminants such as deer, moose, woodland caribou, or wapiti/North American elk. Taché and Craig (Reference Taché and Craig2015) also reported low proportions of ruminant fats in their sample of Vinette 1 pottery, with only two of the 52 vessels submitted to GC-combustion-isotope ratio MS (GC-C-IRMS) analysis having fatty acid δ13C values comparable with wild ruminant fat.
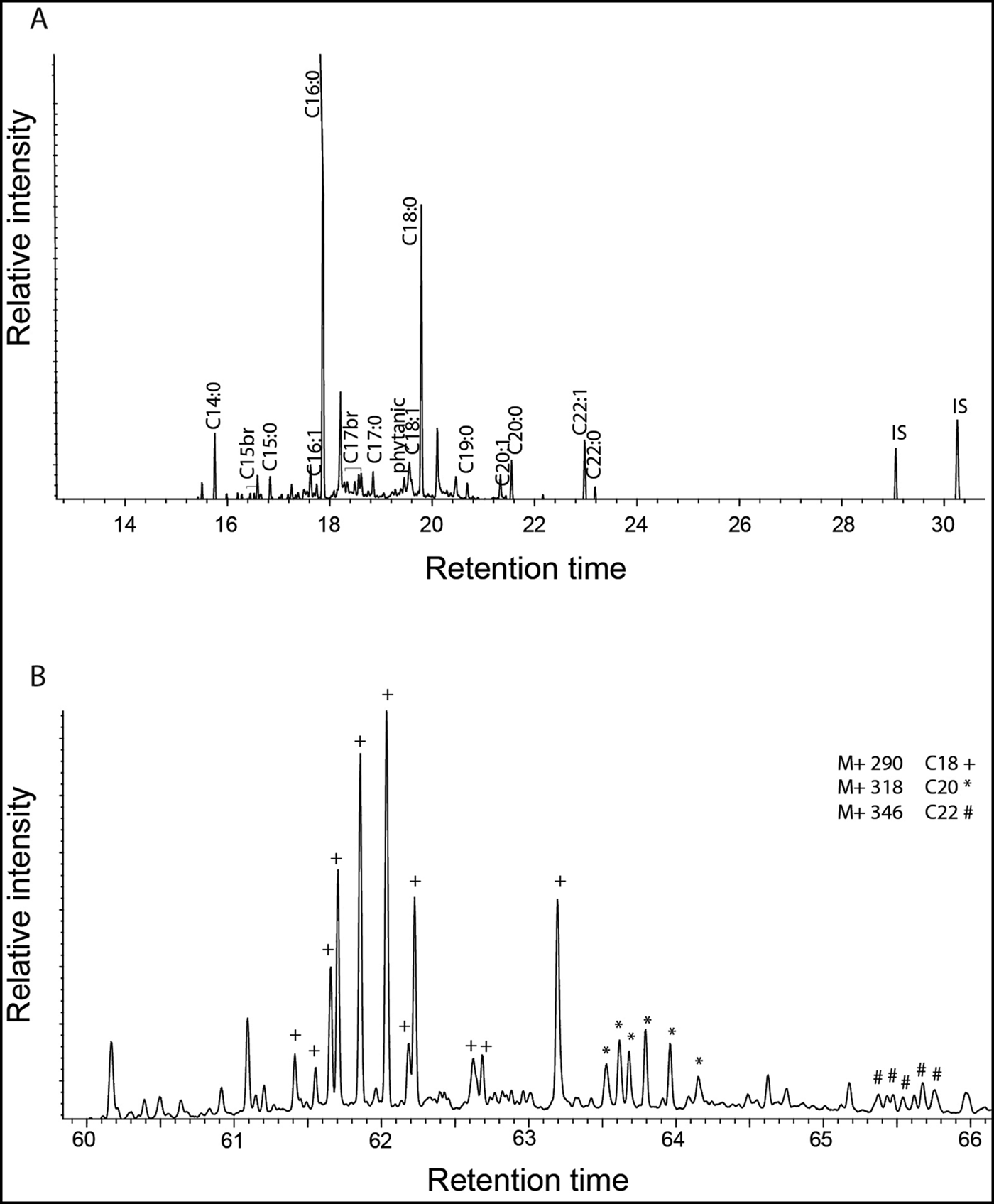
Figure 2. A) Partial chromatogram of lipid extract 66628 from the Schultz site, containing degraded aquatic oil. Cn:x are fatty acids with carbon length n and number of unsaturations x; br are branched-chain acids; IS are the internal standards (n-tetratriacontane and n-hexatriacontane); B) partial m/z 105 ion chromatogram of lipid extract 66628 ran on DB23 column (60m, 250μm, 0.25μm) with the mass spectrometer in SIM mode shows ω-(o-alkylphenyl)alkanoic acids with 18 (+), 20 (*) and 22 (#) carbon atoms.
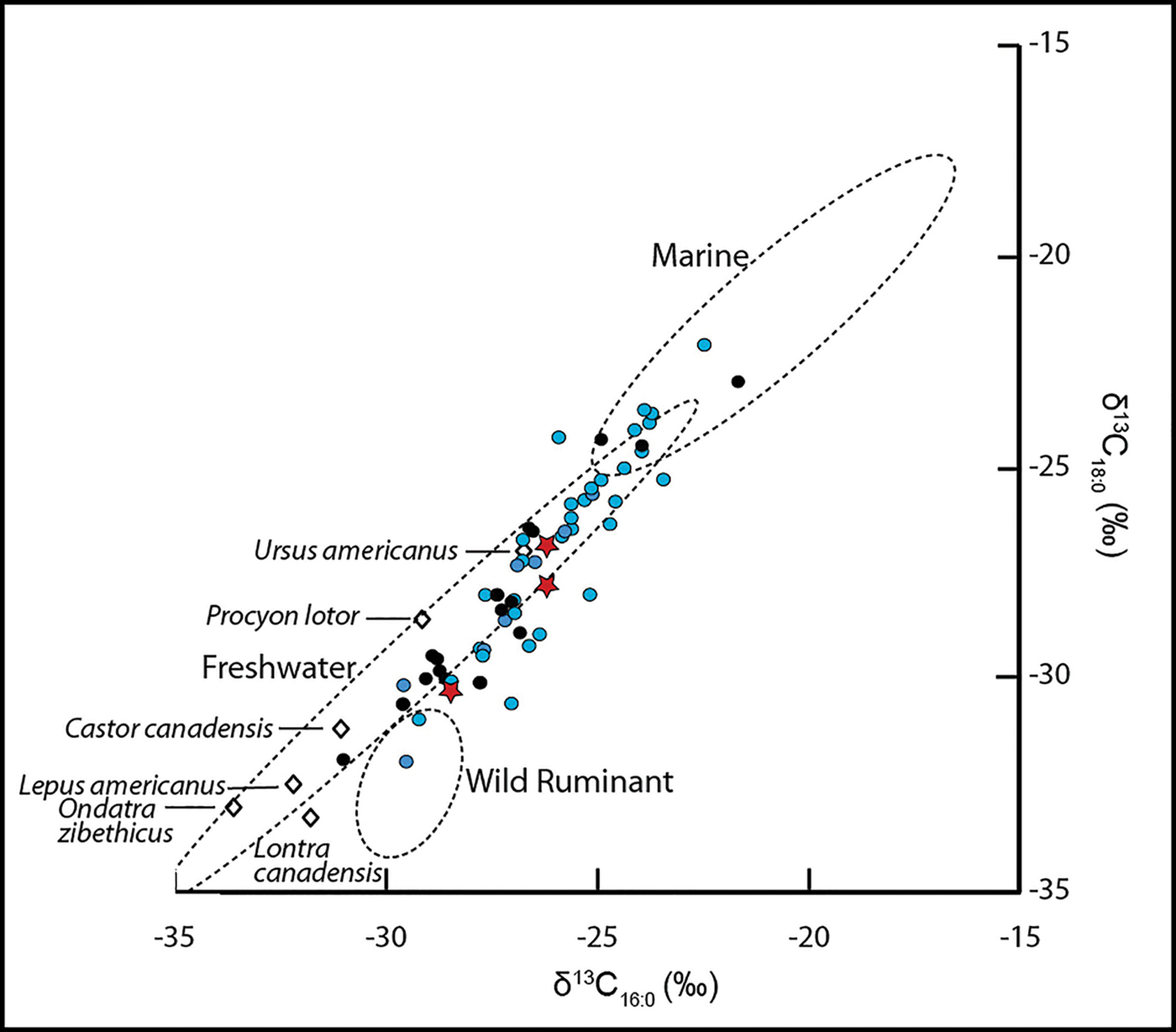
Figure 3. Plot of the δ13C values of C16:0 and C18:0 fatty acids extracted from three Schultz site potsherds (red star symbols). These are compared with Vinette 1 pottery with (blue circles) and without (black circles) aquatic biomarkers, and with reference fats from wild ruminant (n = 19), freshwater (n = 24) and marine (n = 22) organisms plotted as confidence ellipses (1σ; Stastica v.7). Additional North American authentic fat samples from wild herbivorous, omnivorous and piscivorous animals (n = 8) are indicated separately (open diamonds).
Table 1. Lipid analysis of ceramic samples from the Schultz site. Cx:y = fatty acids with carbon length x and number of unsaturations y; phy = phytanic acid; APAA Cx = ω-(o-alkylphenyl)alkanoic acids with carbon length x; %SSR = the contribution of the SRR isomer in total phytanic acid.
Conclusion
We argue on both theoretical and empirical grounds that the analytical approach used by Skibo et al. (Reference Skibo, Malainey and Kooiman2016) is unsuitable for the identification of aquatic-derived lipids due to the variable preservation of fatty acids in archaeological residues, which prevents comparison with modern reference data. Instead, we contend that the criteria outlined by Evershed et al. (Reference Evershed, Copley, Dickson and Hansel2008), Cramp and Evershed (Reference Cramp, Evershed and Turekian2014) and Lucquin et al. (Reference Lucquin, Colonese, Farrell and Craig2016b) provide the only robust approach for identification of aquatic lipids in archaeological pottery—supplemented, where possible, by carbon isotope measurements of individual fatty acids. By applying these criteria to a small sample from the Schultz site, we show that, contrary to the findings of Skibo et al. (Reference Skibo, Malainey and Kooiman2016), aquatic resources were indeed processed in early pottery (2500 BP) in the Great Lakes region.
While we accept that the production of ceramic vessels by early pottery-using hunter-gatherers of north-eastern North America and the Great Lakes region for processing fish may not have been ubiquitous—the sample size is currently too small to confirm this—the two activities do seem to be closely related, when the empirical evidence is examined with due scientific rigour. On a wider point, we recommend that any conclusions regarding pottery use based on distributions of fatty acids are carefully scrutinised and, where possible, verified using other, more robust criteria. Experiments to simulate degradation, of course, can be very useful, but such lines of evidence are at best illustrative and should be used to assist and enhance, rather than reduce and limit interpretations.
Acknowledgements
We thank the Museum of Natural and Cultural History in Eugene, Oregon, for providing access to the NAK8 sample, and the Laboratory of Anthropology at the University of Michigan for providing access to the Schultz site samples. We would also like to thank Mary E. Malainey for her comments and suggestions on an earlier draft. This work was supported by a Marie Curie Incoming International Fellowship (grant#273392), the Social Sciences and Humanities Council of Canada (grant#756-2011-0321), the Arts and Humanities Research Council (AH/L00691X/1) and EU Horizon 2020 (Marie Curie Actions Grant Agreement 676154).
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.15184/aqy.2019.128




