


Introduction
Biological control can be a highly effective and cost-efficient pest control method (Van den Bosch and Telford Reference Van den Bosch and Telford1964; DeBach and Rosen Reference DeBach and Rosen1991). However, as living organisms, biological control agents are strongly influenced by farm management decisions. In an integrated pest management (IPM) programme, chemical and cultural controls are often used concurrently with biocontrol. Broad-spectrum insecticides may kill the natural enemies along with the targeted pests (DeBach Reference DeBach1947; Settle et al. Reference Settle, Ariawan, Astuti, Cahyana, Hakim, Hindayana and Lestari1996; Fagan et al. Reference Fagan, Hakim, Ariawan and Yuliyantiningsih1998; Bommarco et al. Reference Bommarco, Miranda, Bylund and Björkman2011), while cultural controls may either augment or diminish biocontrol potential (DeBach and Rosen Reference DeBach and Rosen1991; Marc et al. Reference Marc, Canard and Ysnel1999). Because integrated pest management has been a cornerstone of agricultural production since the concept was developed in the mid-1900s (Stern et al. Reference Stern, Smith, van den Bosch and Hagen1959; Smith et al. Reference Smith, Apple and Bottrell1976), a critical focus of recent agricultural research has been on predicting which control strategies will be most compatible with each other, and on reconciling the use of chemical and biological controls. Specifically, conservation biological control involves deliberate maintenance of natural enemy niche requisites, while reducing disruptive factors that inhibit natural enemy activity (DeBach Reference DeBach1964; DeBach and Rosen Reference DeBach and Rosen1991; Barbosa Reference Barbosa1998).
In commercial cranberries (Vaccinium macrocarpon Aiton (Ericaceae)) in Wisconsin (United States of America), most growers use pest thresholds and pesticide alternatives in their integrated pest management programmes (Colquhoun and Johnson Reference Colquhoun and Johnson2010). However, Wisconsin cranberry growers have not intentionally implemented biological control to any significant degree (Singleton Reference Singleton2010), despite observations that many native natural enemies exist in the cranberry system, including egg parasitoids, larval parasitoids, and generalist predators (Mahr Reference Mahr1999; Singleton Reference Singleton2010). Of these, the most efficient parasitoids have been shown to affect up to 97% of their pest hosts in a field population (Mahr Reference Mahr1999). It appears, therefore, that conservation biological control could provide significant pest suppression in cranberry systems (Marucci and Moulter Reference Marucci and Moulter1992; Li et al. Reference Li, Henderson and Myers1994; Simser Reference Simser1994; Mahr Reference Mahr1999; Singleton Reference Singleton2010).
About half of Wisconsin cranberry growers flood their beds for one to two days in late spring, as a cultural pest control technique, as well as to remove significant quantities of detritus and detritivores (Steffan et al. Reference Steffan, Singleton, Sokja and Zalapa2012a). This flood takes advantage of the fact that cranberries are primarily a dry, irrigated crop, but the plant can withstand several days of submergence during cooler weather. The flood effectively replaces a corresponding insecticide application, and has been shown to control for the most economically important pests in cranberry (Steffan et al. Reference Steffan, Singleton, Sokja, Zalapa, Dittl and Harbut2012b). However, it remains unclear through what mechanisms the flood controls these pests. It is unlikely that most caterpillar suppression is a result of the flood itself, given past work showing low caterpillar mortality in flood waters (Teixeira and Averill Reference Teixeira and Averill2006; Steffan et al. Reference Steffan, Singleton, Sokja and Zalapa2012a).
We propose two other ways in which the flood may manifest an effect on pest caterpillar population densities. First, by replacing an insecticide application, the flood may conserve biological control agents (Marucci and Moulter Reference Marucci and Moulter1992; Singleton Reference Singleton2010). Second, the flood may suppress pest populations through an indirect effect on generalist predator feeding habits (Fig. 1). The removal of large amounts of detritus and many detritivores, which represent an alternative prey source, may force hungry generalist predators to prey-switch. If this is the case, predators will consume proportionally more pest herbivores (which come from a “green”, or plant-based, food web) than they would in a situation with abundant detritivorous prey (which come from a “brown”, or detritus-based, food web, Birkhofer et al. Reference Birkhofer, Wise and Scheu2008; Kuusk and Ekbom Reference Kuusk and Ekbom2010). If either of these situations are found to occur following flooding in cranberry production, then the flood would in fact represent conservation biological control, wherein changes to management practices preserve the native natural enemy populations.
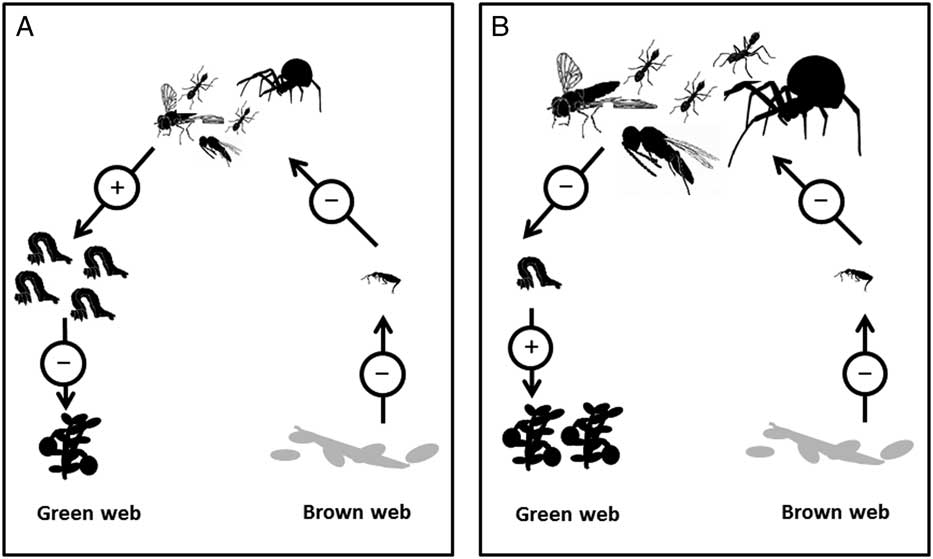
Fig. 1 Two hypothetical consequences of the removal of detritus in a cranberry ecosystem, depicting energy moving up the “green” (plant-based) and the “brown” (detritus-based) food webs. A, Detritus removal leads to starvation of and decreased numbers of detritivores, and correspondingly also decreased numbers of predators. The few remaining predators consume fewer herbivores, leading to increased pest populations and having a detrimental effect on the crop. B, Detritus removal leads to starvation of and decreased numbers of detritivores. However, the generalist predators prey-switch, maintaining population numbers, but consuming proportionally more herbivores. This leads to a decreased pest population, and has a beneficial effect on the crop.
This study provides foundational information on the impacts of spring flooding on cranberry arthropods, illuminating mechanisms through which flooding suppresses pest caterpillars by showing the effects of the flood, compared with an insecticide application, on natural enemy abundance. In addition, comparing pest control methods is valuable from a practical standpoint to the cranberry growers. The moth species investigated, Sparganothis sulfureana Clemens (Lepidoptera: Tortricidae), Rhopobota naevana Hübner (Lepidoptera: Tortricidae), and Acrobasis vaccinii Riley (Lepidoptera: Pyralidae), are among the most damaging cranberry pests. Acrobasis vaccinii is a direct pest, feeding on up to six berries during their single larval generation each year. Rhopobota naevana has two generations per year, and can decrease photosynthetic capacity of the plant as well as damaging buds and flowers. Sparganothis sulfureana has two generations, with the first feeding on cranberry foliage in the spring, and the second feeding inside the fruit. Although the cost of both implementing the flood and applying an insecticide are highly variable, flooding is in general the cheaper pest control strategy.
To address the mechanisms through which flooding may affect pest caterpillar abundances, we looked at the abundances of herbivores, detritivores, parasitoids, and predators following the flood or the spray. We hypothesise that biological control and the cultural control strategy of flooding are compatible, because the flood conserves natural enemies, and that together they can provide pest suppression equivalent to or greater than a conventional insecticide spray.
Methods
A total of 121.4-ha cranberry beds (Stevens variety, ~25 years old) were identified from within a single commercial marsh in central Wisconsin, United States of America. Each of the 76 beds is ~340×50 m, and separated from the next bed by either a dirt road or a dirt road and an ~25 m wide water reservoir. Study beds were separated from each other by at least 120 m. In all, 34 of the beds (including six study beds) received an ~30 hours spring flood on 28–29 May 2014, and did not receive any spring insecticide application (designated as “Flood”). Flood waters came from nearby water reservoirs. The other 42 beds (including six study beds) were not flooded, and were treated with the insecticide Hatchet (active ingredient chlorpyrifos) (Dow AgroSciences; Indianapolis, Indiana, United States of America) on 30 May 2014 (designated as “Spray”). All insecticide applications were administered at the registered label rates, and were applied using a tractor-mounted boom spraying method, in accordance with conventional cranberry cultivation practices. Blocks to flood or spray were constrained by the marsh layout; however, the location of the study beds was randomised from within available beds.
Sampling
Beginning in May 2014, arthropod communities were sampled weekly for seven consecutive weeks. Depending on weather conditions and marsh spray schedules, actual sampling intervals ranged from six days to 10 days between sampling events. The initial sampling period took place seven days before flooding or spraying. Before the seventh week, all beds received an application of the insecticide Altacor (active ingredient chlorantraniliprole) (DuPont, Wilmington, Delaware, United States of America). Each sampling event used three separate methods: pitfall traps, sweep netting, and pheromone-baited traps. Voucher specimens are being archived within the University of Wisconsin Insect Research Collection (Madison, Wisconsin, United States of America).
Pitfall traps were non-baited and measured the activity-density of arthropods in the understorey. Two pitfall traps were placed in each bed, located on opposite ends approximately one-quarter of the long length into the bed and ~2 m from the side of the bed. Each pitfall trap was constructed from a 500-mL plastic cup (Dart Container, Mason, Michigan, United States of America) buried to soil-surface level in the cranberry bed and covered with a plastic roof to prevent rainfall from entering the trap. Each cup contained ~150 mL of 100% propylene glycol (ChemPoint, Bellevue, Washington, United States of America). Traps remained in the field for seven days at a time, contents were collected weekly, and propylene glycol was added as needed. Arthropods were strained and stored in 95% ethyl alcohol (Pharmco-Aaper, Brookfield, Connecticut, United States of America).
Sweep-netting measures the density of arthropods in the cranberry canopy. Each sweep sampling event consisted of six sets of 20 sweeps with a 38-cm-diameter sweep net (BioQuip, Rancho Dominguez, California, United States of America). Sweep sampling took place across the width of the bed, with two sets of 20 sweeps each to transverse the 50-m bed. This was repeated three times moving down the length of the bed. All sweep netting occurred between 10:00 AM and 04:00 PM, with no precipitation. Arthropods were stored in airtight bags on ice in a cooler, and then transferred to vials of 95% ethyl alcohol. All arthropods in the pitfall and sweep-netting samples were identified to order. In order to separate each into functional trophic guilds, some orders were identified to family or genus.
Pheromone-baited traps consisted of P2 diamond-shaped sticky-traps (Great Lakes IPM, Vestaburg, Michigan, United States of America), placed ~0.5 m above the cranberry canopy, and baited with lures for adult male moths of S. sulfureana, A. vaccinii, and R. naevana (ISCA Technology, Riverside, California, United States of America). Each bed contained six total traps in two groups of three placed at opposite ends of the beds near the pitfall traps, such that one baited for each moth species was placed at each end. The number of moths in each pheromone-baited trap was counted and recorded in the field. Traps were replaced as necessary and lures were replaced every three weeks.
Statistical analyses
All statistical analyses were conducted using repeated measures analysis of variance (ANOVA), except for parasitoids due to their low abundance. Repeated measures ANOVA analyses were used to compare treatment effects across all seven weeks of sampling. Parasitoid wasp pitfall trap data were analysed using a Mann–Whitney Rank Sum test, in which samples from week 2 through week 6 were summed for analysis. Week 1 was excluded from parasitoid analysis because it took place before flood or spray events, and week 7 was excluded because all beds were sprayed the previous week. Insufficient parasitoid wasps were found in sweep samples (0–15 per bed), so these data were not analysed.
For pheromone-baited traps, counts for each of the three pest moth species were analysed individually, with the two trap counts for each species in each bed summed. For pitfall and sweep-netting samples, analyses were first conducted at the taxonomic level of order. These were then assigned to feeding guilds (herbivore, detritivore, predator, or parasitoid) based on extension publications and grower expertise. The most abundant order from each guild was considered an indicator of overall guild population dynamics; these were the orders that were used for analyses.
The trap catch for each order from the two pitfalls traps in each bed was summed. In week 4, extensive rain flooded most of the pitfall traps, so pitfall data from week 4 was excluded. For sweep netting, the total number of insects in each order per bed was added together. Arthropod abundances that were not normally distributed were log (x+1) transformed (sweep net samples of Collembola and spiders).
All statistical analyses were conducted in SigmaPlot 12.5 (Systat Software 2013).
Results
During the seven weeks of sampling, 7122 arthropods were collected in sweep netting, and 46 000 arthropods and mollusks were collected in pitfall traps. Throughout their flight, 2288 moths were caught in pheromone-baited traps. Across all sampling methods, the majority of arthropods collected belonged to the Acari (25 000) and Collembola (22 000). Other frequently captured taxa included Lepidoptera (2300), Diptera (2000), and Araneae (1000). Hymenoptera were much less frequently captured (225).
Herbivores
There was no significant difference in pheromone trap catch between flooded and sprayed beds for any of the key pest species (S. sulfureana: F(1,10)=1.655, P=0.227; A. vaccinii: F(1,10)=0.763, P=0.403; R. naevana: F(1,10)=0.335, P=0.575; Fig. 2). Adult moth trap catches for all species in both flood and spray beds followed a typical flight pattern, with a mid-summer pulse in trap counts, and all populations peaking in early July. However, for S. sulfureana there were fewer moths caught in flooded beds than in sprayed beds during peak flight week, as indicated by a significant treatment×week interaction (F(5,50)=4.566, P=0.002; pairwise contrast week 7: t=4.133, P<0.001).
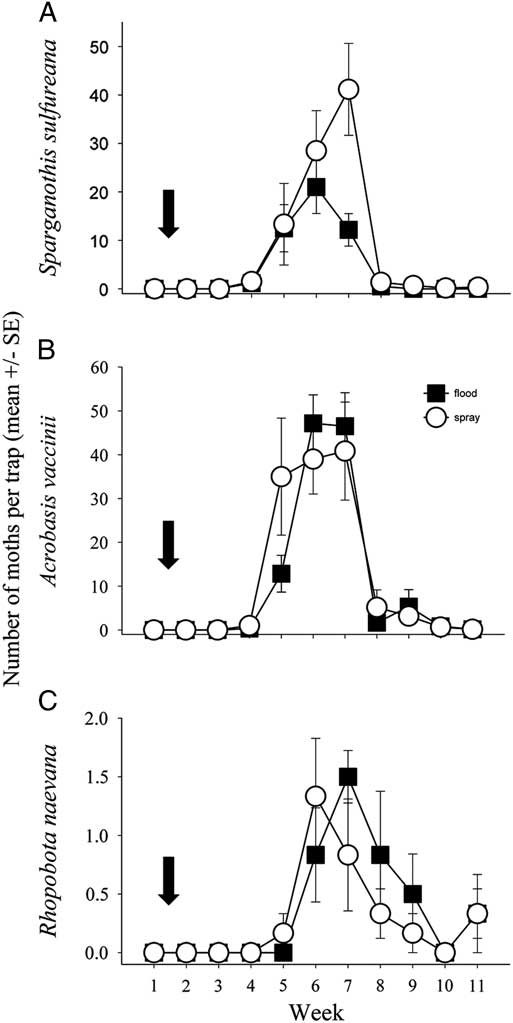
Fig. 2 Adult male moths (as a proxy for pest caterpillar densities) caught in pheromone-baited P2 traps. The open circles represent average moths caught in spray beds, while the black squares represent average moths caught in flood beds, of species (A) Sparganothis sulfureana, (B) Acrobasis vaccinii, and (C) Rhopobota naevana, averaged throughout the summer. The arrow signifies the time of the flood/spray treatment. SE, standard error.
Fewer than eight caterpillars were captured in either pitfall traps or sweep net samples, so these data were not analysed. Other herbivores caught using pitfall traps and sweep netting were in the subclass Acari, and insects from the orders Coleoptera (Chrysomelidae and Curculionidae), Hemiptera (Cicadellidae, Cercopidae, and Miridae), and Thysanoptera. These groups were not analysed, as they are not considered to be important pests of cranberry.
Detritivores
In the pre-treatment sample, the most abundant group of detritivores was Collembola, with captures up to 2000 per pitfall trap. There was a significant effect of week on Collembola numbers from both sweep net samples and pitfall traps (sweep: F(6,60)=4.703, P<0.001; pitfall: F(6,60)=41.302, P<0.001): initial population densities were high, then decreased to near zero by week 3, and subsequently stayed very low for the remainder of the sampling period (Fig. 3). In sweep net samples, Collembola abundance dropped by week 2, while in the understorey abundances stayed high for week 2, but decreased by week 3. There was no effect of treatment on Collembola density (pitfall: F(1,10)=0.053, P=0.823, Fig. 3; sweep: F(1,10)=0.232, P=0.64).
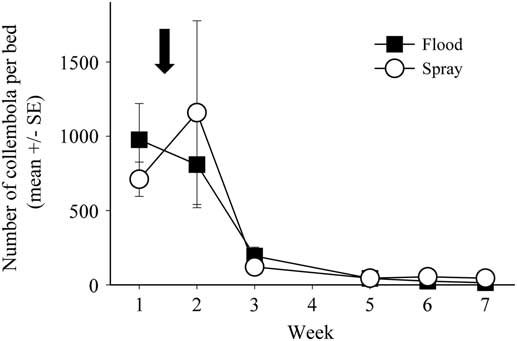
Fig. 3 Collembola activity-density, as a model for overall detritivore activity-density, as measured in pitfall traps across the seven-week sampling period. The open circles represent Collembola caught in spray beds, while the black squares represent Collembola caught in flood beds. The arrow signifies the time of the flood/spray treatment. No data were collected on week 4 due to traps flooding. SE, standard error.
The second most abundant of the detritivores were Acari, whose population trends, like Collembola, did not differ between flood and spray beds (pitfall: F(1,10)=0.364, P=0.56; sweep: F(1,10)=1.003, P=0.34). Other detritivores caught, at much lower densities, included Gastropods, Orthoptera (Gryllidae), Diptera (Nematoceran larvae), and several species of Coleoptera larvae.
Predators
The most abundant order of generalist predators in both pitfall traps and sweep net samples was Araneae (spiders). Although representing a complex and diverse group, for the purposes of this analysis all spiders were analysed together, and used as an indicator of generalist predators. This is based on the majority of feeding habits of spider species, and is used because specific classifications are not known for this agricultural ecosystem (Bardwell and Averill Reference Bardwell and Averill1997).
A total of 330 individual spiders were caught in sweep net samples, and there was no treatment effect on spider abundance (F(1,10)<0.001, P=0.995). However, in pitfall traps there were 700 total spiders captured, with significantly more spiders in the flooded than in the sprayed beds (F(1,10)=7.223, P=0.023; Fig. 4). The significant treatment×week interaction (F(5,50)=3.298, P=0.012) suggests that the effect of flooding on spider activity-density depended on the post-flood timing of sampling. Specifically, there was a significant difference in trap catch only during the first two weeks immediately following the flood/spray pest control event.
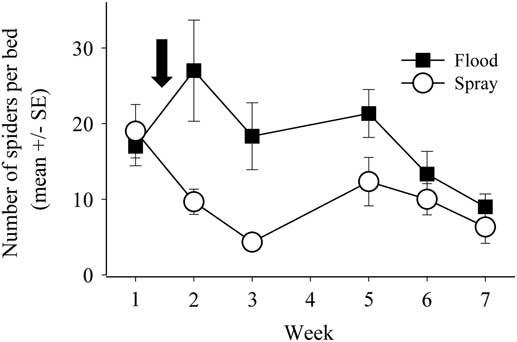
Fig. 4 Spider activity-density, as a model for overall generalist predator activity-density, as measured in pitfall traps across the seven-week sampling period. The open circles represent spiders caught in spray beds, while the black squares represent spiders caught in flood beds. The arrow signifies the time of the flood/spray treatment. No data were collected on week 4 due to traps flooding. SE, standard error.
Other generalist predators caught included Chilopoda, Diptera (Syrphidae larvae), and Coleoptera (Staphylinidae, Carabidae, and Lampyridae). These were present at 20% lower density than the spiders, and were not analysed.
Parasitoids
Throughout the summer, 79 parasitoid wasps were caught by sweep netting and 116 parasitoids were caught in pitfall traps. Most of the parasitoids analysed come from the families Ichneumonidae, Braconidae, and the superfamily Chalcidoidea. Because parasitoids abundances were low, week 2 through week 6 were summed together and analysed using a Mann–Whitney rank sum test. There were significantly more parasitoid wasps caught in the flooded than in the sprayed beds (U=468.50, n 1=n 2=42, P<0.001; Fig. 5).
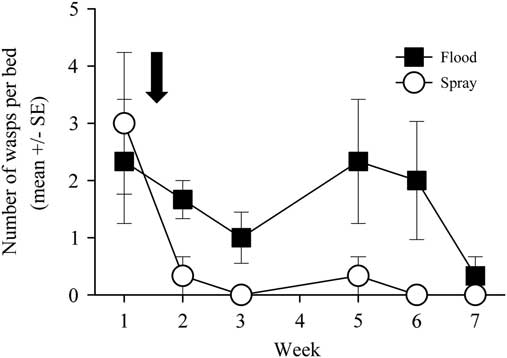
Fig. 5 Parasitoid wasp activity-density as measured in pitfall traps across the seven-week sampling period. The open circles represent parasitoid wasps caught in spray beds, while the black squares represent parasitoid wasps caught in flood beds. The arrow signifies the time of the flood/spray treatment. No data were collected on week 4 due to traps flooding. SE, standard error.
Discussion
In a Wisconsin commercial cranberry system, flooding was shown to be similar to a broad-spectrum insecticide application at controlling the three most economically important cranberry pest moths, S. sulfureana, A. vaccinii, and R. naevana (Fig. 2). This upholds findings from previous studies (Cockfield and Mahr Reference Cockfield and Mahr1992; Steffan et al. Reference Steffan, Singleton, Sokja, Zalapa, Dittl and Harbut2012b), and provides evidence that flooding is similar to a spring insecticide application in terms of pest suppression. Thus, we provide evidence in support of our hypothesis that biological and cultural control strategies are compatible with each other, and that, together, they are equivalent in efficacy to conventional insecticide use.
Another salient finding of this study is that the spring flood elevated the activity-density of natural enemies, specifically of spiders and parasitoid wasps, compared with the use of a spring insecticide application (Figs. 4–5). This finding offers insight into the mechanism through which flooding affects pest management, by looking at trophic dynamics in the marsh community. One possible mechanism through which pests are suppressed by the flood is indirectly, through the relative increase in the number of natural enemies present when using the spring flood compared with the traditional insecticide-based pest management programmes. Multiple studies have supported the notion that natural enemies, especially parasitoid wasps, are potentially important drivers in cranberry pest suppression (Marucci and Moulter Reference Marucci and Moulter1992; Mahr Reference Mahr1999). Observations suggest that 33–50% of S. sulfureana larvae are parasitised in Wisconsin commercial cranberry (Deutsch Reference Deutsch2014). Our finding that the cultural practice of spring flooding conserves natural enemies, more than the conventional practice of an insecticide application, suggests that the increased numbers of natural enemies had some influence on the impact of the flood on pest numbers.
Compatibility between flooding and natural enemy abundance, however, does not necessarily equate with pest suppression, given that many natural enemies also feed on detritivores as well as other predators (Straub et al. Reference Straub, Finke and Snyder2008). Many factors may undermine herbivore control, making it is possible that greater predator abundance and diversity would not result in an increase in herbivore suppression – predator community compositions often change, and with these shifts, the prey targeted may shift as well. Factors affecting herbivore control include predator-predator interference (Sih et al. Reference Sih, Englund and Wooster1998), intraguild predation (Polis Reference Polis1991; Finke and Denno Reference Finke and Denno2005), and variable predator hunting modes and ranges (Preisser et al. Reference Preisser, Orrock and Schmitz2007). However, given that the insecticide application itself controlled pests in the spray treatment, it is likely that the increased numbers of natural enemies in the flood treatment did in fact work to some extent in reducing pest pressure.
Spiders were by far the most abundant predators found, and were more common in flooded than sprayed beds. Because the feeding habits of spiders in commercial cranberry systems have not been well documented, it is unclear to what extent increased spider density would lead to a decrease in pest caterpillars. While many spiders are generalist predators and are likely to consume caterpillars opportunistically, some species of spiders are unlikely to encounter caterpillars on a regular basis (Bardwell and Averill Reference Bardwell and Averill1997). Identifying spiders to a finer taxonomic resolution, as well as directly studying their feeding habits or using gut-content analyses, would clarify the link between spider density and pest suppression.
The parasitoid wasps we collected belong largely to the superfamily Chalcidoidea, as well as several members of the Ichneumonidae and Braconidae. Because many parasitoids are host-specific or attack only a narrow group of species (Van den Bosch and Telford Reference Van den Bosch and Telford1964), they are unlikely to prey-switch away from consuming their target prey, and thus might not be strongly affected by the abundance or paucity of alternate prey species. Although the majority of these wasps parasitise moth larvae, they were not identified to a level of specificity to be sure of their host breadth. Pest caterpillar parasitism rates in flooded versus sprayed cranberry systems might be better assessed by rearing parasitoids from sentinel or wild-caught pest caterpillars. Rearing parasitoids in this way would control for variability in arthropod movement or differences in prey/host availability, and thereby demonstrate more accurately whether flooding conserves the specific parasitoids that target cranberry pest caterpillars.
We had expected that, by removing significant numbers of detritivores, the flood would eliminate this alternate prey resource and force hungry generalist predators to consume proportionally more herbivores, which is likely to include pest caterpillars (Fig. 1). Specifically, we had expected that the decline to near zero in Collembola populations, following both flood and spray treatments, would force spiders to prey-switch and drive down herbivore populations, most dramatically in the flooded beds that exhibited higher spider densities. However, we did not find any treatment effect on the abundance of herbivores or detritivores in this study. However, it is likely that, had we been able to distinguish between the direct effects of the flood and the spray, compared with indirect effects from the increase in natural enemies following the flood (such as with a no-flood, no-spray treatment), we may have found fewer herbivores in treatments with increased natural enemies present.
Altogether, our data show that there were greater numbers of natural enemies in flooded compared with sprayed beds, with both treatments leading to similar trends in pest moth and detritivore densities. Increased numbers of natural enemies in flooded beds is likely to increase biological control efforts, keeping herbivore populations in check and strengthening integrated pest management programmes. Thus, the spring flood in cranberries is not only compatible with biological control, but also appears to be similar in effectiveness to an insecticide application. Replacing a chemical control tactic with a cultural control tactic, which promotes carnivore survival, represents an example of a sustainable, best-management strategy. Further work in this vein may more directly elucidate the mechanisms driving the effect, and thereby provide evidence of conservation biological control.
Acknowledgements
Financial support for this work was provided by the Wisconsin Cranberry Board, the Cranberry Institute, and United State Department of Agriculture–Agricultural Research Service appropriated funds. The authors are also highly indebted to the commercial cranberry growers who allowed work to be done on their marshes, and to the many laboratory and field assistants without whose many hours of work this project would not have been possible.





