



Introduction
Conopidae (Diptera) larvae are obligate endoparasitoids (Gibson and Skevington Reference Gibson and Skevington2013; Gibson et al. Reference Gibson, Skevington and Kelso2013). Stylogaster Macquart (Diptera: Conopidae) is the sister group to other conopids (Marshall Reference Marshall2012; Gibson and Skevington Reference Gibson and Skevington2013). Stylogaster species target nymphs and adult Gryllidae (Orthoptera) and Blattodea, whereas other conopid genera use adult aculeate Hymenoptera (Freeman Reference Freeman1966; Kotrba Reference Kotrba1997; Woodley and Judd Reference Woodley and Judd1998; Schmid-Hempel Reference Schmid-Hempel2001; Rasmussen and Cameron Reference Rasmussen and Cameron2004; Marshall Reference Marshall2012; Gibson et al. Reference Gibson, Skevington and Kelso2013). Stylogaster eggs have been found on other species, such as other Diptera, but the lack of egg development indicates that these may be accidental ovipositions (Couri and Barros Reference Couri and Barros2010; Stuke Reference Stuke2012; Couri et al. Reference Couri, Jordaens, Geeraert, Matheus and Vieira-Araújo2019).
Stylogaster species have been studied mainly in Africa and the Neotropics and are associated with army ants (Hymenoptera: Formicidae), using hosts that have been flushed out by the ants (Lopes Reference Lopes1937; Rettenmyer Reference Rettenmyer1961; Burt et al. Reference Burt, Skevington and Rocha2015). The flies are thought to either stab their ovipositors into the abdomen of the host and insert the egg under a sclerite or shoot the egg from the ovipositor via a flipping motion of the abdomen (Kotrba Reference Kotrba1997; Woodley and Judd Reference Woodley and Judd1998). While most studies have focussed on tropical species, Stylogaster neglecta, S. biannulata (Say), and S. beresfordi Burt are found in the Nearctic region (including Canada), typically in forest understoreys where adults feed on yellow/white flowers (Burt et al. Reference Burt, Skevington and Rocha2015). Only S. biannulata has a recorded host, Gryllus rubens Scudder (Orthoptera: Gryllidae) (Woodley and Judd Reference Woodley and Judd1998). While incubation times were not examined, Woodley and Judd (Reference Woodley and Judd1998) found that in the few cases where adult flies emerged, it was 19–25 days after the puparia were formed. While little is known of the life history of the genus Stylogaster, the related conopids Sicus Latreille and Physocephala Schiner (Diptera: Conopidae), which parasitise Bombus Latreille (Hymenoptera: Apidae) in Europe, North America, and Japan, oviposit during periods of host reproduction (Schmid-Hempel Reference Schmid-Hempel2001) from July until mid-September (Macfarlane et al. Reference Macfarlane, Lipa and Liu1995). They kill their hosts within 10–12 days of oviposition. Hosts are induced to burrow into the ground and the fly pupates inside the dead host (Schmid-Hempel Reference Schmid-Hempel2001).
In old fields at the University of Toronto at Mississauga in August 1996, one of us (W.B.D.) first found Stylogaster neglecta larvae (identified by Jeff Skevington, Canadian National Collection of Insects, Arachnids, and Nematodes, Ottawa, Ontario, Canada) emerging from juvenile Oecanthus nigricornis Walker (Orthoptera: Gryllidae) (black-horned tree crickets), pupating, and then eclosing (Fig. 1). The larvae emerged and pupated between late August and mid-September, with adults eclosing from late November to early January (L.F.B., unpublished data). Oecanthus nigricornis are found across eastern North America, hatching around mid-June and reaching maturity in August. Eggs are laid in August and September and hatch the following June (Fulton Reference Fulton1915; Vickery and Kevan Reference Vickery and Kevan1985). Oecanthus are parasitised by several hymenopteran egg parasitoids (Smith Reference Smith1930a, Reference Smith1930b; Udine and Pinckney Reference Udine and Pinckney1940) and some helminths (Denner Reference Denner1968; Sonin Reference Sonin1990). However, there are currently no known dipteran parasitoids of the genus, despite other Orthoptera (including gryllids) commonly being parasitised by Sarcophagidae (Diptera) and Tachinidae (Diptera). This includes the well-known genus Ormia Robineau-Desvoidy, which is attracted to songs of male gryllid hosts (Rees Reference Rees1973; Cade Reference Cade1975; Lehmann Reference Lehmann2003). Our observations are the first case to our knowledge of Oecanthus juveniles or adults as hosts for a dipteran parasitoid and the first report of a host for S. neglecta.
We collected further O. nigricornis samples at the University of Toronto at Mississauga (Ontario, Canada) in the summer of 2017 and 2018 and identified emerging adults as S. neglecta using Burt et al. (Reference Burt, Skevington and Rocha2015). In 2017, O. nigricornis were collected from natural populations at discrete time intervals to determine when in the season conopid parasitism occurs. In 2018, O. nigricornis were sampled from several sites to determine population-level variation in parasitism.
Methods
Natural history of Stylogaster neglecta
In 2017, we sampled O. nigricornis approximately every two weeks from 28 June until 13 September (Table 1–2) from old fields at the University of Toronto, Mississauga Campus (43.55°N, 79.66°W) (total = 131 O. nigricornis). The start date was chosen as the first day in which we were successfully able to sample O. nigricornis from the site. The end date was chosen as the first sample in which we no longer found any S. neglecta larva emerging from the sampled O. nigricornis. We reared O. nigricornis individually in open-ended plastic cylinders, 8 cm in diameter and 7.5 cm long, with metal mesh covering the top end, at 25 °C and 70% humidity. Oecanthus nigricornis were fed pollen and dry dog food, and containers were sprayed with water every second day. We recorded the date on which O. nigricornis were caught, when they died, and whether a parasitoid emerged. Stylogaster neglecta pupae found in the containers were removed and placed in a petri dish on filter paper and sprayed with water daily. Chi-squared tests were used to determine if there were significant differences in the proportion of O. nigricornis parasitised on different sampling dates. Confidence intervals (CIs) were calculated using the Clopper–Pearson method. Cases of multiple conopid parasitism of a single host were also recorded.

Fig. 1. Stylogaster neglecta female raised from a pupa after emerging from Oecanthus nigricornis. Collected as a pupa on 28 August 1996 by Luc Bussière and emerged as an adult 9 January 1997.
Table 1. Sex, location, voucher identification number, and collector of samples used in analysis and referenced.

Table 2. Dates of collections done over the summer of 2017, with number of samples collected and number of parasitised individuals in each collection.

To examine whether S. neglecta was actively laying eggs the entire summer or during a shorter period of time, an analysis of variance was performed to determine if the length of time between when we collected the host and parasitoid emergence differed between collection dates. If S. neglecta oviposits on hosts throughout the summer, then larvae in the sampled O. nigricornis should be at different stages of development, and there will be no correlation between date of sampling and length of time from host capture until parasitoid emergence. If, however, S. neglecta females oviposited roughly concurrently, then we expect O. nigricornis sampled on the same date would have parasitoids that emerge after similar lengths of time and that the length of time between when the host O. nigricornis was sampled and the parasitoid emerging will be longer earlier in the summer and shorten as the season progresses. Specimens were deposited at the Royal Ontario Museum (Toronto, Ontario, Canada).
Site differences in parasitism
In late July 2018, we collected O. nigricornis in southern Ontario (Canada) and New York State (United States of America): from the Koffler Scientific Reserve (King City Ontario, 44.03°N, 79.53°W), rare Charitable Research Reserve (referred to as rare) (Cambridge Ontario, 43.38°N, 80.39°W), our University of Toronto at Mississauga site previously sampled in 2017, and the Edmund Niles Huyck Preserve and Biological Research Station (referred to as Huyck) (Rensselaerville, New York, 42.10°N, 74.10°W). Late July was chosen based on the results from the 2017 study, which indicated that this was the peak of S. neglecta oviposition activity. For each site, two to four old fields within a 1-km radius of each other were sampled by sweep netting. Oecanthus nigricornis caught ranged from third to fifth instar. Oecanthus nigricornis were frozen and then preserved in 70% ethanol before being dissected, and their abdomens examined using a Wild M5A dissecting microscope (Wetzlar, Hesse, Germany). They were scored as parasitised if S. neglecta eggs or larvae were found (Fig. 2). A chi-squared test was performed to determine if there were significant differences in the proportion of infected O. nigricornis between sites. Confidence intervals were calculated using the Clopper–Pearson method.
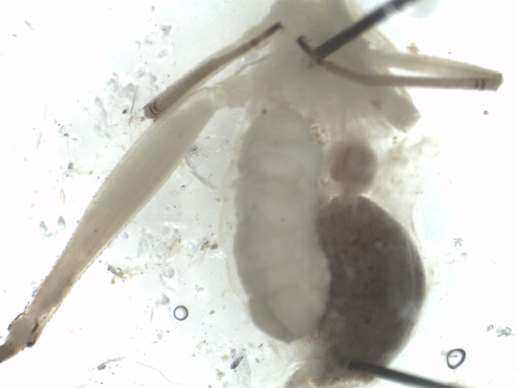
Fig. 2. Juvenile fourth instar Oecanthus nigricornis, parasitised by Stylogaster neglecta. Caught at rare Charitable Research Reserve.
Results
Natural history of Stylogaster neglecta
Twenty-four (20%) of the 120 University of Toronto at Mississauga O. nigricornis were parasitised by S. neglecta. There was a significant difference in the proportion of parasitised O. nigricornis collected on different dates (χ 2 = 37.014, df = 2, P < 0.01) with the highest proportion of parasitised individuals found in the 14 August sample (Fig. 3). Parasitoid larvae emerged from the near the anus of O. nigricornis and pupated outside the host. Host O. nigricornis remained alive for hours to days after emergence, and several observations of O. nigricornis after parasitoid emergence suggested that they had lost the use of their metathoracic legs. There were several cases of multiple parasitism: four O. nigricornis had two S. neglecta larvae, and one hosted three. Parasitoids that shared a host appeared to be smaller than parasitoids that did not (personal observation). Of the pupae, only one eclosed (in December 2017). Mean time from a O. nigricornis being collected to parasitoid emergence was 16.6 days (SD = 9.04; range 3–31 days) (Fig. 4). No O. nigricornis collected before 17 July or after 29 August were parasitised. Consistent with this narrow oviposition period, mean time from host collection to parasitoid emergence was significantly shorter at later sampling dates (F = 32.36, P < 0.001) (Fig. 5).

Fig. 3. Proportion of Oecanthus nigricornis collected in the summer of 2017 that were parasitised at the University of Toronto at Mississauga, with exact binomial confidence intervals and sample sizes.

Fig. 4. Time in days between collection of parasitised Oecanthus nigricornis (University of Toronto at Mississauga 2017) and when larval Stylogaster neglecta emerged for all O. nigricornis.
Site differences in parasitism
Of 674 O. nigricornis from all four sites, 149 or 22% (CI = 0.246, 0.324) were parasitised. There were large differences in parasitism between sites (Fig. 6): at Huyck 2% (n = 104, CI = 0.0024, 0.069), at Koffler 11% (n = 178, CI = 0.079, 0.189), at rare 26% (n = 193, CI = 0.272, 0.434), and at University of Toronto at Mississauga 39% (n = 199, CI = 0.539, 0.717) (χ 2 = 70.23, P < 0.001). Oecanthus nigricornis hosts were mostly third to fifth instars, and of all O. nigricornis parasitised, only one had reached adulthood.

Fig. 5. Time in days between collection of parasitised Oecanthus nigricornis (University of Toronto at Mississauga 2017) and when larval Stylogaster neglecta emerged, averaged for each collection date, with standard deviation.

Fig. 6. Proportion of Oecanthus nigricornis parasitised from samples collected across four sites in July 2018. Exact binomial confidence intervals and sample sizes are given. Huyck, Edmund Niles Huyck Preserve and Biological Research Station; KSR, Koffler Scientific Reserve; UTM, University of Toronto at Mississauga; rare, rare Charitable Research Reserve.
Discussion
We report S. neglecta as the first dipteran parasitoid of O. nigricornis, and this tree cricket as the first known host for this parasitoid. Like S. biannulata, S. neglecta uses a gryllid cricket host in which the larvae develop internally, pupates outside the host, and occasionally exhibits superparasitism (Woodley and Judd Reference Woodley and Judd1998). Like the Bombus specialists Physocephala and Sicus, S. neglecta have a narrow time window of a few weeks in which hosts are parasitised (Macfarlane et al. Reference Macfarlane, Lipa and Liu1995; Schmid-Hempel Reference Schmid-Hempel2001; Otterstatter Reference Otterstatter2004). Unlike the Bombus-hunting conopids, S. neglecta pupates outside of the host. Stylogaster neglecta also appear to have longer incubation times than the 10–12 days reported for the Bombus hunters (Schmid-Hempel Reference Schmid-Hempel2001). Further work should examine the nature of overwintering strategies in S. neglecta and whether it resembles that of other conopids.
The time from host capture until parasitoid emergence decreased over the season with longer intervals in July and shorter ones in August. We did not find any parasitised O. nigricornis in June or September. The shorter intervals in the late summer are because the larvae had already been developing for some time before we collected their hosts. Stylogaster neglecta appears to parasitise hosts from late July to early August and is not active throughout the rest of the summer. This is consistent with unpublished adult collection records associated with the Burt et al. (Reference Burt, Skevington and Rocha2015) revision, which examined records of adult S. neglecta in collections across North America. Of the 128 S. neglecta caught in southern Ontario, eight were caught in June, 114 in July, five in August, and one in September. Such a narrow time of parasitoid adult activity is expected in hosts with discrete generation times (Godfray Reference Godfray1994). Parasitoids that emerge too early may suffer from greater mortality waiting until hosts emerge, while those emerging late may have difficulty either in finding unparasitised hosts or in subduing larger-sized hosts.
Assuming that all S. neglecta larvae have a similar incubation times, and the longer lengths of time from host capture until parasitoid emergence are roughly equivalent to the incubation time, then it would appear that S. neglecta has an incubation period of about 30 days in its host. This estimate is higher than estimates for other conopid genera, such as 10–12 days found in Physocephala and Sicus (Schmid-Hempel Reference Schmid-Hempel2001). However, as noted earlier, the mean time from collecting the host O. nigricornis until parasitoid emergence will be shorter than actual mean incubation time given that eggs had been laid before collection. The actual incubation time is likely to be longer than we recorded but less than 44 days, as there were no indications of parasitism two weeks before the earliest sample.
We found significant differences in the proportion of O. nigricornis that were parasitised across four different sites in southern Ontario and upstate New York. The proportion of parasitised individuals ranged from 39% in University of Toronto at Mississauga to 2% in the Huyck Preserve. While there were significant differences between sites in terms of parasitism rate, our sampling only took place over a single season, so we cannot confirm if these are long-term site differences. There appears to be an inter-year difference in parasitism at University of Toronto at Mississauga, with the parasitism rate in late July 2018 being higher than the parasitism rates found in July 2017. However, the parasitism rate in August 2017 was higher than the single 2018 sample. It appears that the rate of parasitism at University of Toronto at Mississauga peaked slightly earlier in 2018 than in 2017. Parasitoid–host relationships often show cyclical population dynamics across multiple years (Hassell and May Reference Hassell and May1973; Godfray Reference Godfray1994; Godfray et al. Reference Godfray, Hassell and Holt1994), which may explain the between-site differences. Long-term sampling will be necessary to determine if these differences are consistent from year to year.
Acknowledgements
The authors thank John Ratcliffe, Helene Wagner, Nicole Mideo, Andrew Mason, and Ahmed Hasan for their support and edits when preparing this manuscript; the rare Charitable Research Reserve, Edmund Niles Huyck Preserve, and Koffler Scientific Reserve for allowing us to work at their sites as well as their respective contacts at each location; and Jenna Quinn, Anne Rhoads, Adam Caprio, Stephan Schneider, and John Stinchcombe for facilitating the process. The authors thank the undergraduate students in our laboratory, Ali Al Wafi, Kavya Manikonda, Alanah Joyce, and Samantha Hasbum, who helped collect samples. The authors also thank Jessica Browne for her help in the field and comments on the manuscript.







