


1 Introduction
Bacterial suspensions, one of the many realizations of active matter, have been the subject of intense study over the last two decades, both at the level of a single organism (Lauga & Powers Reference Lauga and Powers2009) and with regard to emergent phenomena over large length and time scales (Koch & Subramanian Reference Koch and Subramanian2011; Subramanian & Nott Reference Subramanian and Nott2011). Among other interesting characteristics, the rheological properties of such suspensions are of particular interest, since they are in sharp contrast to that of near-equilibrium complex fluids. In a recent paper (Lopez et al.
Reference Lopez, Gachelin, Douarche, Auradou and Clement2015), dilute suspensions of a pair of strains of wild type E. coli were subjected to uniform shearing flow in a sensitive Couette rheometer over a range of volume fractions and shear rates. For volume fractions corresponding to the dilute regime,
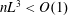 $nL^{3}<O(1)$
,
$nL^{3}<O(1)$
,
 $n$
being the bacterial number density and
$n$
being the bacterial number density and
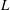 $L$
being the total bacterium length (including both head and flagellum), and for shear rates small compared to the inverse orientation relaxation time (on account of tumbling and rotary diffusion), a schematic of the stress response resulting from the imposition of a constant shear rate for a finite period is shown in figure 1(b). The response includes novel regimes, both steady and transient, of negative shear stresses, a fact attributed to bacterial activity. This response is readily understood, in qualitative terms, based on the known response of a complex fluid with a passive microstructure (Brownian fibres, for instance) that, on deformation, develops a net contractile dipole in response to the same shear rate history. The passive-fluid response is shown in figure 1(a). An instantaneous stress jump corresponding to a viscous contribution (that includes both a solvent and a hydrodynamic microstructural component) arises on the start-up of shear, and this is followed by a further increase to a higher plateau due to an elastic contribution that develops on a time scale of order the microstructural relaxation time, and is indicative of the microstructure approaching an anisotropic steady state with a net contractile dipole (the inextensible Brownian fibres, on average, align with the ambient extensional axis). The cessation of shear leads to an instantaneous drop due to the disappearance of the viscous contribution, followed by the decay of the elastic component to zero. Motile organisms such as spermatozoans, and bacteria such as E. coli, B. subtilis etc., termed pushers, have extensile intrinsic force dipoles that are of an opposite sign to those that develop in a passive microstructure in response to shearing. For small shear rates, the dominant disperse phase stress in a bacterial suspension is expected to arise from the anisotropy of the intrinsic force-dipole orientations, and has an elastic character with a sign opposite to the passive case. Thus, merely changing the sign of the time-varying elastic stress component in the passive response described above rationalizes the experimentally observed response including, in particular, negative stresses during steady shearing above a threshold volume fraction and transient negative stresses following shear cessation for any non-zero volume fraction. The simple explanation above is not offered by the authors of the said article (Lopez et al.
Reference Lopez, Gachelin, Douarche, Auradou and Clement2015) and is also missing in a later comment on the same experiments (Marchetti Reference Marchetti2015).
$L$
being the total bacterium length (including both head and flagellum), and for shear rates small compared to the inverse orientation relaxation time (on account of tumbling and rotary diffusion), a schematic of the stress response resulting from the imposition of a constant shear rate for a finite period is shown in figure 1(b). The response includes novel regimes, both steady and transient, of negative shear stresses, a fact attributed to bacterial activity. This response is readily understood, in qualitative terms, based on the known response of a complex fluid with a passive microstructure (Brownian fibres, for instance) that, on deformation, develops a net contractile dipole in response to the same shear rate history. The passive-fluid response is shown in figure 1(a). An instantaneous stress jump corresponding to a viscous contribution (that includes both a solvent and a hydrodynamic microstructural component) arises on the start-up of shear, and this is followed by a further increase to a higher plateau due to an elastic contribution that develops on a time scale of order the microstructural relaxation time, and is indicative of the microstructure approaching an anisotropic steady state with a net contractile dipole (the inextensible Brownian fibres, on average, align with the ambient extensional axis). The cessation of shear leads to an instantaneous drop due to the disappearance of the viscous contribution, followed by the decay of the elastic component to zero. Motile organisms such as spermatozoans, and bacteria such as E. coli, B. subtilis etc., termed pushers, have extensile intrinsic force dipoles that are of an opposite sign to those that develop in a passive microstructure in response to shearing. For small shear rates, the dominant disperse phase stress in a bacterial suspension is expected to arise from the anisotropy of the intrinsic force-dipole orientations, and has an elastic character with a sign opposite to the passive case. Thus, merely changing the sign of the time-varying elastic stress component in the passive response described above rationalizes the experimentally observed response including, in particular, negative stresses during steady shearing above a threshold volume fraction and transient negative stresses following shear cessation for any non-zero volume fraction. The simple explanation above is not offered by the authors of the said article (Lopez et al.
Reference Lopez, Gachelin, Douarche, Auradou and Clement2015) and is also missing in a later comment on the same experiments (Marchetti Reference Marchetti2015).

Figure 1. The shear stress response of passive (a) and active (b) suspensions in response to a step shear of a finite duration.
In § 2, starting from the governing kinetic equation for the orientation probability density of a single bacterium in the presence of a shearing flow and relaxation processes due to run-and-tumble dynamics and rotary diffusion, we calculate, at leading order in the shear rate, the anisotropic probability density that develops in response to a weak time-dependent simple shear flow. This probability density is then used to determine the shear stress response in a dilute non-interacting bacterial suspension, including both active (arising due to the aforementioned anisotropy of the intrinsic force dipoles) and passive (arising due to the near inextensibility of the bacterial flagellum in weak flows Darnton et al. Reference Darnton, Turner, Rojevsky and Berg2007) components. In § 3, a quantitative calculation of the stress response is carried out for the specific case where the suspension is subject to a steady simple shearing flow over a finite time period, and the results are compared with the aforementioned experiments. The comparison is restricted to the small-shear-rate regime. For strong flows with shear rates greater than the characteristic inverse relaxation time, the passive stress component, that arises from induced dipoles proportional to the shear rate, will likely be dominant and the response starts to resemble that of a passive fluid. This is indeed observed in the said experiments where one sees a transition in the stress response from that in figure 1(b) to the one in figure 1(a) with increasing shear rate (Lopez et al. Reference Lopez, Gachelin, Douarche, Auradou and Clement2015). Further, a calculation of the viscosity for arbitrary shear rates would necessarily involve an elaborate numerical effort similar to those carried out earlier for passive complex fluids (Brenner Reference Brenner1974), and is beyond the scope of this brief communication.
2 The stress in a dilute bacterial suspension
The bulk stress in a suspension of orientable particles requires knowledge of the symmetric part of the first moment of the traction on their surfaces, called the stresslet (Batchelor Reference Batchelor1970b ), and the distribution of particle orientations. In what follows, we first consider the latter for the bacterial suspension (§ 2.1), and then provide a relation for the stress (§ 2.2).
2.1 The distribution of bacterial orientations
The orientation probability density in a homogeneously sheared suspension of bacteria that swim with speed
 $U$
and undergo a combination of run-and-tumble dynamics and rotary diffusion, satisfies the equation (Subramanian & Koch Reference Subramanian and Koch2009)
$U$
and undergo a combination of run-and-tumble dynamics and rotary diffusion, satisfies the equation (Subramanian & Koch Reference Subramanian and Koch2009)
 $$\begin{eqnarray}\frac{\unicode[STIX]{x2202}\unicode[STIX]{x1D6FA}}{\unicode[STIX]{x2202}t}+\frac{1}{\unicode[STIX]{x1D70F}}\left(\unicode[STIX]{x1D6FA}-\int K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })\unicode[STIX]{x1D6FA}(\boldsymbol{p}^{\prime },t)\,\text{d}\boldsymbol{p}^{\prime }\right)-D_{r}\unicode[STIX]{x1D6FB}_{\boldsymbol{p}}^{2}\unicode[STIX]{x1D6FA}=-\unicode[STIX]{x1D735}_{\boldsymbol{ p}}\boldsymbol{\cdot }(\dot{\boldsymbol{p}}\unicode[STIX]{x1D6FA}),\end{eqnarray}$$
$$\begin{eqnarray}\frac{\unicode[STIX]{x2202}\unicode[STIX]{x1D6FA}}{\unicode[STIX]{x2202}t}+\frac{1}{\unicode[STIX]{x1D70F}}\left(\unicode[STIX]{x1D6FA}-\int K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })\unicode[STIX]{x1D6FA}(\boldsymbol{p}^{\prime },t)\,\text{d}\boldsymbol{p}^{\prime }\right)-D_{r}\unicode[STIX]{x1D6FB}_{\boldsymbol{p}}^{2}\unicode[STIX]{x1D6FA}=-\unicode[STIX]{x1D735}_{\boldsymbol{ p}}\boldsymbol{\cdot }(\dot{\boldsymbol{p}}\unicode[STIX]{x1D6FA}),\end{eqnarray}$$
where
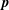 $\boldsymbol{p}$
denotes the bacterium orientation and
$\boldsymbol{p}$
denotes the bacterium orientation and
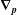 $\unicode[STIX]{x1D735}_{\boldsymbol{p}}$
is the gradient operator in orientation space. The spatial variation of
$\unicode[STIX]{x1D735}_{\boldsymbol{p}}$
is the gradient operator in orientation space. The spatial variation of
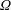 $\unicode[STIX]{x1D6FA}$
on account of swimming with velocity
$\unicode[STIX]{x1D6FA}$
on account of swimming with velocity
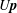 $U\boldsymbol{p}$
,
$U\boldsymbol{p}$
,
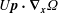 $U\boldsymbol{p}\boldsymbol{\cdot }\unicode[STIX]{x1D735}_{\boldsymbol{x}}\unicode[STIX]{x1D6FA}$
, has been omitted due to spatial homogeneity of the imposed flow. In (2.1),
$U\boldsymbol{p}\boldsymbol{\cdot }\unicode[STIX]{x1D735}_{\boldsymbol{x}}\unicode[STIX]{x1D6FA}$
, has been omitted due to spatial homogeneity of the imposed flow. In (2.1),
 $\unicode[STIX]{x1D70F}$
represents the mean run time of the bacterium between successive tumbles with the tumble events being governed by Poisson statistics, and
$\unicode[STIX]{x1D70F}$
represents the mean run time of the bacterium between successive tumbles with the tumble events being governed by Poisson statistics, and
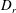 $D_{r}$
represents the rotary diffusivity which characterizes the continuous (athermal) fluctuations in orientation during the run phase. The tumble events are assumed to be instantaneous here with the kernel
$D_{r}$
represents the rotary diffusivity which characterizes the continuous (athermal) fluctuations in orientation during the run phase. The tumble events are assumed to be instantaneous here with the kernel
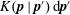 $K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })\,\text{d}\boldsymbol{p}^{\prime }$
in the integral term in (2.1) being the probability density associated with finding a bacterium with post-tumble orientation
$K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })\,\text{d}\boldsymbol{p}^{\prime }$
in the integral term in (2.1) being the probability density associated with finding a bacterium with post-tumble orientation
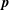 $\boldsymbol{p}$
starting from a pre-tumble orientation of
$\boldsymbol{p}$
starting from a pre-tumble orientation of
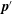 $\boldsymbol{p}^{\prime }$
. Conservation of bacteria during tumble events implies
$\boldsymbol{p}^{\prime }$
. Conservation of bacteria during tumble events implies
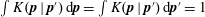 $\int K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })\,\text{d}\boldsymbol{p}=\int K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })\,\text{d}\boldsymbol{p}^{\prime }=1$
, and a convenient form for the transition probability density is
$\int K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })\,\text{d}\boldsymbol{p}=\int K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })\,\text{d}\boldsymbol{p}^{\prime }=1$
, and a convenient form for the transition probability density is
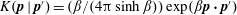 $K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })=(\unicode[STIX]{x1D6FD}/(4\unicode[STIX]{x03C0}\sinh \unicode[STIX]{x1D6FD}))\exp (\unicode[STIX]{x1D6FD}\boldsymbol{p}\boldsymbol{\cdot }\boldsymbol{p}^{\prime })$
where
$K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })=(\unicode[STIX]{x1D6FD}/(4\unicode[STIX]{x03C0}\sinh \unicode[STIX]{x1D6FD}))\exp (\unicode[STIX]{x1D6FD}\boldsymbol{p}\boldsymbol{\cdot }\boldsymbol{p}^{\prime })$
where
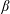 $\unicode[STIX]{x1D6FD}$
is a correlation parameter (Subramanian & Koch Reference Subramanian and Koch2009). The limit
$\unicode[STIX]{x1D6FD}$
is a correlation parameter (Subramanian & Koch Reference Subramanian and Koch2009). The limit
 $\unicode[STIX]{x1D6FD}$
$\unicode[STIX]{x1D6FD}$
 $\rightarrow$
0 corresponds to a random tumble with
$\rightarrow$
0 corresponds to a random tumble with
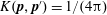 $K(\boldsymbol{p},\boldsymbol{p}^{\prime })=1/(4\unicode[STIX]{x03C0})$
, while
$K(\boldsymbol{p},\boldsymbol{p}^{\prime })=1/(4\unicode[STIX]{x03C0})$
, while
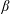 $\unicode[STIX]{x1D6FD}$
$\unicode[STIX]{x1D6FD}$
 $\rightarrow$
$\rightarrow$
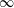 $\infty$
corresponds to smooth swimmers that do not tumble, and whose orientation decorrelates solely due to rotary diffusion. For E. coli, the tumble events typically have a forward bias with
$\infty$
corresponds to smooth swimmers that do not tumble, and whose orientation decorrelates solely due to rotary diffusion. For E. coli, the tumble events typically have a forward bias with
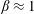 $\unicode[STIX]{x1D6FD}\approx 1$
, implying a mean change in orientation of about
$\unicode[STIX]{x1D6FD}\approx 1$
, implying a mean change in orientation of about
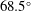 $68.5^{\circ }$
(Berg Reference Berg1993); this average change in orientation accompanying a tumble is an alternative measure of correlation and has also been used earlier to statistically characterize run-and-tumble motion (Lovely & Dahlquist Reference Lovely and Dahlquist1975). In the absence of a flow, correlated tumbling and rotary diffusion together lead to a relaxation towards an isotropic orientation distribution with
$68.5^{\circ }$
(Berg Reference Berg1993); this average change in orientation accompanying a tumble is an alternative measure of correlation and has also been used earlier to statistically characterize run-and-tumble motion (Lovely & Dahlquist Reference Lovely and Dahlquist1975). In the absence of a flow, correlated tumbling and rotary diffusion together lead to a relaxation towards an isotropic orientation distribution with
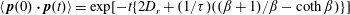 $\langle \,\boldsymbol{p}(0)\boldsymbol{\cdot }\boldsymbol{p}(t)\rangle =\text{exp}[-t\{2D_{r}+(1/\unicode[STIX]{x1D70F})((\unicode[STIX]{x1D6FD}+1)/\unicode[STIX]{x1D6FD}-\coth \unicode[STIX]{x1D6FD})\}]$
. The term involving
$\langle \,\boldsymbol{p}(0)\boldsymbol{\cdot }\boldsymbol{p}(t)\rangle =\text{exp}[-t\{2D_{r}+(1/\unicode[STIX]{x1D70F})((\unicode[STIX]{x1D6FD}+1)/\unicode[STIX]{x1D6FD}-\coth \unicode[STIX]{x1D6FD})\}]$
. The term involving
 $\dot{\boldsymbol{p}}$
in (2.1) describes the rotation of the bacterium due to the ambient linear flow field, and for a slender body, is given by Jeffery (Reference Jeffery1922):
$\dot{\boldsymbol{p}}$
in (2.1) describes the rotation of the bacterium due to the ambient linear flow field, and for a slender body, is given by Jeffery (Reference Jeffery1922):
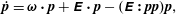 $$\begin{eqnarray}\dot{\boldsymbol{p}}=\unicode[STIX]{x1D74E}\boldsymbol{\cdot }\boldsymbol{p}+\unicode[STIX]{x1D640}\boldsymbol{\cdot }\boldsymbol{p}-(\unicode[STIX]{x1D640}\boldsymbol{ : }\boldsymbol{p}\boldsymbol{p})\boldsymbol{p},\end{eqnarray}$$
$$\begin{eqnarray}\dot{\boldsymbol{p}}=\unicode[STIX]{x1D74E}\boldsymbol{\cdot }\boldsymbol{p}+\unicode[STIX]{x1D640}\boldsymbol{\cdot }\boldsymbol{p}-(\unicode[STIX]{x1D640}\boldsymbol{ : }\boldsymbol{p}\boldsymbol{p})\boldsymbol{p},\end{eqnarray}$$
at leading order in the aspect ratio, where
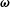 $\unicode[STIX]{x1D74E}$
and
$\unicode[STIX]{x1D74E}$
and
 $\unicode[STIX]{x1D640}$
are the time-dependent vorticity and rate of strain tensors of the imposed flow.
$\unicode[STIX]{x1D640}$
are the time-dependent vorticity and rate of strain tensors of the imposed flow.
The assumption of spatial homogeneity in (2.1) neglects effects related to the accumulation of microscopic swimmers near boundaries observed in experiments (Berke et al.
Reference Berke, Turner, Berg and Lauga2008), and attributed to both kinematic (collision with boundary followed by alignment; Elgeti & Gompper Reference Elgeti and Gompper2013; Elgeti & Gompper Reference Elgeti and Gompper2015) and hydrodynamic mechanisms (image-induced attraction; Hernandez-Ortiz, Stoltz & Graham Reference Hernandez-Ortiz, Stoltz and Graham2005; Berke et al.
Reference Berke, Turner, Berg and Lauga2008). Such preferential accumulation is, however, largely restricted to kinetic boundary layers of order the bacterium mean free path (
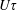 $U\unicode[STIX]{x1D70F}$
). For the experiments in question (Lopez et al.
Reference Lopez, Gachelin, Douarche, Auradou and Clement2015), the gap width of the Couette cell is approximately thirty times greater than
$U\unicode[STIX]{x1D70F}$
). For the experiments in question (Lopez et al.
Reference Lopez, Gachelin, Douarche, Auradou and Clement2015), the gap width of the Couette cell is approximately thirty times greater than
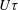 $U\unicode[STIX]{x1D70F}$
, implying that effects arising from the accumulation of highly aligned bacteria close to boundaries would have a negligible effect on the measured bulk rheology.
$U\unicode[STIX]{x1D70F}$
, implying that effects arising from the accumulation of highly aligned bacteria close to boundaries would have a negligible effect on the measured bulk rheology.
2.2 The bulk stress in a bacterial suspension
Knowledge of the orientation probability density from (2.1) allows one to calculate the deviatoric part of the bulk suspension stress as (Batchelor Reference Batchelor1970b ):
 $$\begin{eqnarray}\langle \unicode[STIX]{x1D748}\rangle =2\unicode[STIX]{x1D707}\unicode[STIX]{x1D640}+\langle \unicode[STIX]{x1D748}_{b}\rangle ,\end{eqnarray}$$
$$\begin{eqnarray}\langle \unicode[STIX]{x1D748}\rangle =2\unicode[STIX]{x1D707}\unicode[STIX]{x1D640}+\langle \unicode[STIX]{x1D748}_{b}\rangle ,\end{eqnarray}$$
where
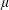 $\unicode[STIX]{x1D707}$
is the solvent viscosity and the disperse (bacterial) phase contribution to the stress is given by:
$\unicode[STIX]{x1D707}$
is the solvent viscosity and the disperse (bacterial) phase contribution to the stress is given by:
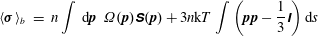 $$\begin{eqnarray}\displaystyle \langle \unicode[STIX]{x1D748}\rangle _{b} & = & \displaystyle n\displaystyle \int \,\text{d}\boldsymbol{p}~\unicode[STIX]{x1D6FA}(\boldsymbol{p})\unicode[STIX]{x1D64E}(\boldsymbol{p})+3n\text{k}T\displaystyle \int \left(\boldsymbol{p}\boldsymbol{p}-\frac{1}{3}\unicode[STIX]{x1D644}\right)\text{d}s\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle \langle \unicode[STIX]{x1D748}\rangle _{b} & = & \displaystyle n\displaystyle \int \,\text{d}\boldsymbol{p}~\unicode[STIX]{x1D6FA}(\boldsymbol{p})\unicode[STIX]{x1D64E}(\boldsymbol{p})+3n\text{k}T\displaystyle \int \left(\boldsymbol{p}\boldsymbol{p}-\frac{1}{3}\unicode[STIX]{x1D644}\right)\text{d}s\end{eqnarray}$$
 $$\begin{eqnarray}\displaystyle & = & \displaystyle n\displaystyle \int \,\text{d}\boldsymbol{p}\unicode[STIX]{x1D6FA}(\boldsymbol{p})\displaystyle \int _{-L/2}^{L/2}s\left[\boldsymbol{p}\boldsymbol{f}(s)+\boldsymbol{f}(s)\boldsymbol{p}-\frac{2}{3}(\boldsymbol{f}(s)\boldsymbol{\cdot }\boldsymbol{p})\unicode[STIX]{x1D644}\right]\text{d}s\nonumber\\ \displaystyle & & \displaystyle +\,3n\text{k}T\displaystyle \int \unicode[STIX]{x1D6FA}(\boldsymbol{p})\left(\boldsymbol{p}\boldsymbol{p}-\frac{1}{3}\unicode[STIX]{x1D644}\right)\text{d}s.\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & = & \displaystyle n\displaystyle \int \,\text{d}\boldsymbol{p}\unicode[STIX]{x1D6FA}(\boldsymbol{p})\displaystyle \int _{-L/2}^{L/2}s\left[\boldsymbol{p}\boldsymbol{f}(s)+\boldsymbol{f}(s)\boldsymbol{p}-\frac{2}{3}(\boldsymbol{f}(s)\boldsymbol{\cdot }\boldsymbol{p})\unicode[STIX]{x1D644}\right]\text{d}s\nonumber\\ \displaystyle & & \displaystyle +\,3n\text{k}T\displaystyle \int \unicode[STIX]{x1D6FA}(\boldsymbol{p})\left(\boldsymbol{p}\boldsymbol{p}-\frac{1}{3}\unicode[STIX]{x1D644}\right)\text{d}s.\end{eqnarray}$$
The stress in (2.5) is specialized to slender bacteria of length
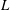 $L$
(a reasonable approximation for E. coli) in the dilute non-interacting regime, and consists of two distinct contributions. The first term in (2.5) is the hydrodynamic stresslet for a given bacterium orientation,
$L$
(a reasonable approximation for E. coli) in the dilute non-interacting regime, and consists of two distinct contributions. The first term in (2.5) is the hydrodynamic stresslet for a given bacterium orientation,
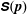 $\unicode[STIX]{x1D64E}(\boldsymbol{p})$
, averaged over the single bacterium orientation distribution
$\unicode[STIX]{x1D64E}(\boldsymbol{p})$
, averaged over the single bacterium orientation distribution
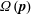 $\unicode[STIX]{x1D6FA}(\boldsymbol{p})$
as governed by (2.1). The linear force density
$\unicode[STIX]{x1D6FA}(\boldsymbol{p})$
as governed by (2.1). The linear force density
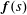 $\boldsymbol{f}(s)$
in this term is given by
$\boldsymbol{f}(s)$
in this term is given by
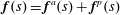 $\boldsymbol{f}(s)=\boldsymbol{f}^{a}(s)+\boldsymbol{f}^{p}(s)$
, where
$\boldsymbol{f}(s)=\boldsymbol{f}^{a}(s)+\boldsymbol{f}^{p}(s)$
, where
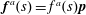 $\boldsymbol{f}^{a}(s)=f^{a}(s)\boldsymbol{p}$
is the active contribution due to the intrinsic force dipole, with
$\boldsymbol{f}^{a}(s)=f^{a}(s)\boldsymbol{p}$
is the active contribution due to the intrinsic force dipole, with
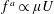 $f^{a}\propto \unicode[STIX]{x1D707}U$
, and is dependent on the particular swimming mechanism; for pushers,
$f^{a}\propto \unicode[STIX]{x1D707}U$
, and is dependent on the particular swimming mechanism; for pushers,
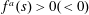 $f^{a}(s)>0({<}0)$
in the head (tail) section. The functional dependence on the axial coordinate
$f^{a}(s)>0({<}0)$
in the head (tail) section. The functional dependence on the axial coordinate
 $s$
for E. coli is specified in § 2.4. The passive component,
$s$
for E. coli is specified in § 2.4. The passive component,
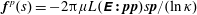 $\boldsymbol{f}^{p}(s)=-2\unicode[STIX]{x03C0}\unicode[STIX]{x1D707}L(\unicode[STIX]{x1D640}\boldsymbol{ : }\boldsymbol{p}\boldsymbol{p})s\boldsymbol{p}/(\ln \unicode[STIX]{x1D705})$
, is known from viscous slender body theory (Batchelor Reference Batchelor1970a
),
$\boldsymbol{f}^{p}(s)=-2\unicode[STIX]{x03C0}\unicode[STIX]{x1D707}L(\unicode[STIX]{x1D640}\boldsymbol{ : }\boldsymbol{p}\boldsymbol{p})s\boldsymbol{p}/(\ln \unicode[STIX]{x1D705})$
, is known from viscous slender body theory (Batchelor Reference Batchelor1970a
),
 $\unicode[STIX]{x1D705}$
being the bacterium aspect ratio.
$\unicode[STIX]{x1D705}$
being the bacterium aspect ratio.
The second term in (2.5) is the direct contribution due to stochastic reorientations (as opposed to their indirect contribution to the first term in (2.5) via
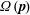 $\unicode[STIX]{x1D6FA}(\boldsymbol{p})$
). This stochastic stress, as written, is relevant to passive suspensions, where it is referred to as the diffusion or entropic stress, being proportional to the temperature
$\unicode[STIX]{x1D6FA}(\boldsymbol{p})$
). This stochastic stress, as written, is relevant to passive suspensions, where it is referred to as the diffusion or entropic stress, being proportional to the temperature
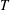 $T$
. This contribution was first identified by Kirkwood and co-workers in the context of solutions of rigid-rod macromolecules (Kirkwood & Auer Reference Kirkwood and Auer1951), and arises due to the reorienting effects of stochastic Brownian couples which relax the orientation distribution towards isotropy (Leal & Hinch Reference Leal and Hinch1971). For the active case, the stochastic changes in orientation arise in a continuous manner during a run (modelled as a rotary diffusion process in (2.1)), and impulsively during the instantaneous tumbling events. Since bacteria like E. coli are large enough to be approximated as non-Brownian (Koch & Subramanian Reference Koch and Subramanian2011), intrinsic reorientations leading to both tumbling and rotary diffusion are athermal in origin (arising from random changes in the internal configuration). Unlike the passive case, such reorientations must therefore be torque-free. This crucial difference implies that the stochastic stress contribution in the active case cannot merely be obtained from its passive analogue in (2.5) by replacing
$T$
. This contribution was first identified by Kirkwood and co-workers in the context of solutions of rigid-rod macromolecules (Kirkwood & Auer Reference Kirkwood and Auer1951), and arises due to the reorienting effects of stochastic Brownian couples which relax the orientation distribution towards isotropy (Leal & Hinch Reference Leal and Hinch1971). For the active case, the stochastic changes in orientation arise in a continuous manner during a run (modelled as a rotary diffusion process in (2.1)), and impulsively during the instantaneous tumbling events. Since bacteria like E. coli are large enough to be approximated as non-Brownian (Koch & Subramanian Reference Koch and Subramanian2011), intrinsic reorientations leading to both tumbling and rotary diffusion are athermal in origin (arising from random changes in the internal configuration). Unlike the passive case, such reorientations must therefore be torque-free. This crucial difference implies that the stochastic stress contribution in the active case cannot merely be obtained from its passive analogue in (2.5) by replacing
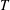 $T$
with an effective temperature related to the appropriate dissipative coefficient (
$T$
with an effective temperature related to the appropriate dissipative coefficient (
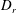 $D_{r}$
and
$D_{r}$
and
 $\unicode[STIX]{x1D70F}$
in (2.1)). The velocity disturbance due to a torque-free reorientation being weaker, an energy-dissipation argument along the lines of Einstein (1906) leads to the conclusion that the active stochastic stress must be smaller in magnitude than its passive counterpart for the same effective temperature. Hence, in contrast to earlier rheological calculations (Haines et al.
Reference Haines, Sokolov, Aranson, Berlyand and Karpeev2009; Saintillan Reference Saintillan2010), we neglect the direct stress contribution due to stochastic reorientations, and the averaged bacterial stress is taken to be:
$\unicode[STIX]{x1D70F}$
in (2.1)). The velocity disturbance due to a torque-free reorientation being weaker, an energy-dissipation argument along the lines of Einstein (1906) leads to the conclusion that the active stochastic stress must be smaller in magnitude than its passive counterpart for the same effective temperature. Hence, in contrast to earlier rheological calculations (Haines et al.
Reference Haines, Sokolov, Aranson, Berlyand and Karpeev2009; Saintillan Reference Saintillan2010), we neglect the direct stress contribution due to stochastic reorientations, and the averaged bacterial stress is taken to be:
 $$\begin{eqnarray}\langle \unicode[STIX]{x1D748}\rangle _{b}=n\displaystyle \int \,\text{d}\boldsymbol{p}~\unicode[STIX]{x1D6FA}(\boldsymbol{p})\displaystyle \int _{-L/2}^{L/2}s\left[\boldsymbol{p}\boldsymbol{f}(s)+\boldsymbol{f}(s)\boldsymbol{p}-\frac{2}{3}(\boldsymbol{f}(s)\boldsymbol{\cdot }\boldsymbol{p})\unicode[STIX]{x1D644}\right]\text{d}s,\end{eqnarray}$$
$$\begin{eqnarray}\langle \unicode[STIX]{x1D748}\rangle _{b}=n\displaystyle \int \,\text{d}\boldsymbol{p}~\unicode[STIX]{x1D6FA}(\boldsymbol{p})\displaystyle \int _{-L/2}^{L/2}s\left[\boldsymbol{p}\boldsymbol{f}(s)+\boldsymbol{f}(s)\boldsymbol{p}-\frac{2}{3}(\boldsymbol{f}(s)\boldsymbol{\cdot }\boldsymbol{p})\unicode[STIX]{x1D644}\right]\text{d}s,\end{eqnarray}$$
with rotary diffusion and tumbling only contributing via their influence on
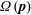 $\unicode[STIX]{x1D6FA}(\boldsymbol{p})$
.
$\unicode[STIX]{x1D6FA}(\boldsymbol{p})$
.
A calculation of the active stochastic stress from first principles would depend sensitively on the swimming mechanism (ciliary as opposed to flagellar propulsion, for instance), and is beyond the scope of this paper. Nevertheless, in appendix A, we estimate the stress that arises due to the combined effects of rotary diffusion and run-and-tumble dynamics, in the same manner as that for passive suspensions, by using an orientation-space-flux-based approach. As argued above, this gives an upper bound for the magnitude of the active stochastic stress. We find this contribution to be less than
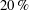 $20\,\%$
of the net stress, thereby justifying its omission.
$20\,\%$
of the net stress, thereby justifying its omission.
2.3 The solution for the orientation probability density
Choosing
 $\unicode[STIX]{x1D70F}$
as the relevant time scale, (2.1) may be non-dimensionalized as:
$\unicode[STIX]{x1D70F}$
as the relevant time scale, (2.1) may be non-dimensionalized as:
 $$\begin{eqnarray}\frac{\unicode[STIX]{x2202}\unicode[STIX]{x1D6FA}}{\unicode[STIX]{x2202}t}+\left(\unicode[STIX]{x1D6FA}-\int K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })\unicode[STIX]{x1D6FA}(\boldsymbol{p}^{\prime },t)\,\text{d}\boldsymbol{p}^{\prime }\right)-(\unicode[STIX]{x1D70F}D_{r})\unicode[STIX]{x1D6FB}_{\boldsymbol{p}}^{2}\unicode[STIX]{x1D6FA}=-Pe\unicode[STIX]{x1D735}_{\boldsymbol{ p}}\boldsymbol{\cdot }(\dot{\boldsymbol{p}}\unicode[STIX]{x1D6FA}),\end{eqnarray}$$
$$\begin{eqnarray}\frac{\unicode[STIX]{x2202}\unicode[STIX]{x1D6FA}}{\unicode[STIX]{x2202}t}+\left(\unicode[STIX]{x1D6FA}-\int K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })\unicode[STIX]{x1D6FA}(\boldsymbol{p}^{\prime },t)\,\text{d}\boldsymbol{p}^{\prime }\right)-(\unicode[STIX]{x1D70F}D_{r})\unicode[STIX]{x1D6FB}_{\boldsymbol{p}}^{2}\unicode[STIX]{x1D6FA}=-Pe\unicode[STIX]{x1D735}_{\boldsymbol{ p}}\boldsymbol{\cdot }(\dot{\boldsymbol{p}}\unicode[STIX]{x1D6FA}),\end{eqnarray}$$
where we retain the same notation for simplicity. In (2.7),
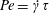 $Pe=\dot{\unicode[STIX]{x1D6FE}}\unicode[STIX]{x1D70F}$
is a Péclet number based on
$Pe=\dot{\unicode[STIX]{x1D6FE}}\unicode[STIX]{x1D70F}$
is a Péclet number based on
 $\unicode[STIX]{x1D70F}$
, run-and-tumble dynamics being the primary means of relaxation and
$\unicode[STIX]{x1D70F}$
, run-and-tumble dynamics being the primary means of relaxation and
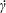 $\dot{\unicode[STIX]{x1D6FE}}$
is a characteristic shear rate amplitude. The parameter
$\dot{\unicode[STIX]{x1D6FE}}$
is a characteristic shear rate amplitude. The parameter
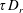 $\unicode[STIX]{x1D70F}D_{r}$
denotes the relative efficiency of the two relaxation processes, and for the wild-type bacteria in question,
$\unicode[STIX]{x1D70F}D_{r}$
denotes the relative efficiency of the two relaxation processes, and for the wild-type bacteria in question,
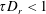 $\unicode[STIX]{x1D70F}D_{r}<1$
. For the small shear rates relevant to the linear response regime,
$\unicode[STIX]{x1D70F}D_{r}<1$
. For the small shear rates relevant to the linear response regime,
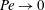 $Pe\rightarrow 0$
, and the probability density may be expanded about an isotropic base state as
$Pe\rightarrow 0$
, and the probability density may be expanded about an isotropic base state as
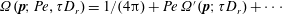 $\unicode[STIX]{x1D6FA}(\boldsymbol{p};Pe,\unicode[STIX]{x1D70F}D_{r})=1/(4\unicode[STIX]{x03C0})+Pe\,\unicode[STIX]{x1D6FA}^{\prime }(\boldsymbol{p};\unicode[STIX]{x1D70F}D_{r})+\cdots \,$
, with
$\unicode[STIX]{x1D6FA}(\boldsymbol{p};Pe,\unicode[STIX]{x1D70F}D_{r})=1/(4\unicode[STIX]{x03C0})+Pe\,\unicode[STIX]{x1D6FA}^{\prime }(\boldsymbol{p};\unicode[STIX]{x1D70F}D_{r})+\cdots \,$
, with
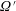 $\unicode[STIX]{x1D6FA}^{\prime }$
satisfying
$\unicode[STIX]{x1D6FA}^{\prime }$
satisfying
 $$\begin{eqnarray}\frac{\unicode[STIX]{x2202}\unicode[STIX]{x1D6FA}^{\prime }}{\unicode[STIX]{x2202}t}+\Big(\unicode[STIX]{x1D6FA}^{\prime }-\int K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })\unicode[STIX]{x1D6FA}^{\prime }(\boldsymbol{p}^{\prime },t)\,\text{d}\boldsymbol{p}^{\prime }\Big)-(\unicode[STIX]{x1D70F}D_{r})\unicode[STIX]{x1D6FB}_{\boldsymbol{p}}^{2}\unicode[STIX]{x1D6FA}^{\prime }=\frac{3}{4\unicode[STIX]{x03C0}}\unicode[STIX]{x1D640}(t)\boldsymbol{ : }\boldsymbol{p}\boldsymbol{p},\end{eqnarray}$$
$$\begin{eqnarray}\frac{\unicode[STIX]{x2202}\unicode[STIX]{x1D6FA}^{\prime }}{\unicode[STIX]{x2202}t}+\Big(\unicode[STIX]{x1D6FA}^{\prime }-\int K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })\unicode[STIX]{x1D6FA}^{\prime }(\boldsymbol{p}^{\prime },t)\,\text{d}\boldsymbol{p}^{\prime }\Big)-(\unicode[STIX]{x1D70F}D_{r})\unicode[STIX]{x1D6FB}_{\boldsymbol{p}}^{2}\unicode[STIX]{x1D6FA}^{\prime }=\frac{3}{4\unicode[STIX]{x03C0}}\unicode[STIX]{x1D640}(t)\boldsymbol{ : }\boldsymbol{p}\boldsymbol{p},\end{eqnarray}$$
at leading order where we have used
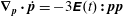 $\unicode[STIX]{x1D735}_{\boldsymbol{p}}\boldsymbol{\cdot }\dot{\boldsymbol{p}}=-3\unicode[STIX]{x1D640}(t)\boldsymbol{ : }\boldsymbol{p}\boldsymbol{p}$
for
$\unicode[STIX]{x1D735}_{\boldsymbol{p}}\boldsymbol{\cdot }\dot{\boldsymbol{p}}=-3\unicode[STIX]{x1D640}(t)\boldsymbol{ : }\boldsymbol{p}\boldsymbol{p}$
for
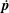 $\dot{\boldsymbol{p}}$
given by (2.2); only the rate of strain associated with the ambient linear flow acts to distort the isotropic distribution at leading order. We now solve the above inhomogeneous integro-differential equation in terms of the Green’s function,
$\dot{\boldsymbol{p}}$
given by (2.2); only the rate of strain associated with the ambient linear flow acts to distort the isotropic distribution at leading order. We now solve the above inhomogeneous integro-differential equation in terms of the Green’s function,
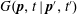 $G(\boldsymbol{p},t\mid \boldsymbol{p}^{\prime },t^{\prime })$
, which satisfies:
$G(\boldsymbol{p},t\mid \boldsymbol{p}^{\prime },t^{\prime })$
, which satisfies:
 $$\begin{eqnarray}\frac{\unicode[STIX]{x2202}G}{\unicode[STIX]{x2202}t}+\left(G-\int K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime \prime })G(\boldsymbol{p}^{\prime \prime },t\mid \mathbf{p}^{\prime },t^{\prime })\,\text{d}\boldsymbol{p}^{\prime \prime }\right)-(\unicode[STIX]{x1D70F}D_{r})\unicode[STIX]{x1D6FB}_{\boldsymbol{p}}^{2}G=\unicode[STIX]{x1D739}(\boldsymbol{p}-\boldsymbol{p}^{\prime })\unicode[STIX]{x1D739}(t-t^{\prime }),\end{eqnarray}$$
$$\begin{eqnarray}\frac{\unicode[STIX]{x2202}G}{\unicode[STIX]{x2202}t}+\left(G-\int K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime \prime })G(\boldsymbol{p}^{\prime \prime },t\mid \mathbf{p}^{\prime },t^{\prime })\,\text{d}\boldsymbol{p}^{\prime \prime }\right)-(\unicode[STIX]{x1D70F}D_{r})\unicode[STIX]{x1D6FB}_{\boldsymbol{p}}^{2}G=\unicode[STIX]{x1D739}(\boldsymbol{p}-\boldsymbol{p}^{\prime })\unicode[STIX]{x1D739}(t-t^{\prime }),\end{eqnarray}$$
where
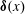 $\unicode[STIX]{x1D739}(x)$
is the Dirac-delta function. We expand
$\unicode[STIX]{x1D739}(x)$
is the Dirac-delta function. We expand
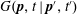 $G(\boldsymbol{p},t\mid \boldsymbol{p}^{\prime },t^{\prime })$
as a series in surface spherical harmonics:
$G(\boldsymbol{p},t\mid \boldsymbol{p}^{\prime },t^{\prime })$
as a series in surface spherical harmonics:
 $$\begin{eqnarray}G(\boldsymbol{p},t\mid \boldsymbol{p}^{\prime },t^{\prime })=\mathop{\sum }_{n=0}^{\infty }\mathop{\sum }_{m=-n}^{n}R_{n,m}(t-t^{\prime })C_{n,m}(\boldsymbol{p}^{\prime })Y_{n}^{m}(\boldsymbol{p}),\end{eqnarray}$$
$$\begin{eqnarray}G(\boldsymbol{p},t\mid \boldsymbol{p}^{\prime },t^{\prime })=\mathop{\sum }_{n=0}^{\infty }\mathop{\sum }_{m=-n}^{n}R_{n,m}(t-t^{\prime })C_{n,m}(\boldsymbol{p}^{\prime })Y_{n}^{m}(\boldsymbol{p}),\end{eqnarray}$$
where
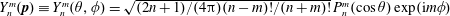 $Y_{n}^{m}(\boldsymbol{p})\equiv Y_{n}^{m}(\unicode[STIX]{x1D703},\unicode[STIX]{x1D719})=\sqrt{(2n+1)/(4\unicode[STIX]{x03C0})(n-m)!/(n+m)!}\,P_{n}^{m}(\cos \unicode[STIX]{x1D703})\exp (\text{i}m\unicode[STIX]{x1D719})$
, with
$Y_{n}^{m}(\boldsymbol{p})\equiv Y_{n}^{m}(\unicode[STIX]{x1D703},\unicode[STIX]{x1D719})=\sqrt{(2n+1)/(4\unicode[STIX]{x03C0})(n-m)!/(n+m)!}\,P_{n}^{m}(\cos \unicode[STIX]{x1D703})\exp (\text{i}m\unicode[STIX]{x1D719})$
, with
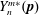 $Y_{n}^{m\ast }(\boldsymbol{p})$
denoting its complex conjugate (Abramowitz & Stegun Reference Abramowitz and Stegun1970); for the specific case of simple shear flow considered below,
$Y_{n}^{m\ast }(\boldsymbol{p})$
denoting its complex conjugate (Abramowitz & Stegun Reference Abramowitz and Stegun1970); for the specific case of simple shear flow considered below,
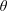 $\unicode[STIX]{x1D703}$
is the polar angle measured from the vorticity axis and
$\unicode[STIX]{x1D703}$
is the polar angle measured from the vorticity axis and
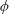 $\unicode[STIX]{x1D719}$
is the azimuthal angle measured relative to the flow direction. On using the relations:
$\unicode[STIX]{x1D719}$
is the azimuthal angle measured relative to the flow direction. On using the relations:
 $$\begin{eqnarray}\displaystyle & \displaystyle \unicode[STIX]{x1D739}(\boldsymbol{p}-\boldsymbol{p}^{\prime })=\mathop{\sum }_{n=0}^{\infty }\mathop{\sum }_{m=-n}^{n}Y_{n}^{m}(\boldsymbol{p})Y_{n}^{m\ast }(\boldsymbol{p}^{\prime }), & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & \displaystyle \unicode[STIX]{x1D739}(\boldsymbol{p}-\boldsymbol{p}^{\prime })=\mathop{\sum }_{n=0}^{\infty }\mathop{\sum }_{m=-n}^{n}Y_{n}^{m}(\boldsymbol{p})Y_{n}^{m\ast }(\boldsymbol{p}^{\prime }), & \displaystyle\end{eqnarray}$$
 $$\begin{eqnarray}\displaystyle & \displaystyle K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })=\mathop{\sum }_{n=0}^{\infty }A_{n}P_{n}(\boldsymbol{p}\boldsymbol{\cdot }\boldsymbol{p}^{\prime })=\mathop{\sum }_{n=0}^{\infty }\mathop{\sum }_{m=-n}^{n}\frac{A_{n}4\unicode[STIX]{x03C0}(-1)^{m}}{(2n+1)}Y_{n}^{m}(\boldsymbol{p})Y_{n}^{-m}(\boldsymbol{p}^{\prime }), & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & \displaystyle K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })=\mathop{\sum }_{n=0}^{\infty }A_{n}P_{n}(\boldsymbol{p}\boldsymbol{\cdot }\boldsymbol{p}^{\prime })=\mathop{\sum }_{n=0}^{\infty }\mathop{\sum }_{m=-n}^{n}\frac{A_{n}4\unicode[STIX]{x03C0}(-1)^{m}}{(2n+1)}Y_{n}^{m}(\boldsymbol{p})Y_{n}^{-m}(\boldsymbol{p}^{\prime }), & \displaystyle\end{eqnarray}$$
where (2.11) is the spectral expansion of the delta function and the addition theorem for spherical harmonics has been used to arrive at (2.12), with
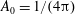 $A_{0}=1/(4\unicode[STIX]{x03C0})$
and a non-zero
$A_{0}=1/(4\unicode[STIX]{x03C0})$
and a non-zero
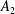 $A_{2}$
denoting correlation between pre- and post-tumble orientations, one obtains from (2.9):
$A_{2}$
denoting correlation between pre- and post-tumble orientations, one obtains from (2.9):
 $$\begin{eqnarray}\displaystyle & & \displaystyle \mathop{\sum }_{n=0}^{\infty }\mathop{\sum }_{m=-n}^{n}\left[\vphantom{\mathop{\sum }_{q=-p}^{p}}\left(\frac{\text{d}R_{n,m}}{\text{d}t}+R_{n,m}\right)C_{n,m}(\boldsymbol{p}^{\prime })Y_{n}^{m}(\boldsymbol{p})+(\unicode[STIX]{x1D70F}D_{r})R_{n,m}C_{n,m}(\boldsymbol{p}^{\prime })\unicode[STIX]{x1D6FB}_{\boldsymbol{p}}^{2}Y_{n}^{m}(\boldsymbol{p})\right.\nonumber\\ \displaystyle & & \displaystyle \qquad \left.-\,R_{n,m}\int \mathop{\sum }_{p=0}^{\infty }\mathop{\sum }_{q=-p}^{p}\frac{A_{p}4\unicode[STIX]{x03C0}(-1)^{q}}{(2p+1)}C_{n,m}(\boldsymbol{p}^{\prime })Y_{p}^{q}(\boldsymbol{p})Y_{p}^{-q}(\boldsymbol{p}^{\prime \prime })Y_{n}^{m}(\boldsymbol{p}^{\prime \prime })\,\text{d}\boldsymbol{p}^{\prime \prime }\right]\nonumber\\ \displaystyle & & \displaystyle \quad =\unicode[STIX]{x1D739}(t-t^{\prime })\mathop{\sum }_{n=0}^{\infty }\mathop{\sum }_{m=-n}^{n}Y_{n}^{m}(\boldsymbol{p})Y_{n}^{m\ast }(\boldsymbol{p}^{\prime }).\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & & \displaystyle \mathop{\sum }_{n=0}^{\infty }\mathop{\sum }_{m=-n}^{n}\left[\vphantom{\mathop{\sum }_{q=-p}^{p}}\left(\frac{\text{d}R_{n,m}}{\text{d}t}+R_{n,m}\right)C_{n,m}(\boldsymbol{p}^{\prime })Y_{n}^{m}(\boldsymbol{p})+(\unicode[STIX]{x1D70F}D_{r})R_{n,m}C_{n,m}(\boldsymbol{p}^{\prime })\unicode[STIX]{x1D6FB}_{\boldsymbol{p}}^{2}Y_{n}^{m}(\boldsymbol{p})\right.\nonumber\\ \displaystyle & & \displaystyle \qquad \left.-\,R_{n,m}\int \mathop{\sum }_{p=0}^{\infty }\mathop{\sum }_{q=-p}^{p}\frac{A_{p}4\unicode[STIX]{x03C0}(-1)^{q}}{(2p+1)}C_{n,m}(\boldsymbol{p}^{\prime })Y_{p}^{q}(\boldsymbol{p})Y_{p}^{-q}(\boldsymbol{p}^{\prime \prime })Y_{n}^{m}(\boldsymbol{p}^{\prime \prime })\,\text{d}\boldsymbol{p}^{\prime \prime }\right]\nonumber\\ \displaystyle & & \displaystyle \quad =\unicode[STIX]{x1D739}(t-t^{\prime })\mathop{\sum }_{n=0}^{\infty }\mathop{\sum }_{m=-n}^{n}Y_{n}^{m}(\boldsymbol{p})Y_{n}^{m\ast }(\boldsymbol{p}^{\prime }).\end{eqnarray}$$
Further, on using
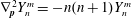 $\unicode[STIX]{x1D6FB}_{\boldsymbol{p}}^{2}Y_{n}^{m}=-n(n+1)Y_{n}^{m}$
, (2.13) simplifies to:
$\unicode[STIX]{x1D6FB}_{\boldsymbol{p}}^{2}Y_{n}^{m}=-n(n+1)Y_{n}^{m}$
, (2.13) simplifies to:
 $$\begin{eqnarray}\displaystyle & & \displaystyle \mathop{\sum }_{n=0}^{\infty }\mathop{\sum }_{m=-n}^{n}\left[\left(\frac{\text{d}R_{n,m}}{\text{d}t}+R_{n,m}+n(n+1)(\unicode[STIX]{x1D70F}D_{r})R_{n,m}\right)-\frac{A_{n}4\unicode[STIX]{x03C0}}{(2n+1)}R_{n,m}\right]C_{n,m}(\boldsymbol{p}^{\prime })Y_{n}^{m}(\boldsymbol{p})\nonumber\\ \displaystyle & & \displaystyle \quad =\unicode[STIX]{x1D739}(t-t^{\prime })\mathop{\sum }_{n=0}^{\infty }\mathop{\sum }_{m=-n}^{n}Y_{n}^{m}(\boldsymbol{p})Y_{n}^{m\ast }(\boldsymbol{p}^{\prime }),\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & & \displaystyle \mathop{\sum }_{n=0}^{\infty }\mathop{\sum }_{m=-n}^{n}\left[\left(\frac{\text{d}R_{n,m}}{\text{d}t}+R_{n,m}+n(n+1)(\unicode[STIX]{x1D70F}D_{r})R_{n,m}\right)-\frac{A_{n}4\unicode[STIX]{x03C0}}{(2n+1)}R_{n,m}\right]C_{n,m}(\boldsymbol{p}^{\prime })Y_{n}^{m}(\boldsymbol{p})\nonumber\\ \displaystyle & & \displaystyle \quad =\unicode[STIX]{x1D739}(t-t^{\prime })\mathop{\sum }_{n=0}^{\infty }\mathop{\sum }_{m=-n}^{n}Y_{n}^{m}(\boldsymbol{p})Y_{n}^{m\ast }(\boldsymbol{p}^{\prime }),\end{eqnarray}$$
on using the orthogonality of the surface spherical harmonics. Now, comparing the coefficients of
 $Y_{n}^{m}(\boldsymbol{p})$
on both sides of the equation, one obtains:
$Y_{n}^{m}(\boldsymbol{p})$
on both sides of the equation, one obtains:
 $$\begin{eqnarray}\displaystyle & \displaystyle C_{n,m}(\boldsymbol{p}^{\prime })=Y_{n}^{m\ast }(\boldsymbol{p}^{\prime }), & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & \displaystyle C_{n,m}(\boldsymbol{p}^{\prime })=Y_{n}^{m\ast }(\boldsymbol{p}^{\prime }), & \displaystyle\end{eqnarray}$$
 $$\begin{eqnarray}\displaystyle & \displaystyle R_{m,n}\equiv R_{n}=\exp \left[-\left\{(1-\frac{4\unicode[STIX]{x03C0}A_{n}}{(2n+1)})+n(n+1)(\unicode[STIX]{x1D70F}D_{r})\right\}(t-t^{\prime })\right]H(t-t^{\prime }), & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & \displaystyle R_{m,n}\equiv R_{n}=\exp \left[-\left\{(1-\frac{4\unicode[STIX]{x03C0}A_{n}}{(2n+1)})+n(n+1)(\unicode[STIX]{x1D70F}D_{r})\right\}(t-t^{\prime })\right]H(t-t^{\prime }), & \displaystyle\end{eqnarray}$$
where
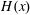 $H(x)$
is the Heaviside function. Thus, the Green’s function takes the form:
$H(x)$
is the Heaviside function. Thus, the Green’s function takes the form:
 $$\begin{eqnarray}\displaystyle \hspace{-15.00002pt}G(\boldsymbol{p},t\mid \boldsymbol{p}^{\prime },t^{\prime }) & = & \displaystyle H(t-t^{\prime })\left[\frac{1}{4\unicode[STIX]{x03C0}}+\mathop{\sum }_{n=1}^{\infty }\mathop{\sum }_{m=-n}^{n}Y_{n}^{m}(\boldsymbol{p})Y_{n}^{m\ast }(\boldsymbol{p}^{\prime })\right.\nonumber\\ \displaystyle & & \displaystyle \left.\times \,\exp \left(-\left\{\left(1-\frac{4\unicode[STIX]{x03C0}A_{n}}{(2n+1)}\right)+n(n+1)(\unicode[STIX]{x1D70F}D_{r})\right\}(t-t^{\prime })\right)\vphantom{\mathop{\sum }_{m=-n}^{n}}\right]\!.\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle \hspace{-15.00002pt}G(\boldsymbol{p},t\mid \boldsymbol{p}^{\prime },t^{\prime }) & = & \displaystyle H(t-t^{\prime })\left[\frac{1}{4\unicode[STIX]{x03C0}}+\mathop{\sum }_{n=1}^{\infty }\mathop{\sum }_{m=-n}^{n}Y_{n}^{m}(\boldsymbol{p})Y_{n}^{m\ast }(\boldsymbol{p}^{\prime })\right.\nonumber\\ \displaystyle & & \displaystyle \left.\times \,\exp \left(-\left\{\left(1-\frac{4\unicode[STIX]{x03C0}A_{n}}{(2n+1)}\right)+n(n+1)(\unicode[STIX]{x1D70F}D_{r})\right\}(t-t^{\prime })\right)\vphantom{\mathop{\sum }_{m=-n}^{n}}\right]\!.\end{eqnarray}$$
In the absence of tumbling, (2.17) takes the familiar form known for the transition probability characterizing Brownian motion on the unit sphere (Ghosh, Samuel & Sinha Reference Ghosh, Samuel and Sinha2012). With
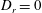 $D_{r}=0$
and for random tumbles,
$D_{r}=0$
and for random tumbles,
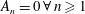 $A_{n}=0\,\forall \,n\geqslant 1$
, (2.17) takes the much simpler form
$A_{n}=0\,\forall \,n\geqslant 1$
, (2.17) takes the much simpler form
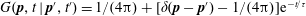 $G(\boldsymbol{p},t\mid \boldsymbol{p}^{\prime },t^{\prime })=1/(4\unicode[STIX]{x03C0})+[\unicode[STIX]{x1D6FF}(\boldsymbol{p}-\boldsymbol{p}^{\prime })-1/(4\unicode[STIX]{x03C0})]\text{e}^{-t/\unicode[STIX]{x1D70F}}$
, implying that all deviations from isotropy decay exponentially on the single time scale
$G(\boldsymbol{p},t\mid \boldsymbol{p}^{\prime },t^{\prime })=1/(4\unicode[STIX]{x03C0})+[\unicode[STIX]{x1D6FF}(\boldsymbol{p}-\boldsymbol{p}^{\prime })-1/(4\unicode[STIX]{x03C0})]\text{e}^{-t/\unicode[STIX]{x1D70F}}$
, implying that all deviations from isotropy decay exponentially on the single time scale
 $\unicode[STIX]{x1D70F}$
(Sandoval et al.
Reference Sandoval, Navaneeth, Subramanian and Lauga2014).
$\unicode[STIX]{x1D70F}$
(Sandoval et al.
Reference Sandoval, Navaneeth, Subramanian and Lauga2014).
With the Green’s function given by (2.17), the flow-induced perturbation to the orientation probability density is
 $$\begin{eqnarray}\unicode[STIX]{x1D6FA}^{\prime }(\boldsymbol{p},t)=\frac{3}{4\unicode[STIX]{x03C0}}\int _{0}^{t}\,\text{d}t^{\prime }\int G(\boldsymbol{p},t\mid \boldsymbol{p}^{\prime },t^{\prime })\unicode[STIX]{x1D640}(t^{\prime })\boldsymbol{ : }\boldsymbol{p}^{\prime }\boldsymbol{p}^{\prime }\,\text{d}\boldsymbol{p}^{\prime }.\end{eqnarray}$$
$$\begin{eqnarray}\unicode[STIX]{x1D6FA}^{\prime }(\boldsymbol{p},t)=\frac{3}{4\unicode[STIX]{x03C0}}\int _{0}^{t}\,\text{d}t^{\prime }\int G(\boldsymbol{p},t\mid \boldsymbol{p}^{\prime },t^{\prime })\unicode[STIX]{x1D640}(t^{\prime })\boldsymbol{ : }\boldsymbol{p}^{\prime }\boldsymbol{p}^{\prime }\,\text{d}\boldsymbol{p}^{\prime }.\end{eqnarray}$$
For a simple shearing flow, with
 $1$
,
$1$
,
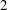 $2$
and
$2$
and
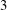 $3$
as the flow, gradient and vorticity directions, respectively,
$3$
as the flow, gradient and vorticity directions, respectively,
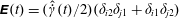 $\unicode[STIX]{x1D640}(t)=(\hat{\dot{\unicode[STIX]{x1D6FE}}}(t)/2)(\unicode[STIX]{x1D6FF}_{i2}\unicode[STIX]{x1D6FF}_{j1}+\unicode[STIX]{x1D6FF}_{i1}\unicode[STIX]{x1D6FF}_{j2})$
,
$\unicode[STIX]{x1D640}(t)=(\hat{\dot{\unicode[STIX]{x1D6FE}}}(t)/2)(\unicode[STIX]{x1D6FF}_{i2}\unicode[STIX]{x1D6FF}_{j1}+\unicode[STIX]{x1D6FF}_{i1}\unicode[STIX]{x1D6FF}_{j2})$
,
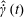 $\hat{\dot{\unicode[STIX]{x1D6FE}}}(t)$
being the dimensionless time-dependent shear rate, and
$\hat{\dot{\unicode[STIX]{x1D6FE}}}(t)$
being the dimensionless time-dependent shear rate, and
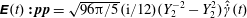 $\unicode[STIX]{x1D640}(t)\boldsymbol{ : }\boldsymbol{p}\boldsymbol{p}=\sqrt{96\unicode[STIX]{x03C0}/5}(\text{i}/12)(Y_{2}^{-2}-Y_{2}^{2})\hat{\dot{\unicode[STIX]{x1D6FE}}}(t)$
. The orthogonality of the surface spherical harmonics leads to the expression
$\unicode[STIX]{x1D640}(t)\boldsymbol{ : }\boldsymbol{p}\boldsymbol{p}=\sqrt{96\unicode[STIX]{x03C0}/5}(\text{i}/12)(Y_{2}^{-2}-Y_{2}^{2})\hat{\dot{\unicode[STIX]{x1D6FE}}}(t)$
. The orthogonality of the surface spherical harmonics leads to the expression
 $$\begin{eqnarray}\unicode[STIX]{x1D6FA}^{\prime }(\boldsymbol{p},t)=\frac{3\sin ^{2}\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719}\cos \unicode[STIX]{x1D719}}{4\unicode[STIX]{x03C0}}\int _{t^{\prime }}^{t}\exp \left[-\left\{\left(1-\frac{4\unicode[STIX]{x03C0}A_{2}}{5}\right)+6(\unicode[STIX]{x1D70F}D_{r})\right\}(t^{\prime \prime }-t^{\prime })\right]\hat{\dot{\unicode[STIX]{x1D6FE}}}(t^{\prime \prime })\,\text{d}t^{\prime \prime },\end{eqnarray}$$
$$\begin{eqnarray}\unicode[STIX]{x1D6FA}^{\prime }(\boldsymbol{p},t)=\frac{3\sin ^{2}\unicode[STIX]{x1D703}\sin \unicode[STIX]{x1D719}\cos \unicode[STIX]{x1D719}}{4\unicode[STIX]{x03C0}}\int _{t^{\prime }}^{t}\exp \left[-\left\{\left(1-\frac{4\unicode[STIX]{x03C0}A_{2}}{5}\right)+6(\unicode[STIX]{x1D70F}D_{r})\right\}(t^{\prime \prime }-t^{\prime })\right]\hat{\dot{\unicode[STIX]{x1D6FE}}}(t^{\prime \prime })\,\text{d}t^{\prime \prime },\end{eqnarray}$$
where
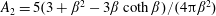 $A_{2}=5(3+\unicode[STIX]{x1D6FD}^{2}-3\unicode[STIX]{x1D6FD}\coth \unicode[STIX]{x1D6FD})/(4\unicode[STIX]{x03C0}\unicode[STIX]{x1D6FD}^{2})$
captures the effect of tumble correlations. For times long compared to
$A_{2}=5(3+\unicode[STIX]{x1D6FD}^{2}-3\unicode[STIX]{x1D6FD}\coth \unicode[STIX]{x1D6FD})/(4\unicode[STIX]{x03C0}\unicode[STIX]{x1D6FD}^{2})$
captures the effect of tumble correlations. For times long compared to
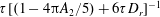 $\unicode[STIX]{x1D70F}[(1-4\unicode[STIX]{x03C0}A_{2}/5)+6\unicode[STIX]{x1D70F}D_{r}]^{-1}$
, the orientation anisotropy predicted by (2.19) peaks along the ambient extensional axis. This anisotropy contributes to the zero-shear viscosity which is evaluated and compared to experiments below. As is known from the microstructural response of passive fluids, at higher orders in the shear rate, the peak in the orientation anisotropy generally tilts towards the flow axis (Brenner Reference Brenner1974); this leads to normal stress differences at quadratic order in the shear rate, a shear thinning contribution to the viscosity at cubic order and so on (Bird, Armstrong & Hassager Reference Bird, Armstrong and Hassager1987).
$\unicode[STIX]{x1D70F}[(1-4\unicode[STIX]{x03C0}A_{2}/5)+6\unicode[STIX]{x1D70F}D_{r}]^{-1}$
, the orientation anisotropy predicted by (2.19) peaks along the ambient extensional axis. This anisotropy contributes to the zero-shear viscosity which is evaluated and compared to experiments below. As is known from the microstructural response of passive fluids, at higher orders in the shear rate, the peak in the orientation anisotropy generally tilts towards the flow axis (Brenner Reference Brenner1974); this leads to normal stress differences at quadratic order in the shear rate, a shear thinning contribution to the viscosity at cubic order and so on (Bird, Armstrong & Hassager Reference Bird, Armstrong and Hassager1987).
2.4 The time-dependent shear stress response
The deviatoric bacterial stress, relevant to a rheological response, may now be obtained at leading order using (2.6). With
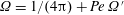 $\unicode[STIX]{x1D6FA}=1/(4\unicode[STIX]{x03C0})+Pe\,\unicode[STIX]{x1D6FA}^{\prime }$
, and
$\unicode[STIX]{x1D6FA}=1/(4\unicode[STIX]{x03C0})+Pe\,\unicode[STIX]{x1D6FA}^{\prime }$
, and
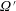 $\unicode[STIX]{x1D6FA}^{\prime }$
given by (2.19), one obtains:
$\unicode[STIX]{x1D6FA}^{\prime }$
given by (2.19), one obtains:
 $$\begin{eqnarray}\displaystyle \langle \unicode[STIX]{x1D748}\rangle _{b} & = & \displaystyle \frac{(\unicode[STIX]{x1D707}\dot{\unicode[STIX]{x1D6FE}})(nL^{3})}{24\ln \unicode[STIX]{x1D705}}\int \,\text{d}\boldsymbol{p}(\unicode[STIX]{x1D640}(t)\boldsymbol{ : }\boldsymbol{p}\boldsymbol{p})\left(\boldsymbol{p}\boldsymbol{p}-\frac{1}{3}\unicode[STIX]{x1D644}\right)\nonumber\\ \displaystyle & & \displaystyle -\,Pe\unicode[STIX]{x1D707}(nUL^{2})\int _{-1/2}^{1/2}s\hat{f}^{a}(s)\,\text{d}s\int \,\text{d}\boldsymbol{p}\,\unicode[STIX]{x1D6FA}^{\prime }(\boldsymbol{p},t)\left(\boldsymbol{p}\boldsymbol{p}-\frac{1}{3}\unicode[STIX]{x1D644}\right),\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle \langle \unicode[STIX]{x1D748}\rangle _{b} & = & \displaystyle \frac{(\unicode[STIX]{x1D707}\dot{\unicode[STIX]{x1D6FE}})(nL^{3})}{24\ln \unicode[STIX]{x1D705}}\int \,\text{d}\boldsymbol{p}(\unicode[STIX]{x1D640}(t)\boldsymbol{ : }\boldsymbol{p}\boldsymbol{p})\left(\boldsymbol{p}\boldsymbol{p}-\frac{1}{3}\unicode[STIX]{x1D644}\right)\nonumber\\ \displaystyle & & \displaystyle -\,Pe\unicode[STIX]{x1D707}(nUL^{2})\int _{-1/2}^{1/2}s\hat{f}^{a}(s)\,\text{d}s\int \,\text{d}\boldsymbol{p}\,\unicode[STIX]{x1D6FA}^{\prime }(\boldsymbol{p},t)\left(\boldsymbol{p}\boldsymbol{p}-\frac{1}{3}\unicode[STIX]{x1D644}\right),\end{eqnarray}$$
where
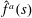 $\hat{f}^{a}(s)$
denotes the non-dimensional active force density. Here, we have chosen the total bacterium length
$\hat{f}^{a}(s)$
denotes the non-dimensional active force density. Here, we have chosen the total bacterium length
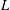 $L$
as a characteristic length scale, so that
$L$
as a characteristic length scale, so that
 $s$
in (2.20), and in what follows, is a non-dimensional axial coordinate. Note that the passive stress response in (2.20) arises from an isotropic orientation distribution at linear order, and therefore, has an instantaneous viscous character similar to that of the solvent. Thus, the non-trivial time dependence, for small shear rates, arises entirely from the time-dependent anisotropy of the intrinsic force dipoles. We take
$s$
in (2.20), and in what follows, is a non-dimensional axial coordinate. Note that the passive stress response in (2.20) arises from an isotropic orientation distribution at linear order, and therefore, has an instantaneous viscous character similar to that of the solvent. Thus, the non-trivial time dependence, for small shear rates, arises entirely from the time-dependent anisotropy of the intrinsic force dipoles. We take
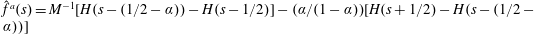 $\hat{f}^{a}(s)=M^{-1}[H(s-(1/2-\unicode[STIX]{x1D6FC}))-H(s-1/2)]-(\unicode[STIX]{x1D6FC}/(1-\unicode[STIX]{x1D6FC}))[H(s+1/2)-H(s-(1/2-\unicode[STIX]{x1D6FC}))]$
where
$\hat{f}^{a}(s)=M^{-1}[H(s-(1/2-\unicode[STIX]{x1D6FC}))-H(s-1/2)]-(\unicode[STIX]{x1D6FC}/(1-\unicode[STIX]{x1D6FC}))[H(s+1/2)-H(s-(1/2-\unicode[STIX]{x1D6FC}))]$
where
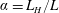 $\unicode[STIX]{x1D6FC}=L_{H}/L$
is the ratio of the head length (
$\unicode[STIX]{x1D6FC}=L_{H}/L$
is the ratio of the head length (
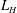 $L_{H}$
) to the total bacterium length and
$L_{H}$
) to the total bacterium length and
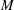 $M$
is the head mobility coefficient. Such a piecewise-constant force density is appropriate in the limit of large aspect ratios (Batchelor Reference Batchelor1970a
), but nevertheless yields a bacterium velocity field in good agreement with experimental observations (Drescher et al.
Reference Drescher, Dunkel, Cisneros, Ganguly and Goldstein2011; Kasyap, Koch & Wu Reference Kasyap, Koch and Wu2014). The assumed
$M$
is the head mobility coefficient. Such a piecewise-constant force density is appropriate in the limit of large aspect ratios (Batchelor Reference Batchelor1970a
), but nevertheless yields a bacterium velocity field in good agreement with experimental observations (Drescher et al.
Reference Drescher, Dunkel, Cisneros, Ganguly and Goldstein2011; Kasyap, Koch & Wu Reference Kasyap, Koch and Wu2014). The assumed
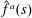 $\hat{f}^{a}(s)$
corresponds to a dimensional force of
$\hat{f}^{a}(s)$
corresponds to a dimensional force of
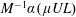 $M^{-1}\unicode[STIX]{x1D6FC}(\unicode[STIX]{x1D707}UL)$
on the head and an intrinsic dipole magnitude of
$M^{-1}\unicode[STIX]{x1D6FC}(\unicode[STIX]{x1D707}UL)$
on the head and an intrinsic dipole magnitude of
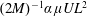 $(2M)^{-1}\unicode[STIX]{x1D6FC}\unicode[STIX]{x1D707}UL^{2}$
. The expression for
$(2M)^{-1}\unicode[STIX]{x1D6FC}\unicode[STIX]{x1D707}UL^{2}$
. The expression for
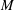 $M$
is known since the E. coli head is a spheroid of aspect ratio
$M$
is known since the E. coli head is a spheroid of aspect ratio
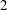 $2$
. The particular wild-type strain (
$2$
. The particular wild-type strain (
 $ATCC9637$
) that we focus on for the experiment–theory comparison has a major axis of
$ATCC9637$
) that we focus on for the experiment–theory comparison has a major axis of
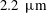 $2.2~\unicode[STIX]{x03BC}\text{m}$
with the minor axis determined from the stated head volume of
$2.2~\unicode[STIX]{x03BC}\text{m}$
with the minor axis determined from the stated head volume of
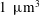 $1~\unicode[STIX]{x03BC}\text{m}^{3}$
. With swimming speeds of
$1~\unicode[STIX]{x03BC}\text{m}^{3}$
. With swimming speeds of
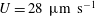 $U=28~\unicode[STIX]{x03BC}\text{m}~\text{s}^{-1}$
and
$U=28~\unicode[STIX]{x03BC}\text{m}~\text{s}^{-1}$
and
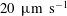 $20~\unicode[STIX]{x03BC}\text{m}~\text{s}^{-1}$
, under oxygen-rich and oxygen-deprived conditions (Lopez et al.
Reference Lopez, Gachelin, Douarche, Auradou and Clement2015), respectively, one obtains force-dipole estimates of 1.26 pN
$20~\unicode[STIX]{x03BC}\text{m}~\text{s}^{-1}$
, under oxygen-rich and oxygen-deprived conditions (Lopez et al.
Reference Lopez, Gachelin, Douarche, Auradou and Clement2015), respectively, one obtains force-dipole estimates of 1.26 pN
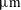 $\unicode[STIX]{x03BC}\text{m}$
and 0.9 pN
$\unicode[STIX]{x03BC}\text{m}$
and 0.9 pN
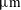 $\unicode[STIX]{x03BC}\text{m}$
, respectively, which compare well with the estimate of 0.8 pN
$\unicode[STIX]{x03BC}\text{m}$
, respectively, which compare well with the estimate of 0.8 pN
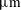 $\unicode[STIX]{x03BC}\text{m}$
obtained from a direct measurement of the disturbance field of a non-tumbling strain of E. coli (Drescher et al.
Reference Drescher, Dunkel, Cisneros, Ganguly and Goldstein2011). Additionally, the hydrodynamic interactions between the head and flagellum, neglected in the
$\unicode[STIX]{x03BC}\text{m}$
obtained from a direct measurement of the disturbance field of a non-tumbling strain of E. coli (Drescher et al.
Reference Drescher, Dunkel, Cisneros, Ganguly and Goldstein2011). Additionally, the hydrodynamic interactions between the head and flagellum, neglected in the
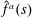 $\hat{f}^{a}(s)$
above, have been shown to lead to only minor modifications (Drescher et al.
Reference Drescher, Dunkel, Cisneros, Ganguly and Goldstein2011).
$\hat{f}^{a}(s)$
above, have been shown to lead to only minor modifications (Drescher et al.
Reference Drescher, Dunkel, Cisneros, Ganguly and Goldstein2011).
Using the
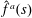 $\hat{f}^{a}(s)$
above, and evaluating the orientation integrals in (2.20), leads to:
$\hat{f}^{a}(s)$
above, and evaluating the orientation integrals in (2.20), leads to:
 $$\begin{eqnarray}\displaystyle \hspace{-15.00002pt}\langle \unicode[STIX]{x1D70E}_{12}\rangle & = & \displaystyle \left[1+(nL^{3})\frac{\unicode[STIX]{x03C0}}{90(\ln \unicode[STIX]{x1D705})}\right]\hat{\dot{\unicode[STIX]{x1D6FE}}}(t)\nonumber\\ \displaystyle & & \displaystyle -\,(nL^{3})\frac{U\unicode[STIX]{x1D70F}}{L}\frac{\unicode[STIX]{x1D6FC}}{10M}\int _{t^{\prime }}^{t}\exp \left[-\left(1-\frac{4\unicode[STIX]{x03C0}A_{2}}{5}+6\unicode[STIX]{x1D70F}D_{r}\right)(t-t^{\prime \prime })\right]\hat{\dot{\unicode[STIX]{x1D6FE}}}(t^{\prime \prime })\,\text{d}t^{\prime \prime },\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle \hspace{-15.00002pt}\langle \unicode[STIX]{x1D70E}_{12}\rangle & = & \displaystyle \left[1+(nL^{3})\frac{\unicode[STIX]{x03C0}}{90(\ln \unicode[STIX]{x1D705})}\right]\hat{\dot{\unicode[STIX]{x1D6FE}}}(t)\nonumber\\ \displaystyle & & \displaystyle -\,(nL^{3})\frac{U\unicode[STIX]{x1D70F}}{L}\frac{\unicode[STIX]{x1D6FC}}{10M}\int _{t^{\prime }}^{t}\exp \left[-\left(1-\frac{4\unicode[STIX]{x03C0}A_{2}}{5}+6\unicode[STIX]{x1D70F}D_{r}\right)(t-t^{\prime \prime })\right]\hat{\dot{\unicode[STIX]{x1D6FE}}}(t^{\prime \prime })\,\text{d}t^{\prime \prime },\end{eqnarray}$$
for the total shear stress measured in units of the solvent stress (
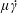 $\unicode[STIX]{x1D707}\dot{\unicode[STIX]{x1D6FE}}$
). As expected, the bacterial stress contribution is
$\unicode[STIX]{x1D707}\dot{\unicode[STIX]{x1D6FE}}$
). As expected, the bacterial stress contribution is
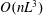 $O(nL^{3})$
in the dilute regime. The active component is opposite in sign to the solvent and passive stress contributions (which have been combined into a single viscous contribution proportional to
$O(nL^{3})$
in the dilute regime. The active component is opposite in sign to the solvent and passive stress contributions (which have been combined into a single viscous contribution proportional to
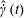 $\hat{\dot{\unicode[STIX]{x1D6FE}}}(t)$
), and is proportional to
$\hat{\dot{\unicode[STIX]{x1D6FE}}}(t)$
), and is proportional to
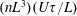 $(nL^{3})(U\unicode[STIX]{x1D70F}/L)$
times a dimensionless integral over the shear rate history that is a function of
$(nL^{3})(U\unicode[STIX]{x1D70F}/L)$
times a dimensionless integral over the shear rate history that is a function of
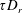 $\unicode[STIX]{x1D70F}D_{r}$
.
$\unicode[STIX]{x1D70F}D_{r}$
.
3 Results
To analyse the step-shear experiments, we take
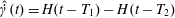 $\hat{\dot{\unicode[STIX]{x1D6FE}}}(t)=H(t-T_{1})-H(t-T_{2})$
, where
$\hat{\dot{\unicode[STIX]{x1D6FE}}}(t)=H(t-T_{1})-H(t-T_{2})$
, where
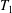 $T_{1}$
and
$T_{1}$
and
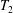 $T_{2}$
are the dimensionless times of shear initiation and termination (in units of
$T_{2}$
are the dimensionless times of shear initiation and termination (in units of
 $\unicode[STIX]{x1D70F}$
). Substitution in (2.21) leads to:
$\unicode[STIX]{x1D70F}$
). Substitution in (2.21) leads to:
 $$\begin{eqnarray}\displaystyle \langle \unicode[STIX]{x1D70E}_{12}\rangle & = & \displaystyle \left[1+(nL^{3})\frac{\unicode[STIX]{x03C0}}{90(\ln \unicode[STIX]{x1D705})}-(nL^{3})\left(\frac{U\unicode[STIX]{x1D70F}}{L}\right)\frac{\unicode[STIX]{x1D6FC}}{10MB}\right]\hat{\dot{\unicode[STIX]{x1D6FE}}}(t)\nonumber\\ \displaystyle & & \displaystyle -\,(nL^{3})\left(\frac{U\unicode[STIX]{x1D70F}}{L}\right)\frac{\unicode[STIX]{x1D6FC}}{10MB}[H(t-T_{2})\exp (B(T_{2}-t))-H(t-T_{1})\exp (B(T_{1}-t))],\nonumber\\ \displaystyle & & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle \langle \unicode[STIX]{x1D70E}_{12}\rangle & = & \displaystyle \left[1+(nL^{3})\frac{\unicode[STIX]{x03C0}}{90(\ln \unicode[STIX]{x1D705})}-(nL^{3})\left(\frac{U\unicode[STIX]{x1D70F}}{L}\right)\frac{\unicode[STIX]{x1D6FC}}{10MB}\right]\hat{\dot{\unicode[STIX]{x1D6FE}}}(t)\nonumber\\ \displaystyle & & \displaystyle -\,(nL^{3})\left(\frac{U\unicode[STIX]{x1D70F}}{L}\right)\frac{\unicode[STIX]{x1D6FC}}{10MB}[H(t-T_{2})\exp (B(T_{2}-t))-H(t-T_{1})\exp (B(T_{1}-t))],\nonumber\\ \displaystyle & & \displaystyle\end{eqnarray}$$
where
 $B=3[(\unicode[STIX]{x1D6FD}\coth \unicode[STIX]{x1D6FD}-1)/\unicode[STIX]{x1D6FD}^{2}+2\unicode[STIX]{x1D70F}D_{r}]$
with
$B=3[(\unicode[STIX]{x1D6FD}\coth \unicode[STIX]{x1D6FD}-1)/\unicode[STIX]{x1D6FD}^{2}+2\unicode[STIX]{x1D70F}D_{r}]$
with
 $B^{-1}\unicode[STIX]{x1D70F}$
denoting the time scale of relaxation of the active-stress contribution arising from a combination of run-and-tumble dynamics and rotary diffusion. Although we focus on a step-shear here, it is worth noting that the memory integral in (2.21) involves a single exponential kernel, and the active response in oscillatory shear will therefore be that of a single-mode Maxwell-type fluid with
$B^{-1}\unicode[STIX]{x1D70F}$
denoting the time scale of relaxation of the active-stress contribution arising from a combination of run-and-tumble dynamics and rotary diffusion. Although we focus on a step-shear here, it is worth noting that the memory integral in (2.21) involves a single exponential kernel, and the active response in oscillatory shear will therefore be that of a single-mode Maxwell-type fluid with
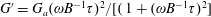 $G^{\prime }=G_{a}(\unicode[STIX]{x1D714}B^{-1}\unicode[STIX]{x1D70F})^{2}/[(1+(\unicode[STIX]{x1D714}B^{-1}\unicode[STIX]{x1D70F})^{2}]$
and
$G^{\prime }=G_{a}(\unicode[STIX]{x1D714}B^{-1}\unicode[STIX]{x1D70F})^{2}/[(1+(\unicode[STIX]{x1D714}B^{-1}\unicode[STIX]{x1D70F})^{2}]$
and
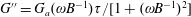 $G^{\prime \prime }=G_{a}(\unicode[STIX]{x1D714}B^{-1})\unicode[STIX]{x1D70F}/[1+(\unicode[STIX]{x1D714}B^{-1})^{2}]$
, where the active modulus
$G^{\prime \prime }=G_{a}(\unicode[STIX]{x1D714}B^{-1})\unicode[STIX]{x1D70F}/[1+(\unicode[STIX]{x1D714}B^{-1})^{2}]$
, where the active modulus
 $G_{a}=(\dot{\unicode[STIX]{x1D6FE}}\unicode[STIX]{x1D70F}/5)((n\unicode[STIX]{x1D6FC}/(2M))(\unicode[STIX]{x1D707}UL^{2}))$
, and may be interpreted as the product of an
$G_{a}=(\dot{\unicode[STIX]{x1D6FE}}\unicode[STIX]{x1D70F}/5)((n\unicode[STIX]{x1D6FC}/(2M))(\unicode[STIX]{x1D707}UL^{2}))$
, and may be interpreted as the product of an
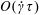 $O(\dot{\unicode[STIX]{x1D6FE}}\unicode[STIX]{x1D70F})$
orientation anisotropy and an
$O(\dot{\unicode[STIX]{x1D6FE}}\unicode[STIX]{x1D70F})$
orientation anisotropy and an
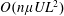 $O(n\unicode[STIX]{x1D707}UL^{2})$
dipole density.
$O(n\unicode[STIX]{x1D707}UL^{2})$
dipole density.
For
 $T_{2}-T_{1}\gg B^{-1}$
, the experimentally relevant limit, the suspension viscosity based on (3.1) attains a steady value well before shear termination, given by
$T_{2}-T_{1}\gg B^{-1}$
, the experimentally relevant limit, the suspension viscosity based on (3.1) attains a steady value well before shear termination, given by
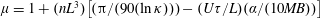 $\unicode[STIX]{x1D707}=1+(nL^{3})\big[(\unicode[STIX]{x03C0}/(90(\ln \unicode[STIX]{x1D705})))-(U\unicode[STIX]{x1D70F}/L)(\unicode[STIX]{x1D6FC}/(10MB))\big]$
. The viscosity vanishes at a critical hydrodynamic volume fraction given by
$\unicode[STIX]{x1D707}=1+(nL^{3})\big[(\unicode[STIX]{x03C0}/(90(\ln \unicode[STIX]{x1D705})))-(U\unicode[STIX]{x1D70F}/L)(\unicode[STIX]{x1D6FC}/(10MB))\big]$
. The viscosity vanishes at a critical hydrodynamic volume fraction given by
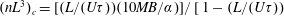 $(nL^{3})_{c}=[(L/(U\unicode[STIX]{x1D70F}))(10MB/\unicode[STIX]{x1D6FC})]/[1-(L/(U\unicode[STIX]{x1D70F}))$
$(nL^{3})_{c}=[(L/(U\unicode[STIX]{x1D70F}))(10MB/\unicode[STIX]{x1D6FC})]/[1-(L/(U\unicode[STIX]{x1D70F}))$
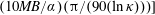 $(10MB/\unicode[STIX]{x1D6FC})(\unicode[STIX]{x03C0}/(90(\ln \unicode[STIX]{x1D705})))\!]$
, leading to an instability, associated with exponentially growing long wavelength orientation and velocity fluctuations at higher volume fractions. This long-wavelength negative-viscosity instability was interpreted by Subramanian & Koch (Reference Subramanian and Koch2009) as being responsible for the onset of spontaneous collective motion in an otherwise quiescent bacterial suspension (the analogous instability in a suspension of straight swimmers was independently identified by Saintillan & Shelley (Reference Saintillan and Shelley2008) and Underhill, Hernandez-Ortiz & Graham (Reference Underhill, Hernandez-Ortiz and Graham2008)). In Lopez et al. (Reference Lopez, Gachelin, Douarche, Auradou and Clement2015), an expression for
$(10MB/\unicode[STIX]{x1D6FC})(\unicode[STIX]{x03C0}/(90(\ln \unicode[STIX]{x1D705})))\!]$
, leading to an instability, associated with exponentially growing long wavelength orientation and velocity fluctuations at higher volume fractions. This long-wavelength negative-viscosity instability was interpreted by Subramanian & Koch (Reference Subramanian and Koch2009) as being responsible for the onset of spontaneous collective motion in an otherwise quiescent bacterial suspension (the analogous instability in a suspension of straight swimmers was independently identified by Saintillan & Shelley (Reference Saintillan and Shelley2008) and Underhill, Hernandez-Ortiz & Graham (Reference Underhill, Hernandez-Ortiz and Graham2008)). In Lopez et al. (Reference Lopez, Gachelin, Douarche, Auradou and Clement2015), an expression for
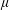 $\unicode[STIX]{x1D707}$
similar to that above was given, but the
$\unicode[STIX]{x1D707}$
similar to that above was given, but the
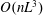 $O(nL^{3})$
active contribution involved two empirically determined relaxation times (see supplementary material of their article), one of which was incorrectly related to the directional persistence of a bacterium trajectory (rather than
$O(nL^{3})$
active contribution involved two empirically determined relaxation times (see supplementary material of their article), one of which was incorrectly related to the directional persistence of a bacterium trajectory (rather than
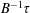 $B^{-1}\unicode[STIX]{x1D70F}$
above), while the other was related to a critical shear rate (clearly inappropriate for a linear response regime where all relaxation process are intrinsic and independent of the imposed forcing).
$B^{-1}\unicode[STIX]{x1D70F}$
above), while the other was related to a critical shear rate (clearly inappropriate for a linear response regime where all relaxation process are intrinsic and independent of the imposed forcing).

Figure 2. The suspension shear stress plotted as a function of time under oxygen-deprived (a) and oxygen-rich (b) conditions. The blue (dashed) lines represent the experimental data extracted from Lopez et al. (Reference Lopez, Gachelin, Douarche, Auradou and Clement2015). The red (dotted) and green (solid) lines denote the theoretical predictions for two different bacterium lengths.

Figure 3. (a) Theoretically predicted stress relaxations over the experimental range of volume fractions (
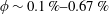 $\unicode[STIX]{x1D719}\sim 0.1\,\%{-}0.67\,\%$
;
$\unicode[STIX]{x1D719}\sim 0.1\,\%{-}0.67\,\%$
;
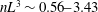 $nL^{3}\sim 0.56{-}3.43$
for
$nL^{3}\sim 0.56{-}3.43$
for
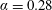 $\unicode[STIX]{x1D6FC}=0.28$
). (b) The zero shear viscosity as a function of the volume fraction for
$\unicode[STIX]{x1D6FC}=0.28$
). (b) The zero shear viscosity as a function of the volume fraction for
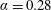 $\unicode[STIX]{x1D6FC}=0.28$
- blue (solid) and
$\unicode[STIX]{x1D6FC}=0.28$
- blue (solid) and
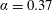 $\unicode[STIX]{x1D6FC}=0.37$
– red (dotted); the brown (dashed) line denotes the experimental straight line fit (Lopez et al.
Reference Lopez, Gachelin, Douarche, Auradou and Clement2015).
$\unicode[STIX]{x1D6FC}=0.37$
– red (dotted); the brown (dashed) line denotes the experimental straight line fit (Lopez et al.
Reference Lopez, Gachelin, Douarche, Auradou and Clement2015).

Figure 4. The suspension shear stress is plotted as a function of time under oxygen-deprived (a) and oxygen-rich (b) conditions. The blue (dashed) lines represent the experimental curves as extracted from Lopez et al. (Reference Lopez, Gachelin, Douarche, Auradou and Clement2015). The red (dotted) and green (solid) lines denote the theoretical predictions for two different bacterium lengths based on the translational diffusivity (
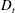 $D_{t}$
) presented in the supplementary data by the authors.
$D_{t}$
) presented in the supplementary data by the authors.

Figure 5. The suspension shear stress is plotted as a function of time under oxygen-deprived (a) and oxygen-rich (b) conditions. The blue (dashed) lines represent the experimental curves as extracted from Lopez et al. (Reference Lopez, Gachelin, Douarche, Auradou and Clement2015). The red (dotted) and green (solid) lines denote the theoretical predictions for two different bacterium lengths based on the large stress relaxation times observed in the experiments.
For purposes of comparison, we first generate theoretical predictions from (3.1) assuming
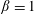 $\unicode[STIX]{x1D6FD}=1$
and
$\unicode[STIX]{x1D6FD}=1$
and
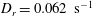 $D_{r}=0.062~\text{s}^{-1}$
, both corresponding to the original measurements of Berg (Reference Berg1993), and in addition, take
$D_{r}=0.062~\text{s}^{-1}$
, both corresponding to the original measurements of Berg (Reference Berg1993), and in addition, take
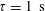 $\unicode[STIX]{x1D70F}=1~\text{s}$
as a typical tumble time. The experiments under oxygen-deprived conditions (with L-serine sustaining bacterial swimming activity) correspond to the stable regime, as evident from the positive plateau stress values, attained in the limit of long times after shear initiation, for all shear rates and for volume fractions ranging from
$\unicode[STIX]{x1D70F}=1~\text{s}$
as a typical tumble time. The experiments under oxygen-deprived conditions (with L-serine sustaining bacterial swimming activity) correspond to the stable regime, as evident from the positive plateau stress values, attained in the limit of long times after shear initiation, for all shear rates and for volume fractions ranging from
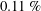 $0.11\,\%$
to
$0.11\,\%$
to
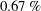 $0.67\,\%$
(figure 1a in Lopez et al.
Reference Lopez, Gachelin, Douarche, Auradou and Clement2015). In contrast, their experiments with suspensions of hyper-activated strains (in the presence of both oxygen and L-serine) show plateau values close to zero, above a threshold volume fraction, for sufficiently small shear rates (their figure 2a). The bacterial suspension in this regime acts, in effect, as a superfluid in response to macroscopic velocity fluctuations. The dashed curve in figure 2(a) is the experimental stress response for wild-type ATCC9637, for a true volume fraction
$0.67\,\%$
(figure 1a in Lopez et al.
Reference Lopez, Gachelin, Douarche, Auradou and Clement2015). In contrast, their experiments with suspensions of hyper-activated strains (in the presence of both oxygen and L-serine) show plateau values close to zero, above a threshold volume fraction, for sufficiently small shear rates (their figure 2a). The bacterial suspension in this regime acts, in effect, as a superfluid in response to macroscopic velocity fluctuations. The dashed curve in figure 2(a) is the experimental stress response for wild-type ATCC9637, for a true volume fraction
 $\unicode[STIX]{x1D719}=(\unicode[STIX]{x03C0}/6)(\unicode[STIX]{x1D6FC}nL^{3}/\unicode[STIX]{x1D705}^{2})=0.0067$
, under oxygen-deprived conditions, and for a shear rate
$\unicode[STIX]{x1D719}=(\unicode[STIX]{x03C0}/6)(\unicode[STIX]{x1D6FC}nL^{3}/\unicode[STIX]{x1D705}^{2})=0.0067$
, under oxygen-deprived conditions, and for a shear rate
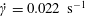 $\dot{\unicode[STIX]{x1D6FE}}=0.022~\text{s}^{-1}$
; the observed stress responses are coincident for
$\dot{\unicode[STIX]{x1D6FE}}=0.022~\text{s}^{-1}$
; the observed stress responses are coincident for
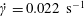 $\dot{\unicode[STIX]{x1D6FE}}=0.022~\text{s}^{-1}$
,
$\dot{\unicode[STIX]{x1D6FE}}=0.022~\text{s}^{-1}$
,
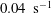 $0.04~\text{s}^{-1}$
and
$0.04~\text{s}^{-1}$
and
 $0.075~\text{s}^{-1}$
, suggesting that they are representative of the linear response regime. The pair of curves shown alongside are the predictions for the same
$0.075~\text{s}^{-1}$
, suggesting that they are representative of the linear response regime. The pair of curves shown alongside are the predictions for the same
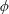 $\unicode[STIX]{x1D719}$
, with
$\unicode[STIX]{x1D719}$
, with
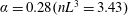 $\unicode[STIX]{x1D6FC}=0.28(nL^{3}=3.43)$
and
$\unicode[STIX]{x1D6FC}=0.28(nL^{3}=3.43)$
and
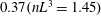 $0.37(nL^{3}=1.45)$
. The range of
$0.37(nL^{3}=1.45)$
. The range of
 $\unicode[STIX]{x1D6FC}$
between these values corresponds to the range of bacterium lengths (
$\unicode[STIX]{x1D6FC}$
between these values corresponds to the range of bacterium lengths (
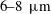 $6{-}8~\unicode[STIX]{x03BC}\text{m}$
) in the experiments (Gachelin et al.
Reference Gachelin, Rousselet, Lindner and Clement2014), and the curve with
$6{-}8~\unicode[STIX]{x03BC}\text{m}$
) in the experiments (Gachelin et al.
Reference Gachelin, Rousselet, Lindner and Clement2014), and the curve with
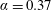 $\unicode[STIX]{x1D6FC}=0.37$
is seen to agree better with the observed response. Figure 2(b) shows the experiment–theory comparison under oxygen-rich conditions, for the same strain, where the shear viscosity asymptotes to a small negative value for long times; the volume fraction here is approximately
$\unicode[STIX]{x1D6FC}=0.37$
is seen to agree better with the observed response. Figure 2(b) shows the experiment–theory comparison under oxygen-rich conditions, for the same strain, where the shear viscosity asymptotes to a small negative value for long times; the volume fraction here is approximately
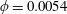 $\unicode[STIX]{x1D719}=0.0054$
(from figure 2b in Lopez et al.
Reference Lopez, Gachelin, Douarche, Auradou and Clement2015). In this case, the theoretical curve with
$\unicode[STIX]{x1D719}=0.0054$
(from figure 2b in Lopez et al.
Reference Lopez, Gachelin, Douarche, Auradou and Clement2015). In this case, the theoretical curve with
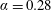 $\unicode[STIX]{x1D6FC}=0.28$
leads to a better comparison. Nevertheless, in both cases, the theoretical curves qualitatively capture the observed responses, including the sign of the stress plateaus.
$\unicode[STIX]{x1D6FC}=0.28$
leads to a better comparison. Nevertheless, in both cases, the theoretical curves qualitatively capture the observed responses, including the sign of the stress plateaus.
Figure 3(a), shows the theoretically predicted temporal stress responses over the experimental range of volume fractions corresponding to
 $nL^{3}\sim 0.56{-}3.43$
(
$nL^{3}\sim 0.56{-}3.43$
(
 $\unicode[STIX]{x1D719}\sim 0.1\,\%{-}0.67\,\%$
) with
$\unicode[STIX]{x1D719}\sim 0.1\,\%{-}0.67\,\%$
) with
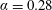 $\unicode[STIX]{x1D6FC}=0.28$
, and corresponding to oxygen-deprived conditions. Figure 3(b) shows the steady viscosities in the low-shear-rate (linear response) regime, corresponding to the long-time plateau values in the temporal responses as a function of the true volume fraction. Also shown is the experimental straight-line fit to the analogous plot in Lopez et al. (Reference Lopez, Gachelin, Douarche, Auradou and Clement2015) (figure 1c therein). The dilute theory, of course, entails a linear decrease of the viscosity with
$\unicode[STIX]{x1D6FC}=0.28$
, and corresponding to oxygen-deprived conditions. Figure 3(b) shows the steady viscosities in the low-shear-rate (linear response) regime, corresponding to the long-time plateau values in the temporal responses as a function of the true volume fraction. Also shown is the experimental straight-line fit to the analogous plot in Lopez et al. (Reference Lopez, Gachelin, Douarche, Auradou and Clement2015) (figure 1c therein). The dilute theory, of course, entails a linear decrease of the viscosity with
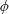 $\unicode[STIX]{x1D719}$
, a decrease first predicted by Hatwalne et al. (Reference Hatwalne, Ramaswamy, Rao and Simha2004). Importantly, the slopes of the theoretical prediction (again for
$\unicode[STIX]{x1D719}$
, a decrease first predicted by Hatwalne et al. (Reference Hatwalne, Ramaswamy, Rao and Simha2004). Importantly, the slopes of the theoretical prediction (again for
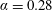 $\unicode[STIX]{x1D6FC}=0.28$
and
$\unicode[STIX]{x1D6FC}=0.28$
and
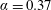 $\unicode[STIX]{x1D6FC}=0.37$
), valid for the low-shear-rate regime, agree reasonably well with the experimental fit.
$\unicode[STIX]{x1D6FC}=0.37$
), valid for the low-shear-rate regime, agree reasonably well with the experimental fit.
The predictions above were generated based on typical values of microscopic swimming parameters (
 $\unicode[STIX]{x1D70F}$
and
$\unicode[STIX]{x1D70F}$
and
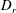 $D_{r}$
). It is also possible to obtain these values from the measured translational diffusivities (
$D_{r}$
). It is also possible to obtain these values from the measured translational diffusivities (
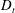 $D_{t}$
) and stress relaxation times (
$D_{t}$
) and stress relaxation times (
 $B^{-1}\unicode[STIX]{x1D70F}$
here;
$B^{-1}\unicode[STIX]{x1D70F}$
here;
 $\unicode[STIX]{x1D70F}_{r}^{\pm }$
in the notation of Lopez et al. (Reference Lopez, Gachelin, Douarche, Auradou and Clement2015), where the superscripts ‘
$\unicode[STIX]{x1D70F}_{r}^{\pm }$
in the notation of Lopez et al. (Reference Lopez, Gachelin, Douarche, Auradou and Clement2015), where the superscripts ‘
 $+$
’ and ‘
$+$
’ and ‘
 $-$
’ refer to the step-up and step-down relaxations). The
$-$
’ refer to the step-up and step-down relaxations). The
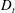 $D_{t}$
values are available from the supplementary material provided by the authors, and the stress relaxation times were obtained by the authors from exponential fits to the temporal stress responses (figure 2c in Lopez et al.
Reference Lopez, Gachelin, Douarche, Auradou and Clement2015). Recall that
$D_{t}$
values are available from the supplementary material provided by the authors, and the stress relaxation times were obtained by the authors from exponential fits to the temporal stress responses (figure 2c in Lopez et al.
Reference Lopez, Gachelin, Douarche, Auradou and Clement2015). Recall that
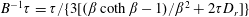 $B^{-1}\unicode[STIX]{x1D70F}=\unicode[STIX]{x1D70F}/\{3[(\unicode[STIX]{x1D6FD}\coth \unicode[STIX]{x1D6FD}-1)/\unicode[STIX]{x1D6FD}^{2}+2\unicode[STIX]{x1D70F}D_{r}]\}$
from theory, and the combined effects of run-and-tumble dynamics and rotary diffusion should lead to a long-time swimmer diffusivity of
$B^{-1}\unicode[STIX]{x1D70F}=\unicode[STIX]{x1D70F}/\{3[(\unicode[STIX]{x1D6FD}\coth \unicode[STIX]{x1D6FD}-1)/\unicode[STIX]{x1D6FD}^{2}+2\unicode[STIX]{x1D70F}D_{r}]\}$
from theory, and the combined effects of run-and-tumble dynamics and rotary diffusion should lead to a long-time swimmer diffusivity of
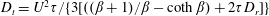 $D_{t}=U^{2}\unicode[STIX]{x1D70F}/\{3[((\unicode[STIX]{x1D6FD}+1)/\unicode[STIX]{x1D6FD}-\coth \unicode[STIX]{x1D6FD})+2\unicode[STIX]{x1D70F}D_{r}]\}$
(Krishnamurthy & Subramanian Reference Krishnamurthy and Subramanian2015). For a given correlation parameter
$D_{t}=U^{2}\unicode[STIX]{x1D70F}/\{3[((\unicode[STIX]{x1D6FD}+1)/\unicode[STIX]{x1D6FD}-\coth \unicode[STIX]{x1D6FD})+2\unicode[STIX]{x1D70F}D_{r}]\}$
(Krishnamurthy & Subramanian Reference Krishnamurthy and Subramanian2015). For a given correlation parameter
 $\unicode[STIX]{x1D6FD}$
, both
$\unicode[STIX]{x1D6FD}$
, both
 $\unicode[STIX]{x1D70F}$
and
$\unicode[STIX]{x1D70F}$
and
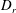 $D_{r}$
may, in principle, be determined since
$D_{r}$
may, in principle, be determined since
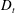 $D_{t}$
and
$D_{t}$
and
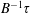 $B^{-1}\unicode[STIX]{x1D70F}$
correspond to different combinations of
$B^{-1}\unicode[STIX]{x1D70F}$
correspond to different combinations of
 $\unicode[STIX]{x1D70F}$
and
$\unicode[STIX]{x1D70F}$
and
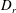 $D_{r}$
. This approach, however, led to unrealistic estimates, especially under oxygen-deprived conditions, primarily because the measured
$D_{r}$
. This approach, however, led to unrealistic estimates, especially under oxygen-deprived conditions, primarily because the measured
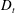 $D_{t}$
values appear too small while the stress relaxation times are too large compared to the
$D_{t}$
values appear too small while the stress relaxation times are too large compared to the
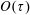 $O(\unicode[STIX]{x1D70F})$
estimates expected based on single-bacterium relaxation. Hence, we fix the
$O(\unicode[STIX]{x1D70F})$
estimates expected based on single-bacterium relaxation. Hence, we fix the
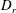 $D_{r}$
and
$D_{r}$
and
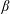 $\unicode[STIX]{x1D6FD}$
values to be the same as those used above, and estimate
$\unicode[STIX]{x1D6FD}$
values to be the same as those used above, and estimate
 $\unicode[STIX]{x1D70F}$
separately based on each of
$\unicode[STIX]{x1D70F}$
separately based on each of
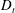 $D_{t}$
and
$D_{t}$
and
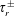 $\unicode[STIX]{x1D70F}_{r}^{\pm }$
. Figures 4 and 5 show experiment–theory comparisons similar to those in figure 2, for both oxygen-deprived and oxygen-rich conditions, and correspond, respectively, to
$\unicode[STIX]{x1D70F}_{r}^{\pm }$
. Figures 4 and 5 show experiment–theory comparisons similar to those in figure 2, for both oxygen-deprived and oxygen-rich conditions, and correspond, respectively, to
 $\unicode[STIX]{x1D70F}$
-estimates based on the
$\unicode[STIX]{x1D70F}$
-estimates based on the
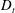 $D_{t}$
and
$D_{t}$
and
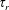 $\unicode[STIX]{x1D70F}_{r}$
measurements. In the oxygen-deprived case,
$\unicode[STIX]{x1D70F}_{r}$
measurements. In the oxygen-deprived case,
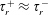 $\unicode[STIX]{x1D70F}_{r}^{+}\approx \unicode[STIX]{x1D70F}_{r}^{-}$
, consistent with theory which predicts each of them to equal
$\unicode[STIX]{x1D70F}_{r}^{+}\approx \unicode[STIX]{x1D70F}_{r}^{-}$
, consistent with theory which predicts each of them to equal
 $B^{-1}\unicode[STIX]{x1D70F}$
;
$B^{-1}\unicode[STIX]{x1D70F}$
;
 $\unicode[STIX]{x1D70F}_{r}^{-}({\approx}3.2~\text{s})$
was significantly larger than
$\unicode[STIX]{x1D70F}_{r}^{-}({\approx}3.2~\text{s})$
was significantly larger than
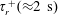 $\unicode[STIX]{x1D70F}_{r}^{+}({\approx}2~\text{s})$
under oxygen-rich conditions, and the latter value was used to estimate
$\unicode[STIX]{x1D70F}_{r}^{+}({\approx}2~\text{s})$
under oxygen-rich conditions, and the latter value was used to estimate
 $\unicode[STIX]{x1D70F}$
in this case. It is clear that the
$\unicode[STIX]{x1D70F}$
in this case. It is clear that the
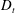 $D_{t}$
-based estimates lead to a serious underprediction by theory in figure 4. This is essentially because the measured values of
$D_{t}$
-based estimates lead to a serious underprediction by theory in figure 4. This is essentially because the measured values of
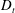 $D_{t}$
led to unrealistically small
$D_{t}$
led to unrealistically small
 $\unicode[STIX]{x1D70F}$
values. For instance,
$\unicode[STIX]{x1D70F}$
values. For instance,
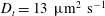 $D_{t}=13~\unicode[STIX]{x03BC}\text{m}^{2}~\text{s}^{-1}$
under serine-rich conditions led to
$D_{t}=13~\unicode[STIX]{x03BC}\text{m}^{2}~\text{s}^{-1}$
under serine-rich conditions led to
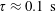 $\unicode[STIX]{x1D70F}\approx 0.1~\text{s}$
, implying that the runs for the ATCC strain under consideration are, in fact, comparable to the duration of typical tumble events (assumed instantaneous here) under oxygen-deprived conditions! Typical values of
$\unicode[STIX]{x1D70F}\approx 0.1~\text{s}$
, implying that the runs for the ATCC strain under consideration are, in fact, comparable to the duration of typical tumble events (assumed instantaneous here) under oxygen-deprived conditions! Typical values of
 $D_{t}$
(
$D_{t}$
(
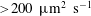 ${>}200~\unicode[STIX]{x03BC}\text{m}^{2}~\text{s}^{-1}$
) quoted elsewhere are much larger (Berg Reference Berg1993; Wu & Libchaber Reference Wu and Libchaber2000). In contrast, the
${>}200~\unicode[STIX]{x03BC}\text{m}^{2}~\text{s}^{-1}$
) quoted elsewhere are much larger (Berg Reference Berg1993; Wu & Libchaber Reference Wu and Libchaber2000). In contrast, the
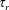 $\unicode[STIX]{x1D70F}_{r}$
-based estimates lead to enormous overpredictions in figure 5 as evident from the large negative magnitudes of the long-time plateaus. We comment on these discrepancies in the concluding section below.
$\unicode[STIX]{x1D70F}_{r}$
-based estimates lead to enormous overpredictions in figure 5 as evident from the large negative magnitudes of the long-time plateaus. We comment on these discrepancies in the concluding section below.
4 Discussion
In this communication, we have presented an analysis that captures the stress response, measured in a transiently sheared bacterial suspension (Lopez et al.
Reference Lopez, Gachelin, Douarche, Auradou and Clement2015), pertaining to the linear response regime. This response has a viscous passive component and an active component with an elastic character arising from the finite time needed for a bacterium to reorient in the imposed shear. Crucially, the elastic contribution is opposite in sign to that encountered in near-equilibrium complex fluids on account of the extensile nature of the bacterium force dipoles. The qualitative trends observed in the experiments are captured by theory in all relevant cases, but there remain quantitative discrepancies. For the best choice of parameters (based on previously reported data on the tumble frequency and rotary diffusivity for E. coli), corresponding to the plot in figure 2(a) for oxygen-deprived conditions, the theoretical relaxations are a second or smaller for the bacterium orientation distribution, and thence, for the active stress. The measured relaxation times are
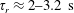 $\unicode[STIX]{x1D70F}_{r}\approx 2{-}3.2~\text{s}$
under varying conditions, which are significantly larger than those expected based on orientation relaxation processes at the single-bacterium level.
$\unicode[STIX]{x1D70F}_{r}\approx 2{-}3.2~\text{s}$
under varying conditions, which are significantly larger than those expected based on orientation relaxation processes at the single-bacterium level.
As things stand, the experimental observations do not allow for an unambiguous interpretation. Any such interpretation depends on whether the bacterial suspension is dilute enough for it to be in the stable regime. The intrinsic dynamics is expected to be very different in the stable and unstable regimes, with the dynamics in the latter regime being dominated by slowly relaxing long-wavelength fluctuations characteristic of collective motion. The linear response will accordingly be different. The threshold prediction by Subramanian & Koch (Reference Subramanian and Koch2009), for the transition of a quiescent bacterial suspension to an unstable regime characterized by spontaneous long-wavelength velocity fluctuations, depends on the choice of microscopic parameters for the ATCC9637 strain. As with the experiment–theory comparisons in the previous section, different choices lead to very different conclusions. The typical choices of
 $\unicode[STIX]{x1D70F}$
and
$\unicode[STIX]{x1D70F}$
and
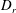 $D_{r}$
corresponding to figure 2 imply marginal stability for the highest volume fraction (
$D_{r}$
corresponding to figure 2 imply marginal stability for the highest volume fraction (
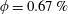 $\unicode[STIX]{x1D719}=0.67\,\%$
), under both oxygen-deprived and hyper-oxygenated conditions. If the experimental range of volume fractions (
$\unicode[STIX]{x1D719}=0.67\,\%$
), under both oxygen-deprived and hyper-oxygenated conditions. If the experimental range of volume fractions (
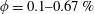 $\unicode[STIX]{x1D719}=0.1{-}0.67\,\%$
) conform to the dilute stable regime, the relaxation times must be independent of the volume fraction, being the same (
$\unicode[STIX]{x1D719}=0.1{-}0.67\,\%$
) conform to the dilute stable regime, the relaxation times must be independent of the volume fraction, being the same (
 $B^{-1}\unicode[STIX]{x1D70F}$
) for both step-up and step-down shear processes. Since the observed stress response is available only at a single volume fraction (
$B^{-1}\unicode[STIX]{x1D70F}$
) for both step-up and step-down shear processes. Since the observed stress response is available only at a single volume fraction (
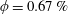 $\unicode[STIX]{x1D719}=0.67\,\%$
) under oxygen-deprived conditions, the volume-fraction-independence cannot be verified. There is a near symmetry between the observed step-up and step-down relaxations (
$\unicode[STIX]{x1D719}=0.67\,\%$
) under oxygen-deprived conditions, the volume-fraction-independence cannot be verified. There is a near symmetry between the observed step-up and step-down relaxations (
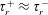 $\unicode[STIX]{x1D70F}_{r}^{+}\approx \unicode[STIX]{x1D70F}_{r}^{-}$
) at the lowest shear rates, as predicted by theory. The observed departure from the zero-shear-rate prediction first occurs at
$\unicode[STIX]{x1D70F}_{r}^{+}\approx \unicode[STIX]{x1D70F}_{r}^{-}$
) at the lowest shear rates, as predicted by theory. The observed departure from the zero-shear-rate prediction first occurs at
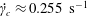 $\dot{\unicode[STIX]{x1D6FE}}_{c}\approx 0.255~\text{s}^{-1}$
with
$\dot{\unicode[STIX]{x1D6FE}}_{c}\approx 0.255~\text{s}^{-1}$
with
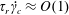 $\unicode[STIX]{x1D70F}_{r}\dot{\unicode[STIX]{x1D6FE}}_{c}\approx O(1)$
(figure 1a in Lopez et al.
Reference Lopez, Gachelin, Douarche, Auradou and Clement2015), implying that the onset of nonlinearity is, in fact, consistent with the larger (in relation to theory) relaxation times measured. Notwithstanding the uncertainty in the stability threshold, the observed relaxation times appear consistent with collective motion. The long relaxation times characteristic of this (unstable) regime are believed to underlie the significantly enhanced tracer particle diffusivities, observed in suspensions of pushers, both in experiments (Wu & Libchaber Reference Wu and Libchaber2000) and simulations (Underhill et al.
Reference Underhill, Hernandez-Ortiz and Graham2008; Krishnamurthy & Subramanian Reference Krishnamurthy and Subramanian2015). Interestingly, recent simulations by Krishnamurthy & Subramanian (Reference Krishnamurthy and Subramanian2015) also indicate a subcritical transition of a suspension of slender swimmers to collective motion (as inferred from the bifurcation of the tracer diffusivity curves in dilute pusher and puller suspensions) even when the suspension viscosity is still positive; a similar behaviour was seen for suspensions of interacting dumbbell-shaped swimmers by Underhill & Graham (Reference Underhill and Graham2011). It is thus tempting to speculate that the bacterial suspension has transitioned to collective motion for volume fractions well below
$\unicode[STIX]{x1D70F}_{r}\dot{\unicode[STIX]{x1D6FE}}_{c}\approx O(1)$
(figure 1a in Lopez et al.
Reference Lopez, Gachelin, Douarche, Auradou and Clement2015), implying that the onset of nonlinearity is, in fact, consistent with the larger (in relation to theory) relaxation times measured. Notwithstanding the uncertainty in the stability threshold, the observed relaxation times appear consistent with collective motion. The long relaxation times characteristic of this (unstable) regime are believed to underlie the significantly enhanced tracer particle diffusivities, observed in suspensions of pushers, both in experiments (Wu & Libchaber Reference Wu and Libchaber2000) and simulations (Underhill et al.
Reference Underhill, Hernandez-Ortiz and Graham2008; Krishnamurthy & Subramanian Reference Krishnamurthy and Subramanian2015). Interestingly, recent simulations by Krishnamurthy & Subramanian (Reference Krishnamurthy and Subramanian2015) also indicate a subcritical transition of a suspension of slender swimmers to collective motion (as inferred from the bifurcation of the tracer diffusivity curves in dilute pusher and puller suspensions) even when the suspension viscosity is still positive; a similar behaviour was seen for suspensions of interacting dumbbell-shaped swimmers by Underhill & Graham (Reference Underhill and Graham2011). It is thus tempting to speculate that the bacterial suspension has transitioned to collective motion for volume fractions well below
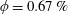 $\unicode[STIX]{x1D719}=0.67\,\%$
. This would seem to resolve any apparent conflict with the aforementioned threshold estimate, but is an aspect that needs verification based on more detailed simulations.
$\unicode[STIX]{x1D719}=0.67\,\%$
. This would seem to resolve any apparent conflict with the aforementioned threshold estimate, but is an aspect that needs verification based on more detailed simulations.
The above comments follow from the best-case scenario for the experiment–theory comparison. Even here, the fact that the presumed existence of collective motion affects only the transient dynamics, and not the steady-state responses (which compare well for oxygen-deprived conditions; see figure 2) is far from obvious. Under oxygen-rich conditions, not only are the observed relaxations longer, there is also a clear asymmetry with the relaxation to quiescence occurring at a substantially slower rate. The magnitudes of the viscous jumps at the shear initiation and termination times also differ perceptibly. In contrast to the above comparisons, using the estimates of
 $\unicode[STIX]{x1D70F}$
based on the measured values of
$\unicode[STIX]{x1D70F}$
based on the measured values of
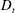 $D_{t}$
and
$D_{t}$
and
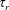 $\unicode[STIX]{x1D70F}_{r}$
measurements leads to large discrepancies between theory and experiment, in both the transient and steady-state responses, even under oxygen-deprived conditions (figures 4 and 5). Although the predictions for the said figures were generated based on a combination of correlated tumbles, the situation remains unchanged even if one uses a prediction based simply on random tumbling with
$\unicode[STIX]{x1D70F}_{r}$
measurements leads to large discrepancies between theory and experiment, in both the transient and steady-state responses, even under oxygen-deprived conditions (figures 4 and 5). Although the predictions for the said figures were generated based on a combination of correlated tumbles, the situation remains unchanged even if one uses a prediction based simply on random tumbling with
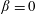 $\unicode[STIX]{x1D6FD}=0$
. The overestimation of the active-stress contribution from the
$\unicode[STIX]{x1D6FD}=0$
. The overestimation of the active-stress contribution from the
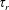 $\unicode[STIX]{x1D70F}_{r}$
-based predictions again points to the presence of collective motion. A smaller active response based on translational diffusivity measurements is not necessarily inconsistent with the measured stress relaxations, given that the diffusivity measurements were carried out offline at a lower volume fraction (
$\unicode[STIX]{x1D70F}_{r}$
-based predictions again points to the presence of collective motion. A smaller active response based on translational diffusivity measurements is not necessarily inconsistent with the measured stress relaxations, given that the diffusivity measurements were carried out offline at a lower volume fraction (
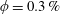 $\unicode[STIX]{x1D719}=0.3\,\%$
). However, as pointed out in § 3, the diffusivity value under oxygen-deprived conditions is much lower than any sensible variation arising from a difference of volume fraction.
$\unicode[STIX]{x1D719}=0.3\,\%$
). However, as pointed out in § 3, the diffusivity value under oxygen-deprived conditions is much lower than any sensible variation arising from a difference of volume fraction.
4.1 The effect of a distribution of run times
A possible cause of the departure of the theoretical predictions from experimental ones could be the inherent variations of the run-and-tumble characteristics between individual cells. The predictions here are for a collection of cells acting individually and assumed to have identical properties, but it is well known that the swimming speed, tumble frequency and rotary diffusivity are distributed over a range among cells, and also that individual cells exhibit stochasticity in their behaviour. The bacterial stress, being proportional to the magnitude of the intrinsic force dipole, is linear in
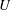 $U$
, and the only effect of a distribution of swimming speeds is that the net active stress now varies as the mean swimming speed
$U$
, and the only effect of a distribution of swimming speeds is that the net active stress now varies as the mean swimming speed
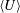 $\langle U\rangle$
. Further, within the linear response framework considered here, and neglecting rotary diffusion (as mentioned earlier, typical values of
$\langle U\rangle$
. Further, within the linear response framework considered here, and neglecting rotary diffusion (as mentioned earlier, typical values of
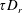 $\unicode[STIX]{x1D70F}D_{r}$
are smaller than unity), the steady-state viscosity is proportional to
$\unicode[STIX]{x1D70F}D_{r}$
are smaller than unity), the steady-state viscosity is proportional to
 $\unicode[STIX]{x1D70F}$
,
$\unicode[STIX]{x1D70F}$
,
 $\unicode[STIX]{x1D70F}$
here being the mean tumble time obtained from the extended run-and-tumble trajectory of a single bacterium (single-bacterium
$\unicode[STIX]{x1D70F}$
here being the mean tumble time obtained from the extended run-and-tumble trajectory of a single bacterium (single-bacterium
 $\unicode[STIX]{x1D70F}$
measurements were originally carried out by Berg and co-workers with the aid of a tracking microscope (Berg & Brown Reference Berg and Brown1972)). Averaging the steady-state response over a distribution of
$\unicode[STIX]{x1D70F}$
measurements were originally carried out by Berg and co-workers with the aid of a tracking microscope (Berg & Brown Reference Berg and Brown1972)). Averaging the steady-state response over a distribution of
 $\unicode[STIX]{x1D70F}$
values again has the effect of merely replacing
$\unicode[STIX]{x1D70F}$
values again has the effect of merely replacing
 $\unicode[STIX]{x1D70F}$
by
$\unicode[STIX]{x1D70F}$
by
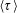 $\langle \unicode[STIX]{x1D70F}\rangle$
, provided the distributions of
$\langle \unicode[STIX]{x1D70F}\rangle$
, provided the distributions of
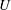 $U$
and
$U$
and
 $\unicode[STIX]{x1D70F}$
are uncorrelated. The effect of averaging the transient (active) response is less trivial. Given that
$\unicode[STIX]{x1D70F}$
are uncorrelated. The effect of averaging the transient (active) response is less trivial. Given that
 $\unicode[STIX]{x1D70F}$
must be positive, a reasonable candidate for the distribution of
$\unicode[STIX]{x1D70F}$
must be positive, a reasonable candidate for the distribution of
 $\unicode[STIX]{x1D70F}$
values is the Gamma distribution defined as
$\unicode[STIX]{x1D70F}$
values is the Gamma distribution defined as
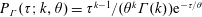 $P_{\unicode[STIX]{x1D6E4}}(\unicode[STIX]{x1D70F};k,\unicode[STIX]{x1D703})=\unicode[STIX]{x1D70F}^{k-1}/(\unicode[STIX]{x1D703}^{k}\unicode[STIX]{x1D6E4}(k))\text{e}^{-\unicode[STIX]{x1D70F}/\unicode[STIX]{x1D703}}$
, where
$P_{\unicode[STIX]{x1D6E4}}(\unicode[STIX]{x1D70F};k,\unicode[STIX]{x1D703})=\unicode[STIX]{x1D70F}^{k-1}/(\unicode[STIX]{x1D703}^{k}\unicode[STIX]{x1D6E4}(k))\text{e}^{-\unicode[STIX]{x1D70F}/\unicode[STIX]{x1D703}}$
, where
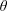 $\unicode[STIX]{x1D703}$
and
$\unicode[STIX]{x1D703}$
and
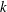 $k$
are the scale and shape factors, respectively, with
$k$
are the scale and shape factors, respectively, with
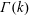 $\unicode[STIX]{x1D6E4}(k)$
being the Gamma function (Abramowitz & Stegun Reference Abramowitz and Stegun1970). Fixing the mean (
$\unicode[STIX]{x1D6E4}(k)$
being the Gamma function (Abramowitz & Stegun Reference Abramowitz and Stegun1970). Fixing the mean (
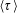 $\langle \unicode[STIX]{x1D70F}\rangle$
) leads to a one-parameter family of distributions with
$\langle \unicode[STIX]{x1D70F}\rangle$
) leads to a one-parameter family of distributions with
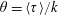 $\unicode[STIX]{x1D703}=\langle \unicode[STIX]{x1D70F}\rangle /k$
. The distribution
$\unicode[STIX]{x1D703}=\langle \unicode[STIX]{x1D70F}\rangle /k$
. The distribution
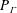 $P_{\unicode[STIX]{x1D6E4}}$
approaches a delta function for
$P_{\unicode[STIX]{x1D6E4}}$
approaches a delta function for
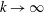 $k\rightarrow \infty$
, leading to the exponential relaxation in (3.1). For finite
$k\rightarrow \infty$
, leading to the exponential relaxation in (3.1). For finite
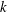 $k$
, the averaged transient is no longer an exponential decay. The
$k$
, the averaged transient is no longer an exponential decay. The
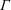 $\unicode[STIX]{x1D6E4}$
-averaged stress relaxation, in the absence of rotary diffusion, is given by:
$\unicode[STIX]{x1D6E4}$
-averaged stress relaxation, in the absence of rotary diffusion, is given by:
 $$\begin{eqnarray}\displaystyle \langle \unicode[STIX]{x1D70E}_{12a}\rangle _{\unicode[STIX]{x1D6E4}} & = & \displaystyle -(nL^{3})\left(\frac{U}{L}\right)\frac{\unicode[STIX]{x1D6FC}\langle \unicode[STIX]{x1D70F}\rangle }{10MB^{\prime }}\left[\hat{\dot{\unicode[STIX]{x1D6FE}}}(t)+2\left(\frac{k^{(k-1)/2}}{\unicode[STIX]{x1D6E4}(k)}\right)\left(\frac{B^{\prime }}{\langle \unicode[STIX]{x1D70F}\rangle }\right)^{(k+1)/2}\right.\nonumber\\ \displaystyle & & \displaystyle \times \!\left[H(t-T_{2})(t-T_{2})^{(k+1)/2}K_{k+1}\left\{2\left(\frac{kB^{\prime }(t-T_{2})}{\langle \unicode[STIX]{x1D70F}\rangle }\right)^{1/2}\right\}\right.\nonumber\\ \displaystyle & & \displaystyle \left.\left.-\,H(t-T_{1})(t-T_{1})^{(k+1)/2}K_{k+1}\left\{2\left(\frac{kB^{\prime }(t-T_{1})}{\langle \unicode[STIX]{x1D70F}\rangle }\right)^{1/2}\right\}\vphantom{\frac{k^{k-1/2}}{\unicode[STIX]{x1D6E4}(k)}}\right]\right],\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle \langle \unicode[STIX]{x1D70E}_{12a}\rangle _{\unicode[STIX]{x1D6E4}} & = & \displaystyle -(nL^{3})\left(\frac{U}{L}\right)\frac{\unicode[STIX]{x1D6FC}\langle \unicode[STIX]{x1D70F}\rangle }{10MB^{\prime }}\left[\hat{\dot{\unicode[STIX]{x1D6FE}}}(t)+2\left(\frac{k^{(k-1)/2}}{\unicode[STIX]{x1D6E4}(k)}\right)\left(\frac{B^{\prime }}{\langle \unicode[STIX]{x1D70F}\rangle }\right)^{(k+1)/2}\right.\nonumber\\ \displaystyle & & \displaystyle \times \!\left[H(t-T_{2})(t-T_{2})^{(k+1)/2}K_{k+1}\left\{2\left(\frac{kB^{\prime }(t-T_{2})}{\langle \unicode[STIX]{x1D70F}\rangle }\right)^{1/2}\right\}\right.\nonumber\\ \displaystyle & & \displaystyle \left.\left.-\,H(t-T_{1})(t-T_{1})^{(k+1)/2}K_{k+1}\left\{2\left(\frac{kB^{\prime }(t-T_{1})}{\langle \unicode[STIX]{x1D70F}\rangle }\right)^{1/2}\right\}\vphantom{\frac{k^{k-1/2}}{\unicode[STIX]{x1D6E4}(k)}}\right]\right],\end{eqnarray}$$

Figure 6. (a) The
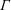 $\unicode[STIX]{x1D6E4}$
-averaged active-stress relaxation plotted as a function of time for a range of values of
$\unicode[STIX]{x1D6E4}$
-averaged active-stress relaxation plotted as a function of time for a range of values of
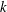 $k$
from 0.1 to 100. (b) The
$k$
from 0.1 to 100. (b) The
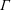 $\unicode[STIX]{x1D6E4}$
-averaged stress relaxation – green (solid), for
$\unicode[STIX]{x1D6E4}$
-averaged stress relaxation – green (solid), for
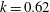 $k=0.62$
is compared to an exponential decay over 2
$k=0.62$
is compared to an exponential decay over 2
 $\langle \unicode[STIX]{x1D70F}\rangle$
– red (dotted) (
$\langle \unicode[STIX]{x1D70F}\rangle$
– red (dotted) (
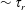 ${\sim}\unicode[STIX]{x1D70F}_{r}$
in the experiments (Lopez et al.
Reference Lopez, Gachelin, Douarche, Auradou and Clement2015)) in (b).
${\sim}\unicode[STIX]{x1D70F}_{r}$
in the experiments (Lopez et al.
Reference Lopez, Gachelin, Douarche, Auradou and Clement2015)) in (b).
with the subscript
 $a$
denoting the active stress. In (4.1),
$a$
denoting the active stress. In (4.1),
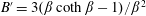 $B^{\prime }=3(\unicode[STIX]{x1D6FD}\coth \unicode[STIX]{x1D6FD}-1)/\unicode[STIX]{x1D6FD}^{2}$
and
$B^{\prime }=3(\unicode[STIX]{x1D6FD}\coth \unicode[STIX]{x1D6FD}-1)/\unicode[STIX]{x1D6FD}^{2}$
and
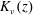 $K_{\unicode[STIX]{x1D708}}(z)$
is the modified Bessel function of the second kind (Abramowitz & Stegun Reference Abramowitz and Stegun1970). The
$K_{\unicode[STIX]{x1D708}}(z)$
is the modified Bessel function of the second kind (Abramowitz & Stegun Reference Abramowitz and Stegun1970). The
 $\unicode[STIX]{x1D6E4}$
-averaged stress relaxations are plotted in figure 6. Figure 6(a) shows the relaxations for different values of
$\unicode[STIX]{x1D6E4}$
-averaged stress relaxations are plotted in figure 6. Figure 6(a) shows the relaxations for different values of
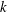 $k$
, while figure 6(b) shows that the prediction of
$k$
, while figure 6(b) shows that the prediction of
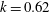 $k=0.62$
comes closest to approximating an exponential decay with a time scale of
$k=0.62$
comes closest to approximating an exponential decay with a time scale of
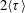 $2\langle \unicode[STIX]{x1D70F}\rangle$
(
$2\langle \unicode[STIX]{x1D70F}\rangle$
(
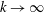 $k\rightarrow \infty$
). It is clear then that a finite variance of tumble times does prolong the stress relaxation, and thereby improve the comparison with the observed transient. We are not aware of the nature of the actual
$k\rightarrow \infty$
). It is clear then that a finite variance of tumble times does prolong the stress relaxation, and thereby improve the comparison with the observed transient. We are not aware of the nature of the actual
 $\unicode[STIX]{x1D70F}$
-distribution, and the only support for the plots in figure 6 is the bacterium-tracking experiments of Berg & Brown (Reference Berg and Brown1972). Figure 4 in their article plots the fraction of runs longer than a specified duration, that is, the complement of the cumulative distribution function of the run duration. If the statistics were exactly Poisson, the plot would be a straight line of slope
$\unicode[STIX]{x1D70F}$
-distribution, and the only support for the plots in figure 6 is the bacterium-tracking experiments of Berg & Brown (Reference Berg and Brown1972). Figure 4 in their article plots the fraction of runs longer than a specified duration, that is, the complement of the cumulative distribution function of the run duration. If the statistics were exactly Poisson, the plot would be a straight line of slope
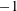 $-1$
on a semi-log graph; the measured run durations across multiple bacteria led to a plot with a positive curvature (curve
$-1$
on a semi-log graph; the measured run durations across multiple bacteria led to a plot with a positive curvature (curve
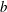 $b$
in figure 4), and we have verified that the upward concavity is consistent with a Gamma distribution for a
$b$
in figure 4), and we have verified that the upward concavity is consistent with a Gamma distribution for a
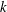 $k$
that is of the same order as that which leads to a relaxation time of
$k$
that is of the same order as that which leads to a relaxation time of
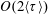 $O(2\langle \unicode[STIX]{x1D70F}\rangle )$
.
$O(2\langle \unicode[STIX]{x1D70F}\rangle )$
.
4.2 Conclusion
On a concluding note, the negative viscosity response, at least to macroscopic velocity variations, is another manifestation of the counter-intuitive behaviour exhibited by bacterial suspensions that is in contrast to the behaviour of near-equilibrium passive suspensions. Other such instances have been demonstrated earlier (Sokolov et al. Reference Sokolov, Apodaca, Grzybowski and Aranson2010). Earlier experiments, for instance those of Aranson & Sokolov (Reference Aranson and Sokolov2009), have indirectly demonstrated a decrease in the shear viscosity with increasing bacteria concentration, with this decrease being directly related to bacterial activity, although the viscosity remained positive for all volume fractions examined. Owing to the low water-like viscosities of dilute bacterial suspensions and the associated torque limitation, direct rheometric measurements had until recently been restricted to volume fractions far beyond the dilute regime (Karmakar et al. Reference Karmakar, Gulvady, Tirumkudulu and Venkatesh2014). The experiments of Lopez et al. (Reference Lopez, Gachelin, Douarche, Auradou and Clement2015) clearly break new ground in terms of being able to access the low-torque regime with the aid of a sensitive rheometer. In future, it would be important to establish the role of collective motion in rheology, and be able to discriminate rheological signatures of the stable and unstable regimes of a bacterial suspension when subject to shear. This requires systematic measurements over a range of volume fractions in the dilute regime with proper characterization of the microscopic swimming parameters. Experiments that simultaneously monitor the rheology and embedded tracer particle displacements would also be crucial; tracer diffusivities have proven to be the most sensitive measure of the onset of collective motion (Underhill et al. Reference Underhill, Hernandez-Ortiz and Graham2008; Saintillan & Shelley Reference Saintillan and Shelley2012; Krishnamurthy & Subramanian Reference Krishnamurthy and Subramanian2015). Moreover, stress-controlled, rather than the strain-rate-controlled experiments carried out in Lopez et al. (Reference Lopez, Gachelin, Douarche, Auradou and Clement2015), would yield valuable insight as previous experiments with concentrated surfactant solutions, and the resulting insight into non-equilibrium jammed-state fluctuations, suggest (Majumdar & Sood Reference Majumdar and Sood2008).
Appendix A. Estimation of the active stochastic stress in a bacterial suspension
Here, we obtain an estimate of the active stochastic stresses due to rotary diffusion and tumbling using an orientation-space-flux-based approach which involves writing the Smoluchowski equation in the form:
 $$\begin{eqnarray}\frac{\unicode[STIX]{x2202}\unicode[STIX]{x1D6FA}}{\unicode[STIX]{x2202}t}++\unicode[STIX]{x1D735}_{\boldsymbol{p}}\boldsymbol{\cdot }(\dot{\boldsymbol{p}}\unicode[STIX]{x1D6FA})+\unicode[STIX]{x1D735}_{\boldsymbol{p}}\boldsymbol{\cdot }(\dot{\boldsymbol{p}}_{st}\unicode[STIX]{x1D6FA})=0,\end{eqnarray}$$
$$\begin{eqnarray}\frac{\unicode[STIX]{x2202}\unicode[STIX]{x1D6FA}}{\unicode[STIX]{x2202}t}++\unicode[STIX]{x1D735}_{\boldsymbol{p}}\boldsymbol{\cdot }(\dot{\boldsymbol{p}}\unicode[STIX]{x1D6FA})+\unicode[STIX]{x1D735}_{\boldsymbol{p}}\boldsymbol{\cdot }(\dot{\boldsymbol{p}}_{st}\unicode[STIX]{x1D6FA})=0,\end{eqnarray}$$
where
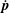 $\dot{\boldsymbol{p}}$
denotes, as before, the rotation due to the ambient shear flow, while
$\dot{\boldsymbol{p}}$
denotes, as before, the rotation due to the ambient shear flow, while
 $\dot{\boldsymbol{p}}_{st}$
denotes the rate of change of orientation due to both rotary diffusion and tumbling. Knowing
$\dot{\boldsymbol{p}}_{st}$
denotes the rate of change of orientation due to both rotary diffusion and tumbling. Knowing
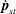 $\dot{\boldsymbol{p}}_{st}$
, the transverse linear force density driving stochastic orientations can be written as
$\dot{\boldsymbol{p}}_{st}$
, the transverse linear force density driving stochastic orientations can be written as
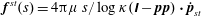 $\boldsymbol{f}^{st}(s)=4\unicode[STIX]{x03C0}\unicode[STIX]{x1D707}\,s/\log \unicode[STIX]{x1D705}(\unicode[STIX]{x1D644}-\boldsymbol{p}\boldsymbol{p})\boldsymbol{\cdot }\dot{\boldsymbol{p}}_{st}$
from viscous slender body theory. The active stochastic stress is then given by the usual formula involving the first moment of this force density:
$\boldsymbol{f}^{st}(s)=4\unicode[STIX]{x03C0}\unicode[STIX]{x1D707}\,s/\log \unicode[STIX]{x1D705}(\unicode[STIX]{x1D644}-\boldsymbol{p}\boldsymbol{p})\boldsymbol{\cdot }\dot{\boldsymbol{p}}_{st}$
from viscous slender body theory. The active stochastic stress is then given by the usual formula involving the first moment of this force density:
 $$\begin{eqnarray}\displaystyle \langle \unicode[STIX]{x1D748}\rangle _{st} & = & \displaystyle \displaystyle -\frac{1}{2}\left\langle \int _{-L/2}^{L/2}s[\boldsymbol{p}\boldsymbol{f}^{st}(s)+\boldsymbol{f}^{st}(s)\boldsymbol{p}]\,\text{d}s\right\rangle ,\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle \langle \unicode[STIX]{x1D748}\rangle _{st} & = & \displaystyle \displaystyle -\frac{1}{2}\left\langle \int _{-L/2}^{L/2}s[\boldsymbol{p}\boldsymbol{f}^{st}(s)+\boldsymbol{f}^{st}(s)\boldsymbol{p}]\,\text{d}s\right\rangle ,\end{eqnarray}$$
 $$\begin{eqnarray}\displaystyle & = & \displaystyle \displaystyle -{\displaystyle \frac{n}{2}}\int \left(\int _{-L/2}^{L/2}s[\boldsymbol{p}\boldsymbol{f}^{st}(s)+\boldsymbol{f}^{st}(s)\boldsymbol{p}]\,\text{d}s\right)\unicode[STIX]{x1D6FA}(\boldsymbol{p})\,\text{d}\boldsymbol{p}.\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle & = & \displaystyle \displaystyle -{\displaystyle \frac{n}{2}}\int \left(\int _{-L/2}^{L/2}s[\boldsymbol{p}\boldsymbol{f}^{st}(s)+\boldsymbol{f}^{st}(s)\boldsymbol{p}]\,\text{d}s\right)\unicode[STIX]{x1D6FA}(\boldsymbol{p})\,\text{d}\boldsymbol{p}.\end{eqnarray}$$
Thus, the key step in the determination of the stochastic stress is identifying
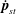 $\dot{\boldsymbol{p}}_{st}$
.
$\dot{\boldsymbol{p}}_{st}$
.
A.1 Stochastic stress due to rotary diffusion and tumbling
The diffusion term in (2.1) can be expressed as
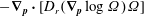 $-\unicode[STIX]{x1D735}_{\boldsymbol{p}}\boldsymbol{\cdot }[D_{r}(\unicode[STIX]{x1D735}_{\boldsymbol{p}}\log \unicode[STIX]{x1D6FA})\unicode[STIX]{x1D6FA}]$
, giving the rate of rotation as
$-\unicode[STIX]{x1D735}_{\boldsymbol{p}}\boldsymbol{\cdot }[D_{r}(\unicode[STIX]{x1D735}_{\boldsymbol{p}}\log \unicode[STIX]{x1D6FA})\unicode[STIX]{x1D6FA}]$
, giving the rate of rotation as
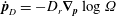 $\dot{\boldsymbol{p}}_{D}=-D_{r}\unicode[STIX]{x1D735}_{\boldsymbol{p}}\log \unicode[STIX]{x1D6FA}$
. The transverse linear force density driving rotary diffusion
$\dot{\boldsymbol{p}}_{D}=-D_{r}\unicode[STIX]{x1D735}_{\boldsymbol{p}}\log \unicode[STIX]{x1D6FA}$
. The transverse linear force density driving rotary diffusion
 $(\boldsymbol{f}^{D})$
is then given as:
$(\boldsymbol{f}^{D})$
is then given as:
 $$\begin{eqnarray}\boldsymbol{f}^{D}(s)=\frac{4\unicode[STIX]{x03C0}\unicode[STIX]{x1D707}s}{\log \unicode[STIX]{x1D705}}(\unicode[STIX]{x1D644}-\boldsymbol{p}\boldsymbol{p})\boldsymbol{\cdot }\dot{\boldsymbol{p}}_{D}=-\frac{4\unicode[STIX]{x03C0}\unicode[STIX]{x1D707}D_{r}s}{\log \unicode[STIX]{x1D705}}(\unicode[STIX]{x1D644}-\boldsymbol{p}\boldsymbol{p})\boldsymbol{\cdot }\unicode[STIX]{x1D735}_{\boldsymbol{p}}(\log \unicode[STIX]{x1D6FA}).\end{eqnarray}$$
$$\begin{eqnarray}\boldsymbol{f}^{D}(s)=\frac{4\unicode[STIX]{x03C0}\unicode[STIX]{x1D707}s}{\log \unicode[STIX]{x1D705}}(\unicode[STIX]{x1D644}-\boldsymbol{p}\boldsymbol{p})\boldsymbol{\cdot }\dot{\boldsymbol{p}}_{D}=-\frac{4\unicode[STIX]{x03C0}\unicode[STIX]{x1D707}D_{r}s}{\log \unicode[STIX]{x1D705}}(\unicode[STIX]{x1D644}-\boldsymbol{p}\boldsymbol{p})\boldsymbol{\cdot }\unicode[STIX]{x1D735}_{\boldsymbol{p}}(\log \unicode[STIX]{x1D6FA}).\end{eqnarray}$$
From (A 3), the stress due to rotary diffusion is:
 $$\begin{eqnarray}\langle \unicode[STIX]{x1D748}\rangle _{D}=-\frac{n}{2}\int \,\text{d}\boldsymbol{p}\unicode[STIX]{x1D6FA}(\boldsymbol{p})\int _{-L/2}^{L/2}s[\boldsymbol{p}\boldsymbol{f}^{D}(s)+\boldsymbol{f}^{D}(s)\boldsymbol{p}]\,\text{d}s,\end{eqnarray}$$
$$\begin{eqnarray}\langle \unicode[STIX]{x1D748}\rangle _{D}=-\frac{n}{2}\int \,\text{d}\boldsymbol{p}\unicode[STIX]{x1D6FA}(\boldsymbol{p})\int _{-L/2}^{L/2}s[\boldsymbol{p}\boldsymbol{f}^{D}(s)+\boldsymbol{f}^{D}(s)\boldsymbol{p}]\,\text{d}s,\end{eqnarray}$$
which on using (A 4), simplifies to:
 $$\begin{eqnarray}\langle \unicode[STIX]{x1D748}\rangle _{D}=\frac{\unicode[STIX]{x03C0}D_{r}(nL^{3})}{\dot{\unicode[STIX]{x1D6FE}}\log \unicode[STIX]{x1D705}}\int \,\text{d}\boldsymbol{p}\unicode[STIX]{x1D6FA}(\boldsymbol{p})\left(\boldsymbol{p}\boldsymbol{p}-\frac{\unicode[STIX]{x1D644}}{3}\right)=\frac{\unicode[STIX]{x03C0}D_{r}(nL^{3})}{\dot{\unicode[STIX]{x1D6FE}}\log \unicode[STIX]{x1D705}}\left\langle \boldsymbol{p}\boldsymbol{p}-\frac{\unicode[STIX]{x1D644}}{3}\right\rangle ,\end{eqnarray}$$
$$\begin{eqnarray}\langle \unicode[STIX]{x1D748}\rangle _{D}=\frac{\unicode[STIX]{x03C0}D_{r}(nL^{3})}{\dot{\unicode[STIX]{x1D6FE}}\log \unicode[STIX]{x1D705}}\int \,\text{d}\boldsymbol{p}\unicode[STIX]{x1D6FA}(\boldsymbol{p})\left(\boldsymbol{p}\boldsymbol{p}-\frac{\unicode[STIX]{x1D644}}{3}\right)=\frac{\unicode[STIX]{x03C0}D_{r}(nL^{3})}{\dot{\unicode[STIX]{x1D6FE}}\log \unicode[STIX]{x1D705}}\left\langle \boldsymbol{p}\boldsymbol{p}-\frac{\unicode[STIX]{x1D644}}{3}\right\rangle ,\end{eqnarray}$$
where we have used the viscous scaling(
 $\unicode[STIX]{x1D707}\dot{\unicode[STIX]{x1D6FE}}$
) for the diffusion stress. Unlike the hydrodynamic stresslet contribution, the leading-order deviatoric contribution in (A 6) arises due to the
$\unicode[STIX]{x1D707}\dot{\unicode[STIX]{x1D6FE}}$
) for the diffusion stress. Unlike the hydrodynamic stresslet contribution, the leading-order deviatoric contribution in (A 6) arises due to the
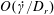 $O(\dot{\unicode[STIX]{x1D6FE}}/D_{r})$
orientation anisotropy. On using the expression for
$O(\dot{\unicode[STIX]{x1D6FE}}/D_{r})$
orientation anisotropy. On using the expression for
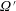 $\unicode[STIX]{x1D6FA}^{\prime }$
from (2.19), and after some simplification, one obtains:
$\unicode[STIX]{x1D6FA}^{\prime }$
from (2.19), and after some simplification, one obtains:
 $$\begin{eqnarray}\langle \unicode[STIX]{x1D70E}_{12}\rangle _{D}=\frac{\unicode[STIX]{x03C0}(D_{r}\unicode[STIX]{x1D70F})(nL^{3})}{5B\log \unicode[STIX]{x1D705}}[\hat{\dot{\unicode[STIX]{x1D6FE}}}-H(t-T_{1})\exp (-B(t-T_{1}))+H(t-T_{2})\exp (-B(t-T_{2}))].\end{eqnarray}$$
$$\begin{eqnarray}\langle \unicode[STIX]{x1D70E}_{12}\rangle _{D}=\frac{\unicode[STIX]{x03C0}(D_{r}\unicode[STIX]{x1D70F})(nL^{3})}{5B\log \unicode[STIX]{x1D705}}[\hat{\dot{\unicode[STIX]{x1D6FE}}}-H(t-T_{1})\exp (-B(t-T_{1}))+H(t-T_{2})\exp (-B(t-T_{2}))].\end{eqnarray}$$
Next, we determine the contribution to the stochastic stress due to the run-and-tumble dynamics. The tumbling operator in (2.1) is given as:
 $$\begin{eqnarray}\frac{1}{\unicode[STIX]{x1D70F}}\left(\unicode[STIX]{x1D6FA}-\int K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })\unicode[STIX]{x1D6FA}(\boldsymbol{p}^{\prime })\,\text{d}\boldsymbol{p}^{\prime }\right)=\frac{1}{\unicode[STIX]{x1D70F}}\int \,\text{d}\boldsymbol{p}^{\prime }(\unicode[STIX]{x1D739}(\boldsymbol{p}-\boldsymbol{p}^{\prime })-K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime }))\unicode[STIX]{x1D6FA}(\boldsymbol{p}^{\prime }).\end{eqnarray}$$
$$\begin{eqnarray}\frac{1}{\unicode[STIX]{x1D70F}}\left(\unicode[STIX]{x1D6FA}-\int K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })\unicode[STIX]{x1D6FA}(\boldsymbol{p}^{\prime })\,\text{d}\boldsymbol{p}^{\prime }\right)=\frac{1}{\unicode[STIX]{x1D70F}}\int \,\text{d}\boldsymbol{p}^{\prime }(\unicode[STIX]{x1D739}(\boldsymbol{p}-\boldsymbol{p}^{\prime })-K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime }))\unicode[STIX]{x1D6FA}(\boldsymbol{p}^{\prime }).\end{eqnarray}$$
In order to express the above contribution as a gradient in orientation space, and thereby, obtain an effective drift velocity, the kernel inside the integral is rewritten as:
 $$\begin{eqnarray}\unicode[STIX]{x1D739}(\boldsymbol{p}-\boldsymbol{p}^{\prime })-K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })=\unicode[STIX]{x1D6FB}_{p}^{2}H_{M}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime }).\end{eqnarray}$$
$$\begin{eqnarray}\unicode[STIX]{x1D739}(\boldsymbol{p}-\boldsymbol{p}^{\prime })-K(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })=\unicode[STIX]{x1D6FB}_{p}^{2}H_{M}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime }).\end{eqnarray}$$
Since the above is the orientational Laplacian forced by a function that integrates to zero over a unit sphere, one may obtain
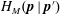 $H_{M}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })$
in terms of the modified Greens function,
$H_{M}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })$
in terms of the modified Greens function,
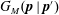 $G_{M}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })$
(Subramanian & Koch Reference Subramanian and Koch2009), as:
$G_{M}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })$
(Subramanian & Koch Reference Subramanian and Koch2009), as:
 $$\begin{eqnarray}\displaystyle H_{M}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })=\int \,\text{d}\boldsymbol{p}^{\prime \prime }G_{M}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime \prime })[\unicode[STIX]{x1D739}(\boldsymbol{p}^{\prime \prime }-\boldsymbol{p}^{\prime })-K(\boldsymbol{p}^{\prime \prime }\mid \boldsymbol{p}^{\prime })], & & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle H_{M}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })=\int \,\text{d}\boldsymbol{p}^{\prime \prime }G_{M}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime \prime })[\unicode[STIX]{x1D739}(\boldsymbol{p}^{\prime \prime }-\boldsymbol{p}^{\prime })-K(\boldsymbol{p}^{\prime \prime }\mid \boldsymbol{p}^{\prime })], & & \displaystyle\end{eqnarray}$$
where
 $$\begin{eqnarray}G_{M}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })=-\frac{1}{4\unicode[STIX]{x03C0}}\mathop{\sum }_{n=1}^{\infty }\frac{(2n+1)}{n(n+1)}P_{n}(\boldsymbol{p}\boldsymbol{\cdot }\boldsymbol{p}^{\prime }).\end{eqnarray}$$
$$\begin{eqnarray}G_{M}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })=-\frac{1}{4\unicode[STIX]{x03C0}}\mathop{\sum }_{n=1}^{\infty }\frac{(2n+1)}{n(n+1)}P_{n}(\boldsymbol{p}\boldsymbol{\cdot }\boldsymbol{p}^{\prime }).\end{eqnarray}$$
The tumbling operator now takes the form:
 $$\begin{eqnarray}\displaystyle \unicode[STIX]{x1D735}_{\boldsymbol{p}}\boldsymbol{\cdot }\left[\frac{1}{\unicode[STIX]{x1D70F}}\int \,\text{d}\boldsymbol{p}^{\prime }\unicode[STIX]{x1D735}_{\boldsymbol{p}}(H_{M}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })\unicode[STIX]{x1D6FA}(\boldsymbol{p}^{\prime }))\right]=\unicode[STIX]{x1D735}_{\boldsymbol{ p}}\boldsymbol{\cdot }[\dot{\boldsymbol{p}}_{T}\{\unicode[STIX]{x1D6FA}\}], & & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle \unicode[STIX]{x1D735}_{\boldsymbol{p}}\boldsymbol{\cdot }\left[\frac{1}{\unicode[STIX]{x1D70F}}\int \,\text{d}\boldsymbol{p}^{\prime }\unicode[STIX]{x1D735}_{\boldsymbol{p}}(H_{M}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })\unicode[STIX]{x1D6FA}(\boldsymbol{p}^{\prime }))\right]=\unicode[STIX]{x1D735}_{\boldsymbol{ p}}\boldsymbol{\cdot }[\dot{\boldsymbol{p}}_{T}\{\unicode[STIX]{x1D6FA}\}], & & \displaystyle\end{eqnarray}$$
where
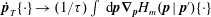 $\dot{\boldsymbol{p}}_{T}\{\cdot \}\rightarrow (1/\unicode[STIX]{x1D70F})\int \,\text{d}\boldsymbol{p}\unicode[STIX]{x1D735}_{\boldsymbol{p}}H_{m}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })\{\cdot \}$
is identified as the tumbling drift operator. The transverse linear force density operator due to tumbling (
$\dot{\boldsymbol{p}}_{T}\{\cdot \}\rightarrow (1/\unicode[STIX]{x1D70F})\int \,\text{d}\boldsymbol{p}\unicode[STIX]{x1D735}_{\boldsymbol{p}}H_{m}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })\{\cdot \}$
is identified as the tumbling drift operator. The transverse linear force density operator due to tumbling (
 $\boldsymbol{f}^{T}$
) is then given as:
$\boldsymbol{f}^{T}$
) is then given as:
 $$\begin{eqnarray}\boldsymbol{f}^{T}(s,\boldsymbol{\cdot })=\frac{4\unicode[STIX]{x03C0}\unicode[STIX]{x1D707}s}{\log \unicode[STIX]{x1D705}}(\unicode[STIX]{x1D644}-\boldsymbol{p}\boldsymbol{p})\boldsymbol{\cdot }\dot{\boldsymbol{p}}_{T}(\boldsymbol{\cdot }),\end{eqnarray}$$
$$\begin{eqnarray}\boldsymbol{f}^{T}(s,\boldsymbol{\cdot })=\frac{4\unicode[STIX]{x03C0}\unicode[STIX]{x1D707}s}{\log \unicode[STIX]{x1D705}}(\unicode[STIX]{x1D644}-\boldsymbol{p}\boldsymbol{p})\boldsymbol{\cdot }\dot{\boldsymbol{p}}_{T}(\boldsymbol{\cdot }),\end{eqnarray}$$
with
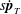 $s\dot{\boldsymbol{p}}_{T}$
being the tangential velocity (operator) due to tumbling. Similar to the diffusion case above, the tumbling stress is given by:
$s\dot{\boldsymbol{p}}_{T}$
being the tangential velocity (operator) due to tumbling. Similar to the diffusion case above, the tumbling stress is given by:
 $$\begin{eqnarray}\displaystyle \langle \unicode[STIX]{x1D748}\rangle _{T}=-\frac{n}{2}\int \,\text{d}\boldsymbol{p}\int _{-L/2}^{L/2}s[\boldsymbol{f}^{T}(s,\unicode[STIX]{x1D6FA})\boldsymbol{p}+\boldsymbol{p}\boldsymbol{f}^{T}(s,\unicode[STIX]{x1D6FA})]\,\text{d}s, & & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle \langle \unicode[STIX]{x1D748}\rangle _{T}=-\frac{n}{2}\int \,\text{d}\boldsymbol{p}\int _{-L/2}^{L/2}s[\boldsymbol{f}^{T}(s,\unicode[STIX]{x1D6FA})\boldsymbol{p}+\boldsymbol{p}\boldsymbol{f}^{T}(s,\unicode[STIX]{x1D6FA})]\,\text{d}s, & & \displaystyle\end{eqnarray}$$
where the drift operator acts on the orientation probability density
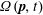 $\unicode[STIX]{x1D6FA}(\boldsymbol{p},t)$
, consistent with the interpretation of the tumbling term in the original Smoluchowski equation. We now use (A 10), (A 12) and (A 13) in the above to obtain:
$\unicode[STIX]{x1D6FA}(\boldsymbol{p},t)$
, consistent with the interpretation of the tumbling term in the original Smoluchowski equation. We now use (A 10), (A 12) and (A 13) in the above to obtain:
 $$\begin{eqnarray}\displaystyle \langle \unicode[STIX]{x1D70E}_{ij}\rangle _{T} & = & \displaystyle -\frac{\unicode[STIX]{x03C0}\unicode[STIX]{x1D707}(nL^{3})}{6\unicode[STIX]{x1D70F}\log \unicode[STIX]{x1D705}}\int \,\text{d}\boldsymbol{p}\int \,\text{d}\boldsymbol{p}^{\prime }\left\{\frac{\unicode[STIX]{x2202}}{\unicode[STIX]{x2202}p_{i}}\left[G_{M}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })-\left(\int G_{M}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime \prime })K(\boldsymbol{p}^{\prime \prime }\mid \boldsymbol{p}^{\prime })\unicode[STIX]{x1D6FA}(\boldsymbol{p}^{\prime })\,\text{d}\boldsymbol{p}^{\prime \prime }\right)\right]p_{j}\right.\nonumber\\ \displaystyle & & \displaystyle \left.+\,\frac{\unicode[STIX]{x2202}}{\unicode[STIX]{x2202}p_{j}}\left[G_{M}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })-\left(\int G_{M}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime \prime })K(\boldsymbol{p}^{\prime \prime }\mid \boldsymbol{p}^{\prime })\unicode[STIX]{x1D6FA}(\boldsymbol{p}^{\prime })\,\text{d}\boldsymbol{p}^{\prime \prime }\right)\right]p_{i}\right\}.\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle \langle \unicode[STIX]{x1D70E}_{ij}\rangle _{T} & = & \displaystyle -\frac{\unicode[STIX]{x03C0}\unicode[STIX]{x1D707}(nL^{3})}{6\unicode[STIX]{x1D70F}\log \unicode[STIX]{x1D705}}\int \,\text{d}\boldsymbol{p}\int \,\text{d}\boldsymbol{p}^{\prime }\left\{\frac{\unicode[STIX]{x2202}}{\unicode[STIX]{x2202}p_{i}}\left[G_{M}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })-\left(\int G_{M}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime \prime })K(\boldsymbol{p}^{\prime \prime }\mid \boldsymbol{p}^{\prime })\unicode[STIX]{x1D6FA}(\boldsymbol{p}^{\prime })\,\text{d}\boldsymbol{p}^{\prime \prime }\right)\right]p_{j}\right.\nonumber\\ \displaystyle & & \displaystyle \left.+\,\frac{\unicode[STIX]{x2202}}{\unicode[STIX]{x2202}p_{j}}\left[G_{M}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime })-\left(\int G_{M}(\boldsymbol{p}\mid \boldsymbol{p}^{\prime \prime })K(\boldsymbol{p}^{\prime \prime }\mid \boldsymbol{p}^{\prime })\unicode[STIX]{x1D6FA}(\boldsymbol{p}^{\prime })\,\text{d}\boldsymbol{p}^{\prime \prime }\right)\right]p_{i}\right\}.\end{eqnarray}$$
Again, the deviatoric stochastic tumbling stress arises from first-order anisotropy in the orientation probability density (
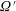 $\unicode[STIX]{x1D6FA}^{\prime }$
), and after some manipulation, one finally obtains:
$\unicode[STIX]{x1D6FA}^{\prime }$
), and after some manipulation, one finally obtains:
 $$\begin{eqnarray}\langle \unicode[STIX]{x1D70E}_{12}\rangle _{T}=\frac{\unicode[STIX]{x03C0}(nL^{3})G(\unicode[STIX]{x1D6FD})}{30B\log \unicode[STIX]{x1D705}}[\hat{\dot{\unicode[STIX]{x1D6FE}}}-H(t-T_{1})\exp (-B(t-T_{1}))+H(t-T_{2})\exp (-B(t-T_{2}))],\end{eqnarray}$$
$$\begin{eqnarray}\langle \unicode[STIX]{x1D70E}_{12}\rangle _{T}=\frac{\unicode[STIX]{x03C0}(nL^{3})G(\unicode[STIX]{x1D6FD})}{30B\log \unicode[STIX]{x1D705}}[\hat{\dot{\unicode[STIX]{x1D6FE}}}-H(t-T_{1})\exp (-B(t-T_{1}))+H(t-T_{2})\exp (-B(t-T_{2}))],\end{eqnarray}$$
where,
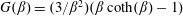 $G(\unicode[STIX]{x1D6FD})=(3/\unicode[STIX]{x1D6FD}^{2})(\unicode[STIX]{x1D6FD}\coth (\unicode[STIX]{x1D6FD})-1)$
. We note that the time dependence in (A 7) and (A 16) arises from
$G(\unicode[STIX]{x1D6FD})=(3/\unicode[STIX]{x1D6FD}^{2})(\unicode[STIX]{x1D6FD}\coth (\unicode[STIX]{x1D6FD})-1)$
. We note that the time dependence in (A 7) and (A 16) arises from
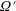 $\unicode[STIX]{x1D6FA}^{\prime }$
and is therefore same as that obtained for the active hydrodynamic stress. This is important since it ensures that the inclusion of an active stochastic stress does not qualitatively alter the stress relaxation obtained in the main paper.
$\unicode[STIX]{x1D6FA}^{\prime }$
and is therefore same as that obtained for the active hydrodynamic stress. This is important since it ensures that the inclusion of an active stochastic stress does not qualitatively alter the stress relaxation obtained in the main paper.
Summing the two contributions, we get the active stochastic stress, due to the combined effects of rotary diffusion and run-and-tumble dynamics, to be:
 $$\begin{eqnarray}\langle \unicode[STIX]{x1D70E}_{12}\rangle _{st}=\frac{\unicode[STIX]{x03C0}(nL^{3})}{30\log \unicode[STIX]{x1D705}}[\hat{\dot{\unicode[STIX]{x1D6FE}}}-H(t-T_{1})\exp (-B(t-T_{1}))+H(t-T_{2})\exp (-B(t-T_{2}))],\end{eqnarray}$$
$$\begin{eqnarray}\langle \unicode[STIX]{x1D70E}_{12}\rangle _{st}=\frac{\unicode[STIX]{x03C0}(nL^{3})}{30\log \unicode[STIX]{x1D705}}[\hat{\dot{\unicode[STIX]{x1D6FE}}}-H(t-T_{1})\exp (-B(t-T_{1}))+H(t-T_{2})\exp (-B(t-T_{2}))],\end{eqnarray}$$
which, interestingly, is independent of both
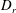 $D_{r}$
and
$D_{r}$
and
 $\unicode[STIX]{x1D70F}$
. In other words, the stochastic stress estimate at leading order appears independent of the precise mechanism of orientation relaxation to isotropy.
$\unicode[STIX]{x1D70F}$
. In other words, the stochastic stress estimate at leading order appears independent of the precise mechanism of orientation relaxation to isotropy.
It is worth mentioning that the stochastic stress above is a pure diffusion stress for
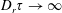 $D_{r}\unicode[STIX]{x1D70F}\rightarrow \infty$
, and is three times the hydrodynamic stresslet in the leading (logarithmic) order slender body theory considered here. This is, of course, a well-known result for a passive suspension of slender fibres where the direct contribution due to the diffusion (Brownian) stress is the dominant (three-quarters) contribution to the zero-shear viscosity. However, the estimate of the stochastic stress above remains substantially smaller than the active hydrodynamic stress that arises due to the anisotropy of the intrinsic force dipoles. This ratio is determined in the next subsection.
$D_{r}\unicode[STIX]{x1D70F}\rightarrow \infty$
, and is three times the hydrodynamic stresslet in the leading (logarithmic) order slender body theory considered here. This is, of course, a well-known result for a passive suspension of slender fibres where the direct contribution due to the diffusion (Brownian) stress is the dominant (three-quarters) contribution to the zero-shear viscosity. However, the estimate of the stochastic stress above remains substantially smaller than the active hydrodynamic stress that arises due to the anisotropy of the intrinsic force dipoles. This ratio is determined in the next subsection.
A.2 Comparison of stochastic stresses with the hydrodynamic stress
We begin by summarizing the individual contributions, from rotary diffusion and run-and-tumble dynamics, to the steady-state stochastic stress as evaluated in § A.1:
 $$\begin{eqnarray}\displaystyle \left.\begin{array}{@{}c@{}}\langle \unicode[STIX]{x1D70E}_{12}\rangle _{D}={\displaystyle \frac{\unicode[STIX]{x03C0}(D_{r}\unicode[STIX]{x1D70F})(nL^{3})}{5B\log \unicode[STIX]{x1D705}}},\\ \langle \unicode[STIX]{x1D70E}_{12}\rangle _{T}={\displaystyle \frac{\unicode[STIX]{x03C0}(nL^{3})G(\unicode[STIX]{x1D6FD})}{30B\log \unicode[STIX]{x1D705}}}.\end{array}\right\} & & \displaystyle\end{eqnarray}$$
$$\begin{eqnarray}\displaystyle \left.\begin{array}{@{}c@{}}\langle \unicode[STIX]{x1D70E}_{12}\rangle _{D}={\displaystyle \frac{\unicode[STIX]{x03C0}(D_{r}\unicode[STIX]{x1D70F})(nL^{3})}{5B\log \unicode[STIX]{x1D705}}},\\ \langle \unicode[STIX]{x1D70E}_{12}\rangle _{T}={\displaystyle \frac{\unicode[STIX]{x03C0}(nL^{3})G(\unicode[STIX]{x1D6FD})}{30B\log \unicode[STIX]{x1D705}}}.\end{array}\right\} & & \displaystyle\end{eqnarray}$$
The ratio of the magnitude of the total stochastic stress to the total hydrodynamic stress (active
 $+$
passive) is given as:
$+$
passive) is given as:
 $$\begin{eqnarray}\frac{|\langle \unicode[STIX]{x1D70E}_{12}\rangle _{D}|+|\langle \unicode[STIX]{x1D70E}_{12}\rangle _{T}|}{|\langle \unicode[STIX]{x1D70E}_{12}\rangle _{Passive}|+|\langle \unicode[STIX]{x1D70E}_{12}\rangle _{Active}|}=\frac{3\unicode[STIX]{x03C0}M}{\unicode[STIX]{x03C0}M+9\unicode[STIX]{x1D6FC}\log \unicode[STIX]{x1D705}\left({\displaystyle \frac{U}{L}}\right)(B^{-1}\unicode[STIX]{x1D70F})}.\end{eqnarray}$$
$$\begin{eqnarray}\frac{|\langle \unicode[STIX]{x1D70E}_{12}\rangle _{D}|+|\langle \unicode[STIX]{x1D70E}_{12}\rangle _{T}|}{|\langle \unicode[STIX]{x1D70E}_{12}\rangle _{Passive}|+|\langle \unicode[STIX]{x1D70E}_{12}\rangle _{Active}|}=\frac{3\unicode[STIX]{x03C0}M}{\unicode[STIX]{x03C0}M+9\unicode[STIX]{x1D6FC}\log \unicode[STIX]{x1D705}\left({\displaystyle \frac{U}{L}}\right)(B^{-1}\unicode[STIX]{x1D70F})}.\end{eqnarray}$$
Note that
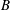 $B$
, which is the scale factor for the relaxation time (
$B$
, which is the scale factor for the relaxation time (
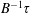 $B^{-1}\unicode[STIX]{x1D70F}$
), is a function of
$B^{-1}\unicode[STIX]{x1D70F}$
), is a function of
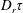 $D_{r}\unicode[STIX]{x1D70F}$
and the correlation parameter
$D_{r}\unicode[STIX]{x1D70F}$
and the correlation parameter
 $\unicode[STIX]{x1D6FD}$
. The active hydrodynamic stress involves two time scales: one corresponding to the time taken by the swimmer to swim a distance equal to its body length
$\unicode[STIX]{x1D6FD}$
. The active hydrodynamic stress involves two time scales: one corresponding to the time taken by the swimmer to swim a distance equal to its body length
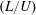 $(L/U)$
, and the other being
$(L/U)$
, and the other being
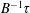 $B^{-1}\unicode[STIX]{x1D70F}$
. In order to determine the ratio of the stochastic to the hydrodynamic stresses, one needs an estimate of the ratio of these two time scales. For oxygen-deprived conditions,
$B^{-1}\unicode[STIX]{x1D70F}$
. In order to determine the ratio of the stochastic to the hydrodynamic stresses, one needs an estimate of the ratio of these two time scales. For oxygen-deprived conditions,
 $U\sim 20~\unicode[STIX]{x03BC}\text{m}~\text{s}^{-1}$
,
$U\sim 20~\unicode[STIX]{x03BC}\text{m}~\text{s}^{-1}$
,
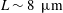 $L\sim 8~\unicode[STIX]{x03BC}\text{m}$
,
$L\sim 8~\unicode[STIX]{x03BC}\text{m}$
,
 $B^{-1}\unicode[STIX]{x1D70F}\sim 0.8~\text{s}$
, and we have
$B^{-1}\unicode[STIX]{x1D70F}\sim 0.8~\text{s}$
, and we have
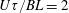 $U\unicode[STIX]{x1D70F}/BL=2$
, as a reasonable estimate. On the other hand, for the large relaxation times (
$U\unicode[STIX]{x1D70F}/BL=2$
, as a reasonable estimate. On the other hand, for the large relaxation times (
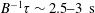 $B^{-1}\unicode[STIX]{x1D70F}\sim 2.5{-}3~\text{s}$
) corresponding to oxygen-rich conditions, we have
$B^{-1}\unicode[STIX]{x1D70F}\sim 2.5{-}3~\text{s}$
) corresponding to oxygen-rich conditions, we have
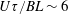 $U\unicode[STIX]{x1D70F}/BL\sim 6$
.
$U\unicode[STIX]{x1D70F}/BL\sim 6$
.
In figure 7, we plot the aforementioned ratio (A 19), as also the ratios of the individual contributions, to the total hydrodynamic stress as a function of
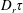 $D_{r}\unicode[STIX]{x1D70F}$
. This is done for the
$D_{r}\unicode[STIX]{x1D70F}$
. This is done for the
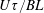 $U\unicode[STIX]{x1D70F}/BL$
values, corresponding to oxygen-deprived and oxygen-rich conditions, obtained above (
$U\unicode[STIX]{x1D70F}/BL$
values, corresponding to oxygen-deprived and oxygen-rich conditions, obtained above (
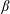 $\unicode[STIX]{x1D6FD}$
is set to unity as is the case for E. coli; no significant change is observed for
$\unicode[STIX]{x1D6FD}$
is set to unity as is the case for E. coli; no significant change is observed for
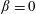 $\unicode[STIX]{x1D6FD}=0$
corresponding to random tumbling.). The total stochastic stress is, of course, independent of
$\unicode[STIX]{x1D6FD}=0$
corresponding to random tumbling.). The total stochastic stress is, of course, independent of
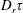 $D_{r}\unicode[STIX]{x1D70F}$
, and less than
$D_{r}\unicode[STIX]{x1D70F}$
, and less than
 $20\,\%$
of the total hydrodynamic stress. This then serves as an upper bound for the stress contribution arising from the torque-free orientation fluctuations for an actual bacterium.
$20\,\%$
of the total hydrodynamic stress. This then serves as an upper bound for the stress contribution arising from the torque-free orientation fluctuations for an actual bacterium.

Figure 7. The ratios of the total stochastic stress – red (solid), the diffusion stress component – blue (dotted) and the run-and-tumble stress component – green (dash-dotted) to the total hydrodynamic stress, plotted as a function of
 $D_{r}\unicode[STIX]{x1D70F}$
. Panel (a) is for
$D_{r}\unicode[STIX]{x1D70F}$
. Panel (a) is for
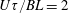 $U\unicode[STIX]{x1D70F}/BL=2$
, corresponding to oxygen-deprived conditions, and (b) is for
$U\unicode[STIX]{x1D70F}/BL=2$
, corresponding to oxygen-deprived conditions, and (b) is for
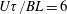 $U\unicode[STIX]{x1D70F}/BL=6$
, corresponding to oxygen-rich conditions.
$U\unicode[STIX]{x1D70F}/BL=6$
, corresponding to oxygen-rich conditions.
A.3 Inclusion of stochastic stress in total stress
Having concluded above that the stochastic stresses lead to a marginal change in the stress magnitude, we now look at the stress relaxation that results from the inclusion of these stresses. Figure 8 shows the deviatoric stress as a function of time for the same set of parameters as in figure 2 (
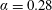 $\unicode[STIX]{x1D6FC}=0.28$
), both with and without the stochastic stresses. As expected, the inclusion of the stochastic stress does not qualitatively alter the relaxation, and only leads to the total stress relaxing to a slightly higher plateau, which in turn decreases slightly the peak negative stress following shear cessation.
$\unicode[STIX]{x1D6FC}=0.28$
), both with and without the stochastic stresses. As expected, the inclusion of the stochastic stress does not qualitatively alter the relaxation, and only leads to the total stress relaxing to a slightly higher plateau, which in turn decreases slightly the peak negative stress following shear cessation.
Finally, it is worth reiterating that the manner in which the stochastic stress has been obtained above does not account for the torque-free nature of the stochastic reorientations. The stress given by (A 17) applies, in fact, to a suspension of slender rods that reorient due to continuous random torques with an amplitude proportional to
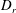 $D_{r}$
and impulsive random torques, acting at time instants that are Poisson-distributed, with an amplitude proportional to
$D_{r}$
and impulsive random torques, acting at time instants that are Poisson-distributed, with an amplitude proportional to
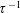 $\unicode[STIX]{x1D70F}^{-1}$
. As explained in the main paper, and mentioned above, (A 17) serves as an upper bound in the actual torque-free case. Within the framework of a simple slender-rod approximation considered here, a torque-free reorientation would actually imply a zero stochastic stress. Thus, the minimal swimmer model that allows an actual calculation of a torque-free stress contribution would have to go beyond the slender-rod geometry. This in turn might lead to significantly greater complexity, since one would need to potentially account for the intricate sequence of configurational changes in the flagellar bundle that lead to orientation changes in an actual swimming bacterium (Turner, Ryu & Berg Reference Turner, Ryu and Berg2000; Berg Reference Berg2004; Darnton et al.
Reference Darnton, Turner, Rojevsky and Berg2007).
$\unicode[STIX]{x1D70F}^{-1}$
. As explained in the main paper, and mentioned above, (A 17) serves as an upper bound in the actual torque-free case. Within the framework of a simple slender-rod approximation considered here, a torque-free reorientation would actually imply a zero stochastic stress. Thus, the minimal swimmer model that allows an actual calculation of a torque-free stress contribution would have to go beyond the slender-rod geometry. This in turn might lead to significantly greater complexity, since one would need to potentially account for the intricate sequence of configurational changes in the flagellar bundle that lead to orientation changes in an actual swimming bacterium (Turner, Ryu & Berg Reference Turner, Ryu and Berg2000; Berg Reference Berg2004; Darnton et al.
Reference Darnton, Turner, Rojevsky and Berg2007).

Figure 8. The stress relaxation curves with – red (dash-dotted) and without – green (solid) the stochastic stress contribution with parameter values chosen corresponding to figure 2.







