



Introduction
The genus Longidorus Micoletzky, Reference Micoletzky1922, known as needle nematode, comprises a specious genus and belongs to the family Longidoridae Thorne, Reference Thorne1935. It currently contains around 185 valid species (Gharibzadeh et al. Reference Gharibzadeh, Pourjam and Pedram2018; Bakhshi Amrei et al. Reference Bakhshi Amrei, Peneva, Rakhshandehroo and Pedram2020, Reference Bakhshi Amrei, Peneva, Rakhshandehroo and Pedram2022; Liébanas et al. Reference Liébanas, Clavero-Camacho, Cantalapiedra-Navarrete, Guerrero, Palomares-Rius, Castillo and Archidona-Yuste2022; Mobasseri et al. Reference Mobasseri, Pourjam, Farshiani and Pedram2022; Pour Ehteshan et al. Reference Pour Ehtesham, Pedram, Atighi and Jahanshahi Afshar2023). They are polyphagous root ectoparasites, feeding on a variety of woody and herbaceous plants. They directly feed on root cells and arrest root tip growth and sometimes, form gall on root tips (Wyss Reference Wyss and Lee2002). Some species, however, transmit plant-pathogenic nepoviruses (Taylor and Brown Reference Taylor and Brown1997; Decraemer and Robbins Reference Decraemer and Robbins2007; Cai et al. Reference Cai, Archidona-Yuste, Cantalapiedra-Navarrete, Palomares-Rius and Castillo2020a).
Integrative diagnosis of Longidorus species, combining morphological and molecular data, is largely used for taxonomic studies of longidorids by nematologists (Gharibzadeh et al. Reference Gharibzadeh, Pourjam and Pedram2018; Cai et al. Reference Cai, Archidona-Yuste, Cantalapiedra-Navarrete, Palomares-Rius and Castillo2020a, Reference Cai, Archidona‐Yuste, Cantalapiedra‐Navarrete, Palomares‐Rius and Castillob; Bakhshi Amrei et al. Reference Bakhshi Amrei, Peneva, Rakhshandehroo and Pedram2020, Reference Bakhshi Amrei, Peneva, Rakhshandehroo and Pedram2022; Liébanas et al. Reference Liébanas, Clavero-Camacho, Cantalapiedra-Navarrete, Guerrero, Palomares-Rius, Castillo and Archidona-Yuste2022; Mobasseri et al. Reference Mobasseri, Pourjam, Farshiani and Pedram2022). In addition to 21 Longidorus species occurring in Iran (Pedram Reference Pedram, Ghaderi, Kashi and Karegar2018), the species L. azarbaijanensis Gharibzadeh, Pourjam & Pedram, Reference Gharibzadeh, Pourjam and Pedram2018, L. behshahrensis Bakhshi Amrei, Peneva, Rakhshandehroo & Pedram, Reference Bakhshi Amrei, Peneva, Rakhshandehroo and Pedram2020, L. armeniacae Bakhshi Amrei, Peneva, Rakhshandehroo & Pedram, Reference Bakhshi Amrei, Peneva, Rakhshandehroo and Pedram2022, L. sabalanicus Asgari, Eskandari, Castillo & Palomares-Rius, Reference Asgari, Eskandari, Castillo and Palomares-Rius2022, L. hyrcanus Mobasseri, Pourjam, Farashiani & Pedram, Reference Mobasseri, Pourjam, Farshiani and Pedram2022, and L. soosanae Pour Ehtesham, Pedram, Atighi & Jahanshahi Afshar, Reference Pour Ehtesham, Pedram, Atighi and Jahanshahi Afshar2023, have been described from the country.
In the present study, the characteristics of four Longidorus species from southern and southeastern Iran will be discussed.
Materials and methods
Soil samplings, nematode isolation, and morphological studies
A total of 40 soil samples were collected from around the roots of different plants in Southern Khorasan and Bushehr provinces. The longidorid specimens were extracted from soil using two 20 and 60 mesh sieves (with openings equal to 850 μm and 250 μm) and handpicked under a Nikon SMZ1000 (Nikon, Tokyo, Japan) stereomicroscope. The recovered specimens were heat-killed by adding hot 4% formalin solution, transferred to anhydrous glycerine according to De Grisse (Reference De Grisse1969), and mounted on permanent slides. The morphometrics were provided using Olympus BH2 (Olympus, Tokyo, Japan) and Nikon E600 (Nikon, Tokyo, Japan) light microscopes equipped with a drawing tube. Digital images of the fresh individuals and mounted specimens were taken using an Olympus DP72 digital camera (Olympus, Tokyo, Japan) attached to an Olympus BX51 microscope (Olympus, Tokyo, Japan) powered with differential interference contrast optics. Line drawings were made using the drawing tube and Corel DRAW® software (www.coreldraw.com) version 2020.
Molecular characterisation
For DNA extraction of newly recovered populations, a living adult female nematode of each population was selected, washed with sterile water, checked on a temporary slide, photographed, transferred to a drop of TE buffer (10 mM Tris-Cl, 0.5 mM EDTA; pH 9.0) on a clean slide, and squashed using a clean slide cover. The suspension was collected by adding 25 μl TE buffer and stored at −20°C to be used in polymerase chain reaction (PCR). The DNA sample of the type population of Longidorus tabrizicus Niknam, Pedram, Ghahremani Nejad, Ye, Robbins & Tanha Maafi, Reference Niknam, Pedram, Ghahremani Nejad, Ye, Robbins and Tanha Maafi2010 was used for its LSU D2-D3 amplification/sequencing. The small-subunit (SSU) rDNA was amplified using two primer pairs: forward 988F (5′-CTCAAAGATTAAGCCATGC-3′) and reverse 1912R (5′-TTTACGGTCAGAACTAGGG-3′); and forward 1813F (5′-CTGCGTGAGAGGTGAAAT-3′) and reverse 2646R (5′-GCTACCTTGTTACGACTTTT-3′) (Holterman et al. Reference Holterman, Van Der Wurff, van Den Elsen, van Megen, Bongers, Holovachov, Bakker and Helder2006). The D2-D3 expansion segments of large-subunit (LSU) rDNA were amplified using primers forward D2A (5′-ACAAGTACCGTGAGGGAAAGTTG-3′) and reverse D3B (5′-TCGGAAGGAACCAGCTACTA-3′) (Nunn Reference Holterman, Van Der Wurff, van Den Elsen, van Megen, Bongers, Holovachov, Bakker and Helder1992). The internal transcribed spacer 1 region (ITS1) was amplified using two primer pairs: forward rDNA1 (5′-TTGATTACGTCCCTGCCCTTT-3′) and reverse rDNA2 (5′-TTTCACTCGCCGTTACTAAGG-3′) (Subbotin et al. Reference Subbotin, Halford, Warry and Perry2000); forward TW81 (5′-GTTTCCGTAGGTGAACCTGC-3′) and reverse AB28 (5′-ATATGCTTAAGTTCAGCGGGT-3′) (Joyce et al. Reference Joyce, Reid, Driver, Curran, Burnell, Ehlers and Masson1994). The PCR mixture (30 ml) contained the following: 15 μl Taq DNA polymerase 2× Master Mix RED, 2mM MgCl2 (Ampliqon, Odenese, Denmark), 8 μl distilled water, 1 μl of each primer (10 pmlo/μL), and 5 μl of DNA template. The PCR to amplify three genomic loci was performed with the following cycles: one cycle of 94°C for 5 min, followed by 35 cycles of 94°C for 30s, annealing temperature of 52°C for 1 min, 72°C for 1 min, and finally one cycle of 72°C for 10 min. The successfully amplified loci were sequenced using Applied Biosystems 3500 (ABI) sequencer, Pishgam corporation, Tehran, Iran. The obtained sequences were deposited into the GenBank database under the accession numbers presented in Table 1.
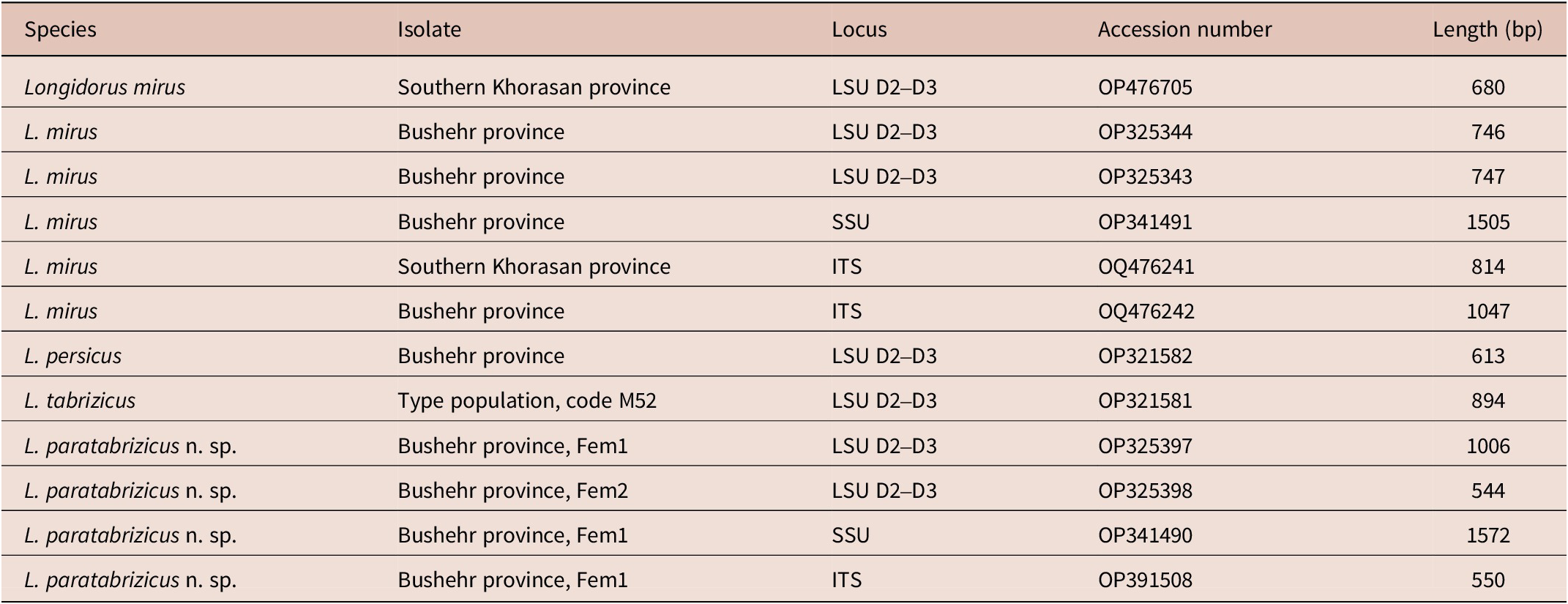
Table 1. Species, sequenced loci, accession numbers, and isolates names newly sequenced in present study
Phylogenetic analyses
The chromatograms of raw DNA sequences were checked using Chromas Lite 2.1.1 (http://technelysium.com.au/), manually edited/trimmed, and assembled when needed. The newly obtained SSU, LSU, and ITS sequences were compared with other available sequences in the GenBank database using the basic local alignment search tool (BLAST, https://blast.ncbi.nlm.nih.gov/Blast.cgi). Several sequences of representatives of the family Longidoridae were selected for SSU and LSU phylogenies. By having few available sequences having high coverage, newly generated ITS sequences were directly compared with other relevant sequences. Representatives of Xiphinema Cobb, Reference Cobb1913 were used as outgroups in SSU and LUS phylogenies. In total, 119 SSU and 130 LSU sequences (including newly generated sequences of the new species and Xiphinema sequences as outgroups) were included in each phylogeny, respectively. The LSU and SSU datasets were aligned using ClustalX2 (http://www.clustal.org/), and the resultant alignments were manually edited using MEGA6 (Tamura et al. Reference Tamura, Stecher, Peterson, Filipski and Kumar2013). The best-fitting substitution model for both datasets was selected using PAUP*/MrModeltest.2 (Nylander Reference Nylander2004). The Akaike-supported model, a general time-reversible model—including among-site rate heterogeneity and estimates of invariant sites (GTR + gamma [G] + invariant [I])—was selected and used in both phylogenies. Bayesian analyses were performed using MrBayes 3.1.2 (Ronquist and Huelsenbeck Reference Ronquist and Huelsenbeck2003), running the chains (two chains, as default) for five million generations for both datasets. After discarding burn-in samples, the remaining samples were retained for further analyses. The Markov chain Monte Carlo (MCMC) method within a Bayesian framework was used to estimate the posterior probabilities of the phylogenetic trees (Larget and Simon Reference Larget and Simon1999) using the 50% majority rule. Convergence of model parameters and topologies was assessed based on average standard deviation of split frequencies and potential scale reduction factor values. The adequacy of the posterior sample size was evaluated using autocorrelation statistics as implemented in Tracer v.1.6 (Rambaut and Drummond Reference Rambaut and Drummond2009). The output files of the trees were visualised using Dendroscope v3.2.8 (Huson and Scornavacca Reference Huson and Scornavacca2012), and trees were digitally drawn in CorelDRAW software version 2020.
Results
A population of Longidorus most closely resembling L. tabrizicus, herein named as L. paratabrizicus n. sp. was described. Three populations of L. mirus were recovered in association with narcissus and pistachio in southeastern Iran and date palm in southern Iran. One population of L. persicus was recovered in association with date palm in southern Iran. Several populations of L. orientalis with high population density were recovered from southern Iran in association with date palm.
Longidorus paratabrizicus n. sp.
Order: Dorylaimida; Suborder: Dorylaimina.
Figures 1 and 2 show line drawings and light microphotographs of L. paratabrizicus n. sp.

Figure 1. Longidorus paratabrizicus n. sp. (A) anterior body region; (B) Amphidial fovea; (C) pharynx; (D and E) pre-last and last juvenile developmental stages; (F) male posterior body region; (G and H) Female tail.

Figure 2. Light microphotographs of Longidorus paratabrizicus n. sp. (A) anterior body region; (B) vulval region; (C) male posterior body region; C1, spicule; (D and E) tail of pre-last and last juvenile developmental stages, respectively; (F) female tail (scale bar = 10 μm).
The morphometrics of L. paratabrizicus n. sp. are presented in Table 2.
Table 2. Morphometrics of Longidorus paratabrizicus n. sp. All measurements are in μm and in the form: mean ± S.D. (range)
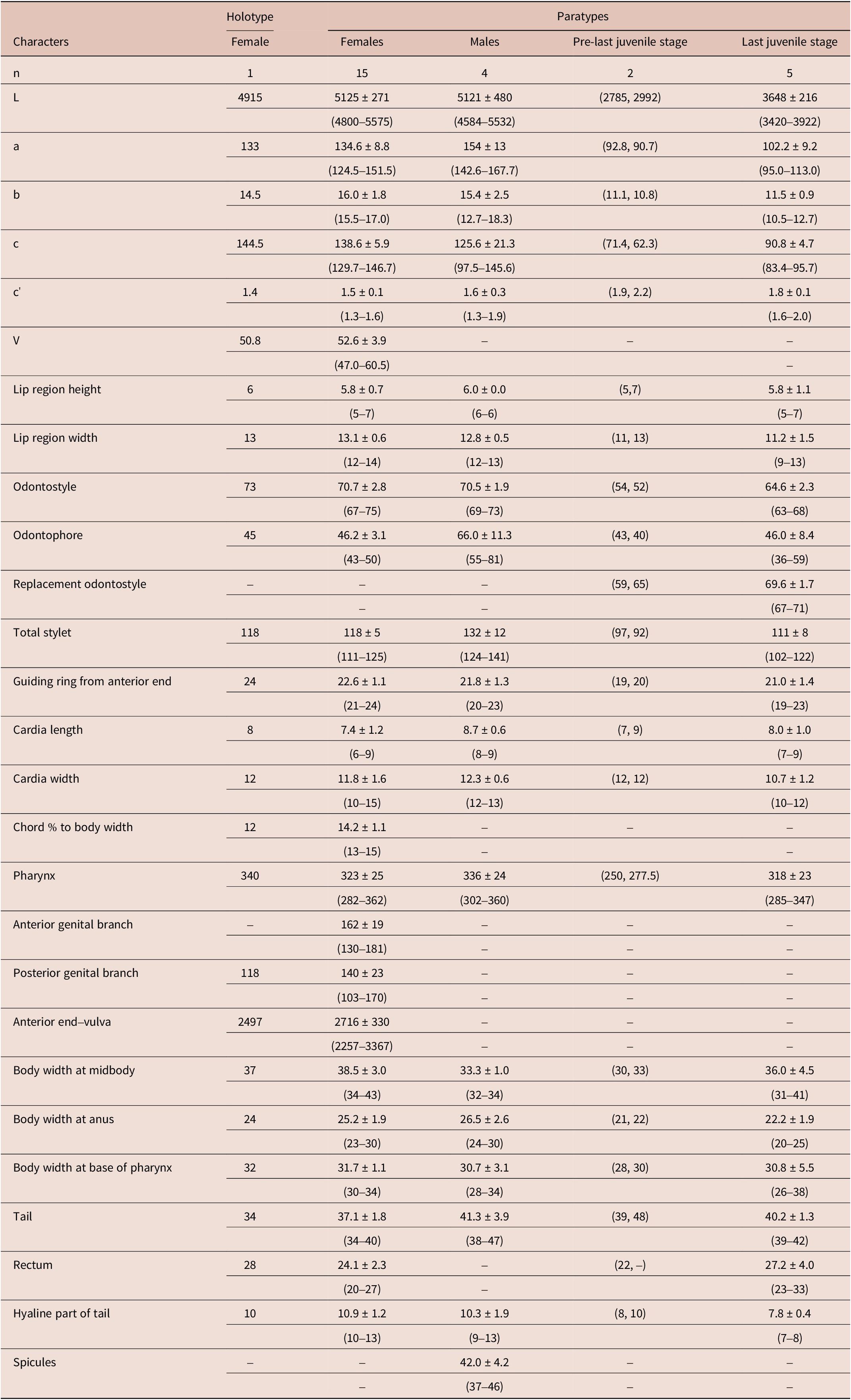
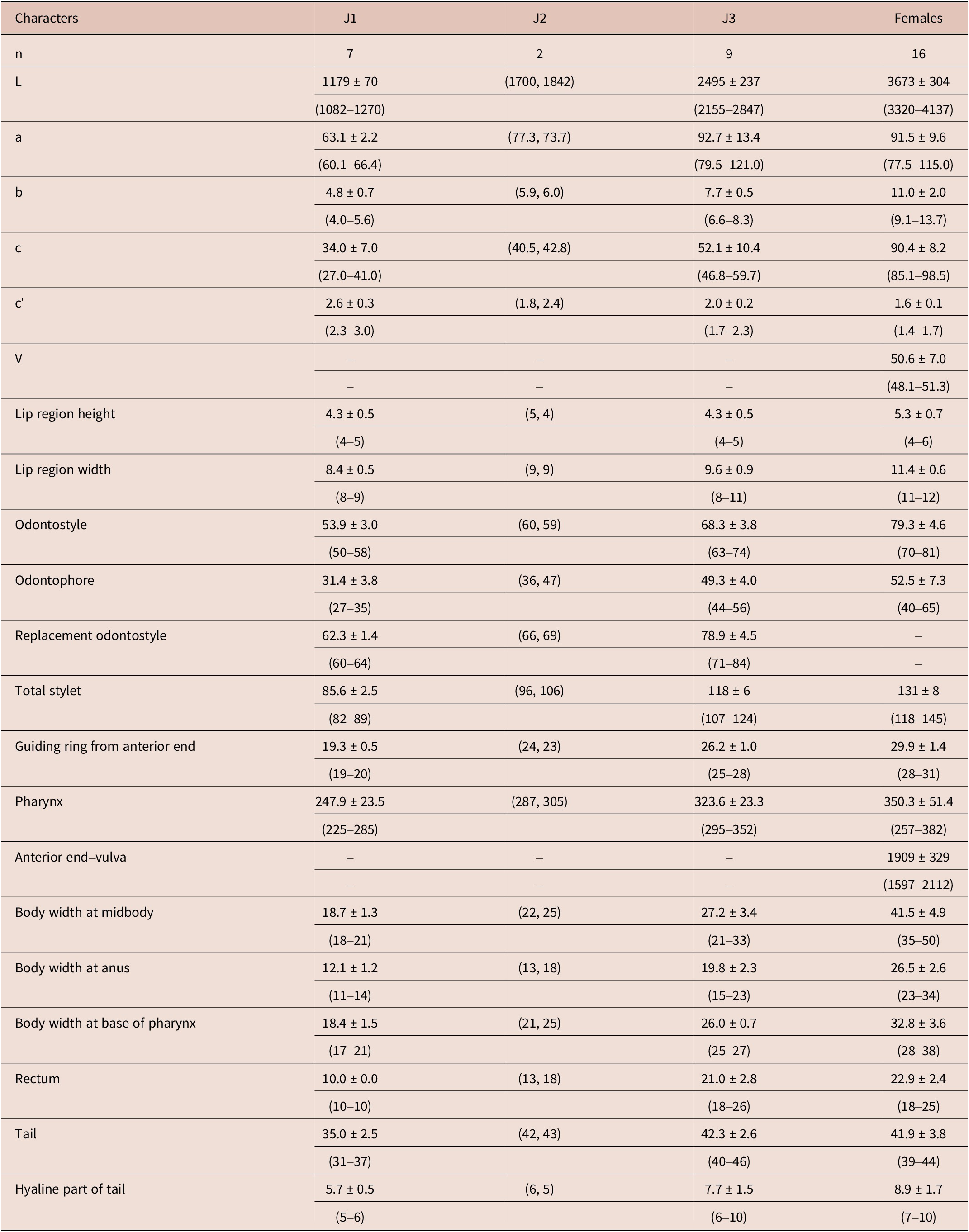
Table 3. Morphometrics of Longidorus mirus Khan, Chawla & Seshadri, Reference Khan, Chawla and Seshadri1971 from the rhizosphere of date palm from Bushehr province. All measurements are in μm and in the form: mean ± S.D. (range)
Description
Females
Cylindrical nematodes, assuming an open C after heat killing, tapering anteriorly. Cuticle two-layered, 1.9–2.4 μm thick at postlabial region, 2.2–4.2 μm thick at midbody, and 2.5–4.3 μm at the anus. Lateral chords 33–40% of corresponding body width. The lip region anteriorly flattened and separated from the rest of the body by depression. Amphidial fovea pocket-shaped without lobes. Odontostyle 1.1–1.8 times longer than odontophore, the latter simple, rod-like. Pharynx dorylaimoid, the pharyngeal bulb occupying 20–30% of the pharynx. The dorsal gland nucleus (DN) at 20–28% and two ventrosublateral nuclei (S1N) at 51–62%. Cardia semispherical. Reproductive system didelphic-amphidelphic, both branches almost equally developed, each composed of a reflexed ovary 111–198 μm long, oviduct 100–166 μm long, sphincter between the oviduct and tubular uterus, vagina perpendicular to body axis and vulva a transverse slit in ventral view. Prerectum 2.8–3.0 times and rectum 0.7–1.1 times anal body width long. Tail conical, dorsally convex, ventrally almost straight (or slightly concave), with narrow bluntly rounded tip.
Males
Similar to females in general morphology, except for the reproductive system and posterior body end more ventrally bent after fixation. The reproductive system composed of two opposed testes. Spicules dorylaimoid, massive, 8–14 μm wide, lateral accessory pieces 10.5–14.0 μm long. Tail conical, dorsally convex, ventrally concave. Two precloacal pairs of supplements preceded by a row of 8–11 ventromedian supplements in the ventral region, the two posteriormost supplements seen at different zoom.
Juveniles
Two last and pre-last stages of juveniles were recovered. The relation of body length and functional and replacement odontostyle of juveniles to the body length and odontostyle of females is given in Figure 3. Tail in both recovered stages conical, dorsally convex, with a narrow-rounded tip.

Figure 3. Scatter plot of the functional and replacement odontostyle length in relation to the body length of the juvenile stages and females of Longidorus paratabrizicus n. sp.
Etymology
The specific epithet “paratabrizicus” shows the new species most closely resembles Longidorus tabrizicus.
Type habitat and locality
Rhizosphere of date palm trees in Dashtestan region, Bushehr province, southern Iran. The GPS information of the sampling location is as follows: N29°21.322′, E51°15.779′.
Type material. Holotype female, four paratype females, and four paratype males were deposited at WaNeCo collection, Wageningen, the Netherlands (http://www.waneco.eu/). Ten paratype females and paratype juveniles were deposited at the Nematode Collection of the Faculty of Agriculture, Tarbiat Modares University, Tehran, Iran. The LSID code of this publication is: urn:lsid:zoobank.org:pub:4F79E12C-DA8A-420D-85E5-65C198310C71.
Diagnosis and relationships
Longidorus paratabrizicus n. sp. is characterised by 4.8–5.6 mm long females having anteriorly flattened lip region separated from rest of body by depression, amphidial fovea pocket-shaped without lobes, odontostyle 67–75 μm long, and odontophore 43–50 μm long, vulva at 47.0–60.5% of body length, tail conical, dorsally convex, ventrally almost straight with bluntly rounded tip (c = 129.7–146.7, c´ = 1.3–1.6), and males in population. The number of juvenile developmental stages of the new species was not determined as only two last and pre-last stages of juveniles were recovered. The polytomous identification codes of the new species according to Chen et al. (Reference Chen, Hooper, Loof and Xu1997) and Peneva et al. (Reference Peneva, Lazarova, De Luca and Brown2013), are as follows: A2, B2, C2, D3, E1, F23, G3, H23, I2, J?, K?. By having similar polytomous codes resulting from close morphology and close morphometrics, mainly similar lip region, and similar general shape of the tail, the new species most closely resembles five species viz. L. artemisiae Rubtsova, Chizhov & Subbotin, Reference Rubtsova, Chizhov and Subbotin1999, L. globulicauda Dalmasso, Reference Dalmasso1969, L. patuxentensis Kantor, Subbotin, Im & Handoo, Reference Kantor, Subbotin, Im and Handoo2024, L. sturhani Rubtsova, Subbotin, Brown & Moens, Reference Rubtsova, Subbotin, Brown and Moens2001, and L. tabrizicus. The differences between the new species with the aforementioned species follow:
Compared with L. artemisiae, the new species has the shorter body (4.8–5.6 vs. 5.1–6.5 mm), narrower lip region (12–14 vs. 13.8–16.8 μm), shorter odontostyle (67–75 vs. 84–98 μm) and anteriorly located guiding ring (21–24 vs. 27–34 μm from anterior end).
Compared with L. globulicauda, the new species has a lip region separated from the rest body by depression (vs. offset, after the original drawings) and tail in ventral side less concave and almost straight (vs. concave, ventrally slightly bent, after the original drawings). Furthermore, the new species has greater a (124.5–151.5 vs. 80–117), greater b (15.5–17 vs. 12.7–14.9), greater c (129.7–146.7 vs. 78–92), shorter odontophore (43–50 vs. 54–60 μm), anteriorly located guiding ring (21–24 vs. 29–34 μm from anterior end), and males in population vs. not.
Compared with L. patuxentensis, it has pocket-shaped amphidial fovea simple at base (vs. bilobed), narrower vs. wider tail tip, greater a (124.5–151.5 vs. 82.3–119.3), greater c´ (1.3–1.6 vs. 0.9–1.4), narrower lip region (12–14 vs. 15–17 μm wide), shorter odontostyle (67–75 vs. 77–92 μm), and males in population vs. not.
Compared with L. sturhani, the new species has pocket-shaped amphidial fovea simple at the base (vs. bilobed), ventrally almost flat tail with narrower rounded end (vs. concave in ventral side with widely rounded tip), and shorter odontostyle (67–75 vs. 77–96 μm).
Compared with L. tabrizicus, the tentative cryptic form of the new species, it has a narrower rounded end of tail vs. wider and greater a ratio of males (142.6–167.7 vs. 81.5–134) (see below for molecular differences of both species).
Iranian populations of Longidorus mirus Khan, Chawla & Seshadri, Reference Khan, Chawla and Seshadri1971
Figures 4–6 show line drawings and light microphotographs of L. mirus from Bushehr and Southern Khorasan provinces.
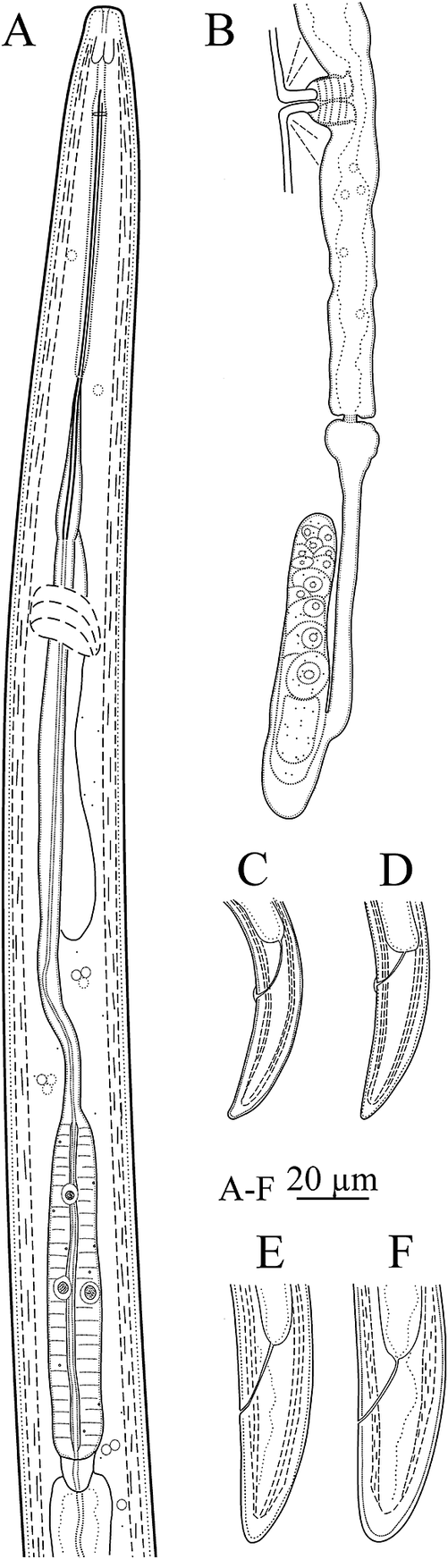
Figure 4. Line drawings of Bushehr population of Longidorus mirus Khan, Chawla & Seshadri, Reference Khan, Chawla and Seshadri1971. (A) pharynx; (B) posterior genital tract; (C–E) tail of first, second, and third juvenile developmental stages, respectively; (F) tail of female.
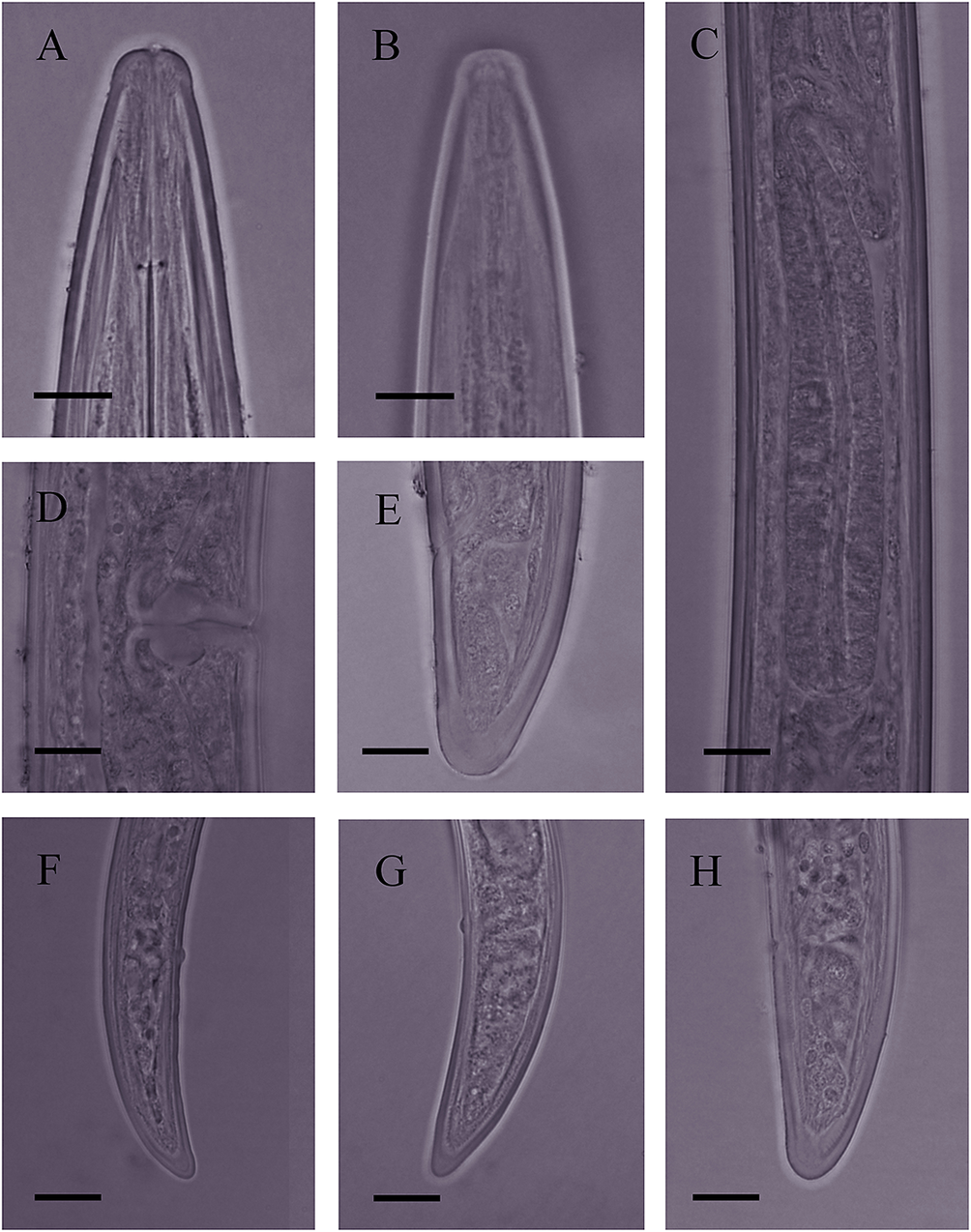
Figure 5. Light microphotographs of Bushehr population of Longidorus mirus Khan, Chawla & Seshadri, Reference Khan, Chawla and Seshadri1971. (A) anterior body region; (B) amphidial fovea; (C) pharyngeal bulb; (D) vulval region; (E) female tail; (F–H) tail of first, second, and third juvenile developmental stages, respectively (scale bar = 10 μm).
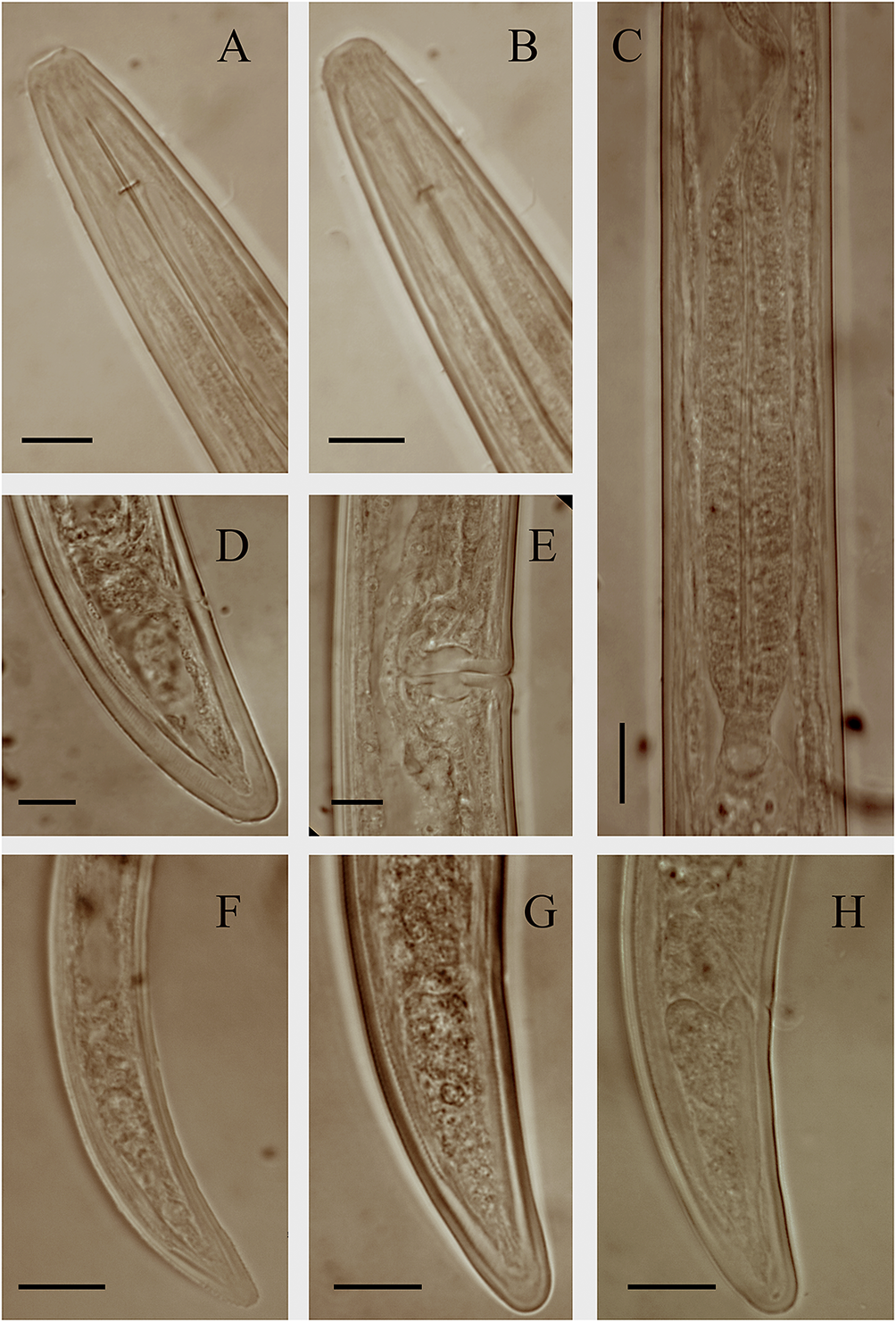
Figure 6. Light microphotographs of Southern Khorasan (Khusf) population of Longidorus mirus Khan, Chawla & Seshadri, Reference Khan, Chawla and Seshadri1971. (A) anterior body region; (B) amphidial fovea; (C) pharyngeal bulb; (D) female tail; (E) vagina; (F–H) tail of first, second, and third juvenile developmental stages, respectively (scale bar = 10 μm).
The morphometrics of L. mirus are presented in Tables 3 and 4.
Descriptions
Female (based on Southern Khorasan and Bushehr populations)
Slender nematodes of medium size, ventrally curved after killing, assuming an open C shape, very gradually narrowing toward both extremities. Some specimens more ventrally bent at the posterior body region. Cuticle two-layered, 2.0–2.8 μm thick at postlabial region, 3.2–4.5 μm thick at midbody, 3.0–4.5 μm thick at the anus, and 5–7 μm thick at tail tip. Lateral chords 25–34.6% of corresponding body width. Lip region almost rounded, separated from the rest of body by a depression. Amphidial fovea pouch-like and symmetrically bilobed, opening not visible. Guiding ring at 2.4–3.0 times lip region width posterior to anterior end. Odontostyle 1.4–1.7 times longer than odontophore, the latter simple. Pharynx dorylaimoid, the pharyngeal bulb occupying 14–33% of the pharynx. The pharyngeal glands nuclei three, their placement as follows: the dorsal gland nucleus (DN) at 26.8–33.7 % and two ventrosublateral nuclei (S1N) at 50–53%. Cardia conoid, 7.0–9.5 μm high and 8.8–10.5 μm wide. Reproductive system didelphic-amphidelphic, both branches almost equally developed, generally small, each composed of a reflexed ovary 82–131 μm long, oviduct 110–177 μm long, sphincter between the oviduct and uterus, the latter tubular 52–57 μm long. Vagina 49.53–63.12% of corresponding body width long. Vulva a transverse slit in ventral view. Prerectum 3.2–4.6, and rectum 0.7–1.0 times anal body width long. Tail conical, dorsally convex, ventrally flat with widely rounded tip.
Male
Not Found.
Juveniles (based on Southern Khorasan and Bushehr populations)
Three juvenile developmental stages (J1–J3) were identified and separated according to Robbins et al. (Reference Robbins, Brown, Halbrendt and Vrain1995). Their scatter diagram based on the relationships between body length and the functional and replacement odontostyle of juveniles and females of populations from Bushehr province is given in Figure 7. Juveniles similar to females in appearance, except for their smaller body, replacement odontostyle preserved in the anterior narrow region of pharynx, and the reproductive system not developed. The first juvenile J1 with a replacement odontostyle lying on the odontophore, its tip close to the base of the functional odontostyle. Its tail conical, dorsally convex, ventrally concave with rounded tip. In the second and third juvenile developmental stages (J2 and J3), the replacement odontostyle far from the functional odontostyle. J2 with a conical tail similar to that in J1, but thicker, and J3 with a dorsally convex and ventrally almost flat tail with a widely rounded tip.

Figure 7. Scatter plot of the functional and replacement odontostyle length in relation to the body length of the juvenile stages and females of Longidorus mirus Khan, Chawla & Seshadri, Reference Khan, Chawla and Seshadri1971.
Remarks
Longidorus mirus was originally described from India in association with maize and is currently only known for its type locality (Khan et al. Reference Khan, Chawla and Seshadri1971). The morphological features and the morphometrics of the Iranian populations of L. mirus are in accordance with the type population of the species. The matrix code of the Iranian populations according to Chen et al. (Reference Chen, Hooper, Loof and Xu1997) and Peneva et al. (Reference Peneva, Lazarova, De Luca and Brown2013) are: A23-B12-C23-D2-E2-F2-G12-H23-I1-J2-K6, similar to the codes provided for the species by Chen et al. (Reference Chen, Hooper, Loof and Xu1997). The GPS information of the Bushehr and Southern Khorasan populations are as follows: N29°.27.567′, E51°17.006′ for Bushehr population, and N32°46.2373′, E58°52.2689′ and N34°04.4893′, E057°19.5398′ for two Southern Khorasan populations. The Bushehr province population was recovered from the rhizosphere of date palm in the Vahdatiyeh region, and the Southern Khorasan populations were recovered from the rhizosphere of narcissus in Khusf region and pistachio in the city of Boshruyeh.
Longidorus mirus has close morphology and morphometrics with L. auratus Jacobs & Heyns, Reference Jacobs and Heyns1987 and L. africanus Merny, Reference Merny1966. Compared to L. auratus (for original morphometrics of L. auratus, see Table 4), it has different a value (70–90 in the type population of L. mirus vs. 94–141 in the type population of L. auratus).
Table 4. Morphometrics of Longidorus mirus Khan, Chawla & Seshadri, Reference Khan, Chawla and Seshadri1971 from Southern Khorasan province and type population of L. auratus Jacobs & Heyns, Reference Jacobs and Heyns1987. All measurements are in μm and in the form: mean ± S.D. (range)

The juvenile developmental stages of L. auratus are, however, not known, and molecular data for type or topotype populations of both species are lacking. Females of Iranian populations of L. mirus closely resemble females of L. africanus, and they form cryptic forms of each other. In the absence of all juvenile developmental stages and molecular data, both species are indistinguishable (see below).
Bushehr province population of Longidorus persicus Esmaeili, Heydari, Archidona-Yuste, Castillo & Palomares-Rius, Reference Esmaeili, Heydari, Archidona-Yuste, Castillo and Palomares-Rius2017
Figure 8 shows light microphotographs of L. persicus

Figure 8. Light microphotographs of Bushehr population of Longidorus persicus Esmaeili, Heydari, Archidona-Yuste, Castillo & Palomares-Rius, Reference Esmaeili, Heydari, Archidona-Yuste, Castillo and Palomares-Rius2017. (A) anterior body region; (B) vulval region; (C and D) female tail (scale bar = 10 μm).
The morphometrics of L. persicus are presented in Table 5.
Table 5. Morphometrics of Bushehr province populations of Longidorus persicus Esmaeili, Heydari, Archidona-Yuste, Castillo & Palomares-Rius, Reference Esmaeili, Heydari, Archidona-Yuste, Castillo and Palomares-Rius2017 and L. orientalis Loof, Reference Loof1982. All measurements are in μm and in the form: mean ± S.D. (range)
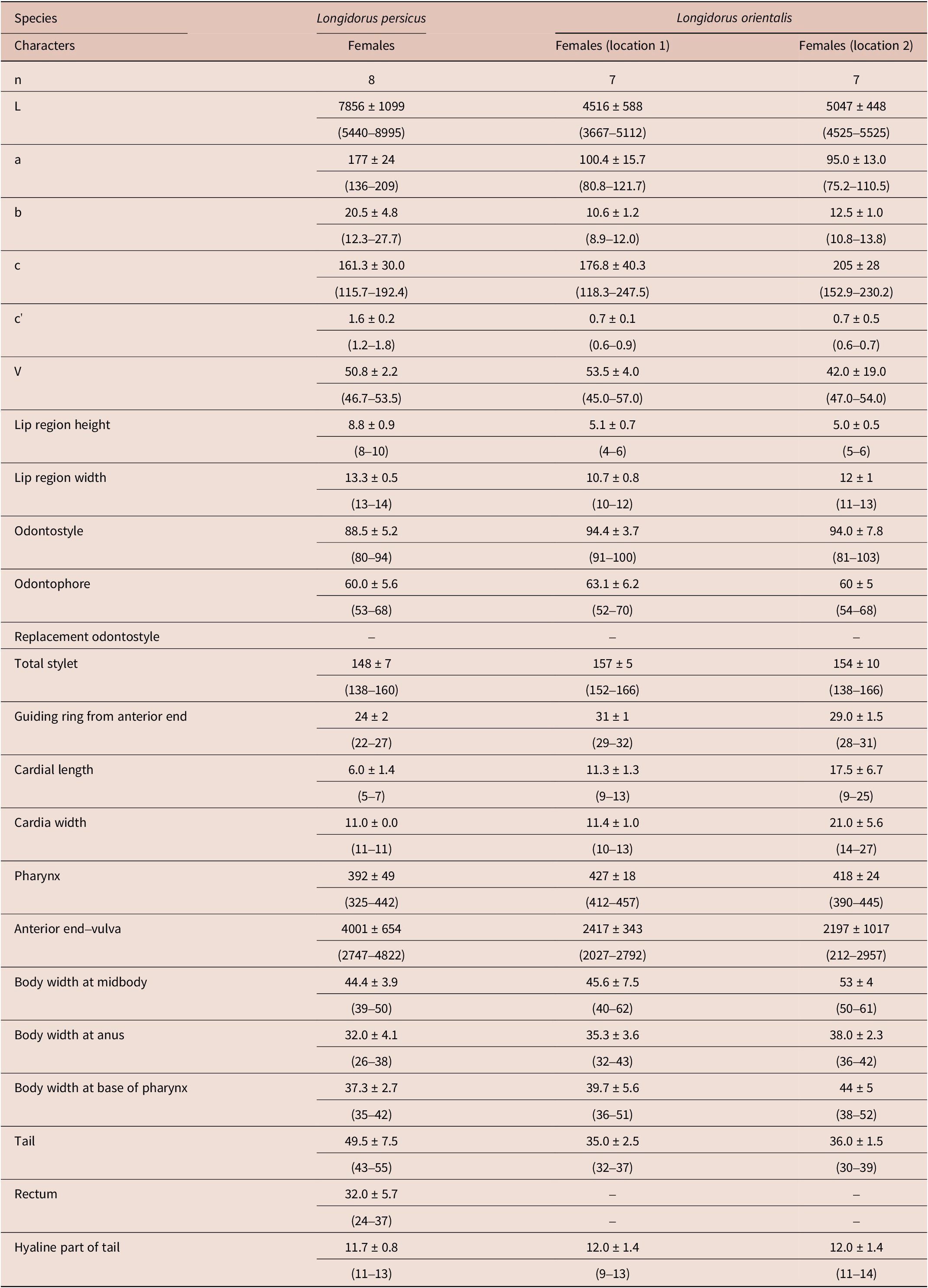
During the present study, Longidorus persicus was recovered from the rhizosphere of date palm in the Dashtestan region, Bushehr province. This is the second report of the species after its description from western Iran (Esmaeili et al. Reference Esmaeili, Heydari, Archidona-Yuste, Castillo and Palomares-Rius2017), extending its distribution area to southern Iran. The Bushehr population has no remarkable morphological or morphometric differences with the type population. The GPS information of the occurring location of the species is as follows: N29°2.625′, E50°52.324′.
Bushehr province population of Longidorus orientalis Loof, Reference Loof1982
Figure 9 shows light microphotographs of L. orientalis.
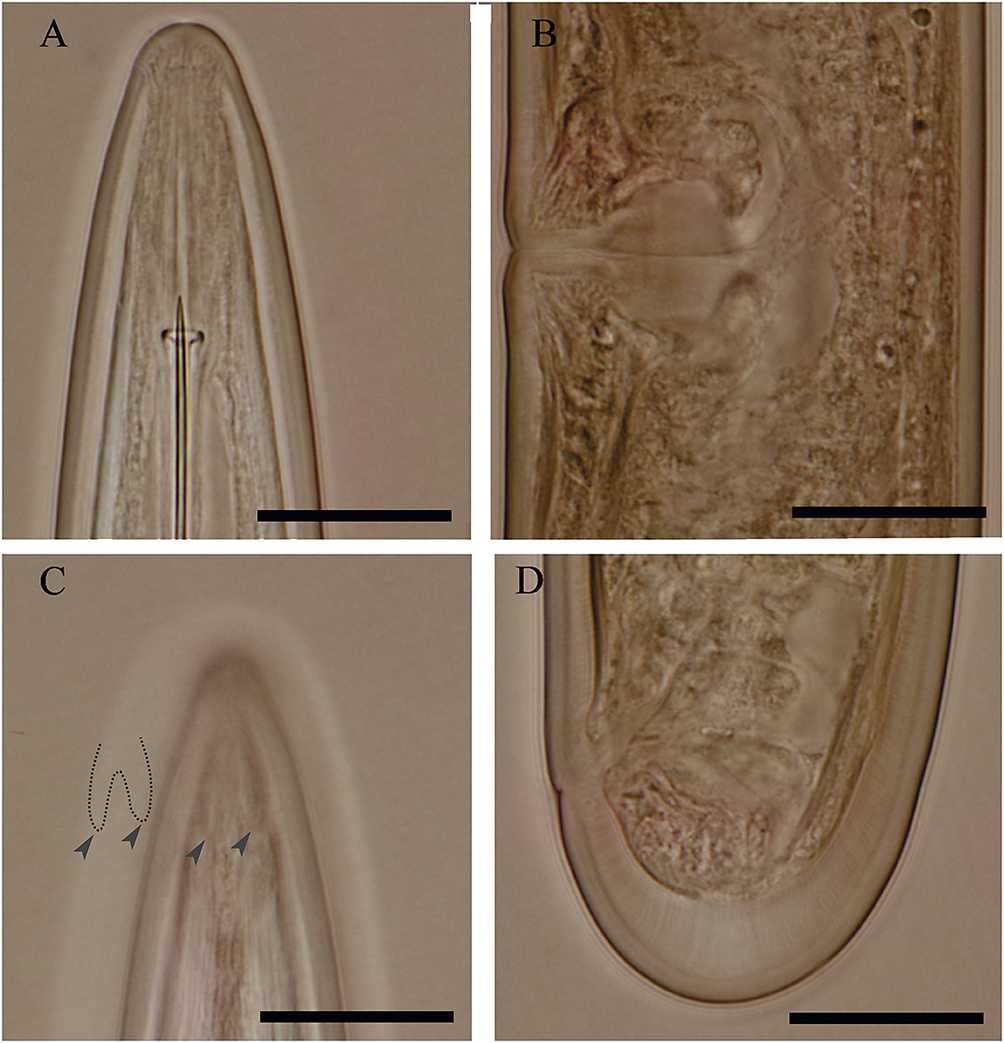
Figure 9. Light microphotographs of Bushehr population of Longidorus orientalis Loof, Reference Loof1982. (A) anterior body region; (B) vulval region; (C) amphidial fovea; (D) tail (scale bar = 20 μm).
The morphometrics of two populations of L. orientalis (out of several recovered populations) are presented in Table 5.
This species is common in southern Iran (Pedram Reference Pedram, Ghaderi, Kashi and Karegar2018). During the present study, it was observed in almost all soil samples collected from the Dashtestan region in remarkably high population density. The recovered populations had no morphological and morphometric differences with the type population or other populations reported from the country. The GPS information of two locations having remarkably high population density is as follows: location 1, N29°.26.110′, E51°10.493′; location 2, N29°.26.294′, E51°11.545′.
Molecular characterisations and phylogenetic analyses
Information on the newly generated sequences during the present study, including the accession numbers and the sequenced loci as well as their length (bp), are given in Table 1.
The BLAST search using the newly generated SSU sequence for Longidorus paratabrizicus n. sp. revealed it has 98.09–98.79% identity with several SSU sequences of Longidorus spp. already deposited into the database. In SSU phylogeny (Figure 10), the newly generated sequence of L. paratabrizicus n. sp. occupied a distant placement compared with the original SSU sequences of its cryptic form, L. tabrizicus. The BLAST search using the newly generated LSU sequences of the new species revealed their identity with currently available LSU sequences is less than 93%. Both newly generated sequences of L. tabrizicus and L. paratabrizicus n. sp. occupied distant placements in the LSU tree (Figure 11). By less coverage value of the ITS sequence of the new species while BLAST search with other sequences of the genus already deposited into the database, no further phylogenetic analysis was performed using ITS sequences (the maximum coverage, 64%, belonged to AJ549988, assigned to L. profundorum Hooper, Reference Hooper1996, and its coverage value with other sequences was less than 40%).

Figure 10. Bayesian 50% majority rule consensus tree inferred from the SSU rDNA of Longidorus paratabrizicus n. sp. and Bushehr population of L. mirus Khan, Chawla & Seshadri, Reference Khan, Chawla and Seshadri1971 under GTR + G + I model. Bayesian posterior probability values >0.50 are given for corresponding clades. The newly generated sequences are in bold font. GTR, general time-reversible; G, gamma; I, invariant; rDNA, ribosomal DNA; SSU, small-subunit.
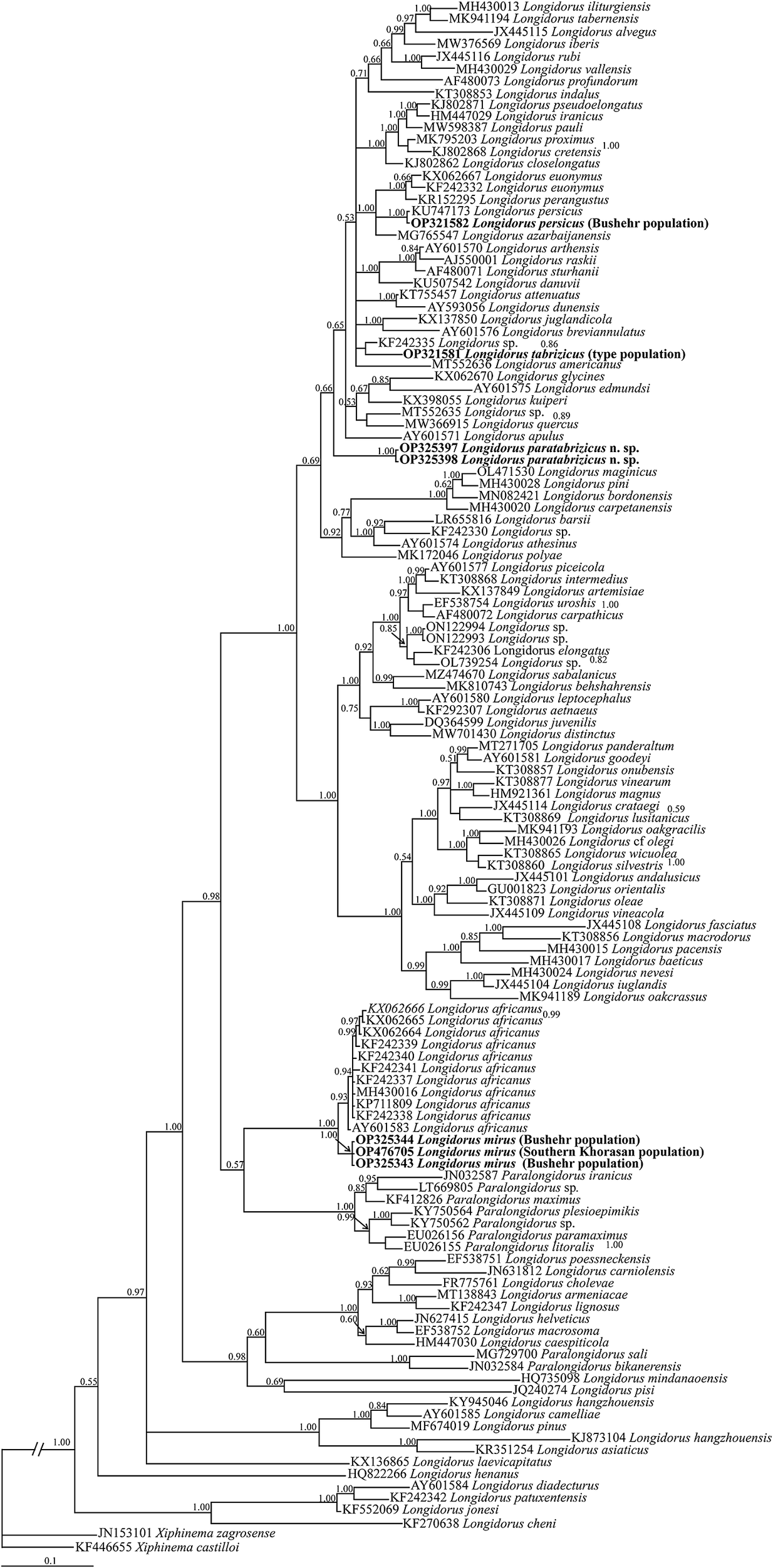
Figure 11. Bayesian 50% majority rule consensus tree inferred from the LSU rDNA D2-D3 sequences of Longidorus paratabrizicus n. sp., Iranian populations of L. mirus Khan, Chawla & Seshadri, Reference Khan, Chawla and Seshadri1971, Bushehr population of L. persicus Esmaeili, Heydari, Archidona-Yuste, Castillo & Palomares-Rius, Reference Esmaeili, Heydari, Archidona-Yuste, Castillo and Palomares-Rius2017, and type population sequence of L. tabrizicus Niknam, Pedram, Ghahremani Nejad, Ye, Robbins & Tanha Maafi, 2010 under the GTR + G + I model. Bayesian posterior probability values >0.50 are given for the corresponding clades. The newly generated sequences are in bold font. GTR, general time-reversible; G, gamma; I, invariant; LSU, large-subunit; rDNA, ribosomal DNA.
The BLAST search using the SSU sequence of the Bushehr population of L. mirus revealed it has a 98.14–99.94% identity with several SSU sequences of Longidorus spp. available in GenBank. In the SSU tree (Figure 10), the SSU sequence of Bushehr province population of L. mirus formed a low-supported clade with another clade including sequences assigned to L. africanus (AY283164 and KF242279) and three other sequences of Longidorus sp., L. ferrisi Robbins, Ye & Pedram, Reference Robbins, Ye and Pedram2009, and Paralongidorus sali Siddiqi, Hooper & Khan, Reference Siddiqi, Hooper and Khan1963.
The three newly generated LSU sequences for Iranian populations of L. mirus were almost identical, and only two degenerate codes were observed. The BLAST search of the longest sequence (OP325343) of L. mirus revealed its highest identity values (97.93–98.35% identity, 12–16 mismatches) belong to sequences KX062666, KX062665, KX062664, KF242339, KF242340, KF242341, KF242337, MH430016, KP711809, KF242338, and AY601583 assigned to several isolates of L. africanus. The identity values with other sequences assigned to other species were less than these values. In the LSU tree (Figure 11), sequences of Iranian populations of L. mirus have formed a clade with several isolates of L. africanus from all over the world.
The two ITS sequences (OQ476241, OQ476242) generated for Bushehr and Southern Khorasan populations of L. mirus were different in length but almost identical in their overlapping region while aligning and had only one gap (real difference) and two degenerate differences while aligning. The BLAST search of OQ476241 revealed it has 99.61–99.74% identity (two to three mismatches, one gap) with several sequences assigned to Longidorus africanus from Iran under the accession numbers AB537952, AB537953, AB537955, AB537956, AB537958, AB537959, and AB537960. Its identity with another Iranian sequence AB537957 assigned to L. africanus was 97.69% (13 indels and nine gaps). Its identity with two other currently available ITS sequences assigned to L. africanus from Europe (KX062689 and KX062690) was 98.15 and 97.07% (24 indels and seven gaps; and 15 indels and four gaps). The BLAST search of OQ476242 revealed it has 99.61–99.62% identity (two to three indels, one gap) with seven aforementioned sequences assigned to Longidorus africanus from Iran. Its identity with two European sequences (KX062689 and KX062690) was 95.62 and 95.45% (20 indels and 18 gaps; 47 indels and 27 gaps, respectively). The BLAST search using two newly generated ITS sequences of L. mirus further showed their coverage with sequences other than aforementioned accession numbers is 49%, belonging to OP391507, assigned to Longiorus sp., and less than 20% compared with all other sequences; as the result, no further phylogenetic analysis was performed using ITS sequences.
The newly generated LSU sequence for the Bushehr population of L. persicus was identical to that of the type population, and the phylogenetic relationships of the clade of the two sequences of L. persicus with some other sequences were not resolved in Figure 11 LSU tree, due to polytomy.
Discussion
The herein-described new species, Longidorus paratabrizicus n. sp., represents another example of cryptic speciation of soil-inhabiting nematodes. It is morphologically close to L. tabrizicus. The difference in female tail shape, compared with the common form in L. tabrizicus and male index a, could be interpreted as intraspecies variations in lacking molecular data. Furthermore, L. tabrizicus has three juvenile developmental stages, but the juvenile developmental stages of the new species were not determined as only two pre-last and last juvenile stages were recovered. Formerly, the characters of juveniles have been used to separate cryptic forms of Longidorus (Cai et al. Reference Cai, Archidona‐Yuste, Cantalapiedra‐Navarrete, Palomares‐Rius and Castillo2020b). Both species, L. tabrizicus and L. paratabrizicus n. sp., occupied distant placements in SSU and LSU trees, as already presented, corroborating that they are separate species, cryptic forms of each other. As already emphasised, molecular data should be exploited to separate cryptic forms of Longidorus (Cai et al. Reference Cai, Archidona‐Yuste, Cantalapiedra‐Navarrete, Palomares‐Rius and Castillo2020b). Formerly, several other examples of cryptic speciation have been documented for longidorids (e.g. Pedram et al. Reference Pedram, Pourjam, Namjou, Atighi, Cantalapiedra-Navarrete, Liébanas, Palomares-Rius and Castillo2012; Archidona-Yuste et al. Reference Archidona-Yuste, Navas-Cortés, Cantalapiedra-Navarrete, Palomares-Rius and Castillo2016) or other groups (e.g. Cantalapiedra-Navarrete et al. Reference Cantalapiedra-Navarrete, Navas-Cortés, Liébanas, Vovlas, Subbotin, Palomares-Rius and Castillo2013; Clavero-Camacho et al. Reference Clavero-Camacho, Palomares-Rius, Cantalapiedra-Navarrete, León-Ropero, Martín-Barbarroja, Archidona-Yuste and Castillo2021). As noted by Jahanshahi Afshar (Reference Jahanshahi Afshar2019), sequences from type or topotype specimens are useful for reliable identifications using molecular data, and in the case of the presently described new species, the analyses using the newly generated LSU sequence of the type specimens of L. tabrizicus, in addition to the results obtained from analysis of its original SSU data, further confirmed the two species are separate.
Longidorus mirus looks similar to L. auratus, but there are differences in some morphometric indices like index a. They represent tentative cryptic forms and in lacking juveniles characters and molecular data, their separation could be problematic. As a result, a new synonymy was however not proposed in this relation, and future data may better clarify their taxonomic status. It has close morphology with L. africanus, too. However, the number of juvenile developmental stages are useful to separate both species. L. mirus has three juvenile developmental stages (see above), whereas L. africanus has four juvenile developmental stages (Bravo and Roca Reference Bravo and Roca1995). The number of juvenile developmental stages has previously been used to separate two closely resembling longidorid species (Jahanshahi Afshar et al. Reference Jahanshahi Afshar, Pedram and Mobasseri2021). Although sequences from the type specimens of L. africanus are currently not available, the presently generated LSU sequences of L. mirus form a separate sister clade to several sequences assigned to L. africanus from other localities in the LSU tree. There are only two SSU sequences (KF242279, AY283164) assigned to L. africanus currently deposited into the Genbank database. However, their identity needs further validation, and again, similar to previous cases (Jahsnahshai Afshar Reference Jahanshahi Afshar2019; Lazarova et al. Reference Lazarova, Oliveira, Prior, Peneva and Kumari2019), sequences from type or topotype populations could clarify their status. Finally, based upon high identity value of ITS sequences of L. mirus with accession numbers AB537952, AB537953, AB537955, AB537956, AB537958, AB537959 and AB537960 assigned to “L. africanus” from Iran, it may be concluded that these seven sequences belong to L. mirus. The occurrence and high density of L. persicus and L. orientalis in date palm gardens of the Bushehr province, in addition to two former species prevalent in the region, could be a warning of their potential threat to this commercial crop, an open field for future study.
Acknowledgements
The kind help of Eng. Fatemeh Bazeghi with critically reading the MS is appreciated.
Financial support
We appreciate the financial support of Ferdowsi University of Mashhad and Tarbiat Modares University and the scientific support of Southern Khorasan Agriculture and Natural Resources Research and Education Centre.
Competing interest
There are no competing interests.
Ethical standards
The authors assert that all procedures contributing to this work comply with the ethical standards of the relevant national and institutional guides on the care and use of laboratory animals.
















