



Non-technical Summary
The Late Cretaceous gastropods from Turkey have been insufficiently studied. We herein present a taxonomic study of the subgenus Trochactaeon (Trochactaeon) from the upper part of the Karababa Formation (Karababa-C member) at the Saytepe section in southeastern Turkey. The Upper Cretaceous fossil-bearing Karababa Formation, cropping out in southeastern Turkey, yields a remarkable assemblage of acteonellid gastropods and rudist bivalves. The subgenus Trochactaeon, a very successful and widespread taxon within the heterobranch gastropod family Acteonellidae, dominated acteonellid assemblages throughout the Late Cretaceous. This is the first record of Trochactaeon (Trochactaeon) giganteus subglobosus from Turkey. It is from a single lower Campanian bed in the upper part of the Karababa Formation of the Gölbaşı region (south of Adıyaman), corresponding to the northwestern part of the Arabian Platform. This record complements information on the temporal and spatial distribution of Trochactaeon at the southern margin of the Tethyan Ocean during the last part of the Cretaceous Period. This discovery increases the documented diversity of the paleofauna from the Upper Cretaceous succession in southeastern Turkey and provides new insights into the paleoenvironment of the carbonate ramp of the northern Arabian plate, and the paleobiogeography of Campanian gastropods in general.
Introduction
The Cenomanian–Maastrichtian time span is marked in the Tethyan realm by paleogeographic and paleoenvironmental changes that had a profound effect on the gastropod biodiversity (Sohl, Reference Sohl1987). Members of Acteonellidae Gill, Reference Gill1871, which are firmly nested within the Heterobranchia, represent some of the most important components of marine communities during the Cretaceous (Kollmann, Reference Kollmann2014). From the gastropod point of view, and more particularly from that of the acteonellids, the Late Cretaceous is marked by great diversification of representatives of the family Acteonellidae (Sohl and Kollmann, Reference Sohl and Kollmann1985; Kollmann, Reference Kollmann2014). The currently known European record of acteonellid gastropods is relatively abundant and diverse, especially in the Upper Cretaceous strata of the Eastern Alps (Kollmann, Reference Kollmann1967). However, records of Cretaceous gastropods from Turkey are very rare and they are usually limited to a few specimens. Cretaceous strata of the Adıyaman area of southeastern Turkey comprise a relatively thick succession that is superbly exposed and relatively fossiliferous (Fig.1.1). These strata form an important sedimentary archive for understanding of the paleogeography of the northern Arabian Platform, which formed part of an extensive carbonate platform on the southern margin of the Tethys Sea (Yılmaz et al., Reference Yılmaz, Cook, Hoşgör, Wagreich, Rebman and Murray2018). Thus, any new addition to the faunas of poorly known regions, such as northern Arabia, provides new and interesting material for paleobioegeographical analyses.
Until we began our exploration and study on Cretaceous gastropods of Turkey, only a few nominate gastropod taxa had been known. For many years, a Late Cretaceous gastropod assemblage found in northwestern Anatolia and described by Stchepinsky (Reference Stchepinsky1942) was the only assemblage known from the entire area. Cretaceous rocks are widespread in southeastern Turkey, however only echinoderm remains, belemnitellids, venerid bivalves, and rudist faunas have been described (Özer, Reference Özer1992; Farinacci and Manni, Reference Farinacci and Manni2003; Hoşgör and Kostak, Reference Hoşgör and Kostak2012; Mülayim et al., Reference Mülayim, Yılmaz and Ferré2018; Hoşgör and Yılmaz, Reference Hoşgör and Yılmaz2019).
During the Late Cretaceous, the Gaziantep, Adıyaman, and Mardin-Derik region was located at the southern margin of the Tethys Sea, prior to its closure (Hoşgör and Yılmaz, Reference Hoşgör and Yılmaz2019). Shallow marine carbonate successions deposited in the area record several successive transitions from outer shelf to inner shelf and platform conditions (Yılmaz et al., Reference Yılmaz, Cook, Hoşgör, Wagreich, Rebman and Murray2018). The Upper Cretaceous Karababa Formation forms the upper part of the Mardin Group, a thick succession of mostly carbonate Cretaceous rocks known from outcrops and the subsurface in southeastern Turkey (Çelikdemir et al., Reference Çelikdemir, Dülger, Görür, Wagner, Uygur and Spencer1991). Here, we provide the first record of a remarkable mollusk assemblage from an isolated outcrop of bioclastic limestones of the Karababa Formation (Fig. 1.2).

Figure 1. (1) Geological map of southeast Turkey (Hoşgör and Yılmaz, Reference Hoşgör and Yılmaz2019). (2) Regional geological map of Akpınar Village near the Gölbaşı area showing the location of the studied outcrops and the cross section.
Knowledge of Cretaceous gastropods from the northern Arabian Platform has been restricted to general descriptions of faunal contents of Cretaceous strata presented by Conrad (Reference Conrad and Lynch1852), Blankenhorn (Reference Blanckenhorn1890, Reference Blanckenhorn1927), Böhm (Reference Böhm1900), and Delpey (Reference Delpey1940). Trochactaeon is an extinct genus of acteonellid gastropods that had significant occurrences in the circum-equatorial Tethyan belt (Albian–Maastrichtian). Distribution and diversity patterns of acteonellids in the Cretaceous have been well studied (e.g., Kollmann, Reference Kollmann1967, Reference Kollmann2014; Sohl and Kollmann, Reference Sohl and Kollmann1985; Kowalke and Bandel, Reference Kowalke and Bandel1996). The Late Cretaceous paleogeography of the European, North African, and southwest Asian branches of the Tethyan Realm also allowed a wide geographical dispersal of the Acteonellidae, especially from the time of sea-level rise in the Cenomanian and onwards. The discovery presented here provides additional information and extends the known geographical distribution of Trochactaeon beyond Europe, indicating that it had a larger paleogeographical distribution than was previously thought. Previously, the ‘Trochactaeon’ assemblage was also widely recorded in the southern ranges of the Greater Caucasus and Europe through field-work studies mainly concerning biostratigraphic analysis of depositional architecture for the Upper Cretaceous successions (Pčelintsev, Reference Pčelintsev1953; Pokorný, Reference Pokorný1959; Kollmann, Reference Kollmann1967; Czabalay, Reference Czabalay1973; Akopjan, Reference Akopjan1976). This paper deals with the taxonomic description, stratigraphic and geographic ranges, and the paleoenvironmental significance of Trochactaeon (Trochactaeon) giganteus subglobosus (Münster, Reference Münster and Goldfuss1844) in the Adıyaman area (SE Turkey).
Geological setting
The Karababa Formation is exposed in extensive outcrops in the Antakya, Adıyaman, Diyarbakır, and Şırnak regions in southeastern Turkey, where it forms the upper part of the Mardin Group carbonates, which are part of the Arabian platform succession (Fig. 1.1). Numerous studies have been carried out on the Karababa Formation, including microfacies and sequence stratigraphy, geochemistry, and reservoir characterization (Çelikdemir et al., Reference Çelikdemir, Dülger, Görür, Wagner, Uygur and Spencer1991; Cater and Gillcrist, Reference Cater and Gillcrist1994; Yılmaz et al., Reference Yılmaz, Cook, Hoşgör, Wagreich, Rebman and Murray2018). However, there is little information about macrofossil assemblages of this formation in SE Turkey. The age of the Karababa Formation has been reported in the literature as late Coniacian to early Campanian, based on biostratigraphic studies of benthic and planktonic foraminifera (Çoruh et al., Reference Çoruh, Yakar and Ediger1997; Mülayim et al., Reference Mülayim, Mancini, Çemen and Yılmaz2016; Yılmaz et al., Reference Yılmaz, Cook, Hoşgör, Wagreich, Rebman and Murray2018).
The Karababa Formation is subdivided into three informal members designated by the letters A, B, and C from base to top. The Karababa-A member is a dark brownish gray, very fine-grained limestone. It contains abundant organic matter and pelagic foraminifera. The conformably overlying Karababa-B member, which was deposited in a shallower-water environment than the Karababa-A member, is composed of calcareous packstones/wackestones that are rich in pelagic foraminifera. The overlying Karababa-C member is a bioclastic limestone that is partly dolomitized (Yılmaz et al., Reference Yılmaz, Cook, Hoşgör, Wagreich, Rebman and Murray2018). The depositional succession of these members within a shallow marine sequence has been described from several exploration wells (Mülayim et al., Reference Mülayim, Mancini, Çemen and Yılmaz2016). Few outcrop studies have been published on the Karababa Formation, and these consider only the biostratigraphic aspects of a limited number of sections (Mülayim et al., Reference Mülayim, Mancini, Çemen and Yılmaz2016; Yılmaz et al., Reference Yılmaz, Cook, Hoşgör, Wagreich, Rebman and Murray2018). The Karababa Formation is unconformably (or conformably in some localities) underlain by the Derdere Formation (Cenomanian–Turonian) and unconformably overlain by the Karaboğaz and Sayındere formations (Campanian) (Fig. 1.2).
Saytepe section
The Saytepe section was measured in the Karababa Formation (C-member) (Fig. 2). The base of the section consists of 15-m-thick, medium-bedded limestones, in which macro- and microfossils are absent. These limestones pass upward into rudist-bearing limestones interbedded with algal limestones (~40 m thick). Three rudist levels can be distinguished, from bottom to top (Fig. 2.2). The first (lower) rudist level (40–50 cm thick, yellowish gray) consists of rare radiolitid valves and valve fragments. Some left valve transverse sections show a single row of radial canals like those seen in Joufia serbica (Milovanović, Reference Milovanović1956), which recently was described by Özer et al. (Reference Özer, Skelton, Tarlao and Tunis2021), and radiolitid right valves (?Sauvagesia sp.) (Fig. 2). The second rudist level (60–70 cm thick, dark gray) is composed mainly of hippuritid right valves, usually exposed as transverse and slightly oblique sections scattered in a wackestone–packstone texture showing diagenetic effects (dolomitization, compaction, and micritization). Very sparse radiolitids are also present. Vaccinites loftusi (Woodward, Reference Woodward1855) is abundantly represented, but some specimens resemble Pironaea sp. (Munujos et al., Reference Munujos, Pons and Vicens2016). The third level (50–60 cm thick, gray) is directly above the previous rudist level and consists of radiolitids exposed in transversal, oblique, and radial right-valve sections and abundant fragments. Biradiolites bulgaricus Pamouktchiev, Reference Pamouktchiev1967, Biradiolites sp., Bournonia cf. B. dinarica Slišković, Reference Slišković1983, Durania sp., and Radiolites sp. can be identified. This level is composed of packstones–wackestones that include radiolitid fragments, some of which are very thick. The right valve transverse sections of radiolitids represent isolated scattered individuals in life position, but abundant disoriented and reworked radiolitid fragments also are present. The distribution of these rudist species in the Mediterranean area (Steuber, Reference Steuber2002) suggests a Campanian age for the rudist fauna.

Figure 2. (1) Field views of lithofacies in the Saytepe section, acteonellid assemblage slabs, and photomicrographs of the packstone–grainstone microfacies; scale bars = 100 μm. (2, 3) First rudist level showing the left valve transverse sections of Joufia serbica (white arrows), indeterminable radiolitid right valve transverse and oblique sections (black arrows), and radiolitid right valve fragments (dashed black arrows). Second rudist level showing the right valve transverse sections of Vaccinates loftusi. Note diagenetic effects. Third rudist level consists mainly of radiolitid transverse, oblique, and radial right valve sections. Scales are hammer and 10-mm scale bars.
The uppermost part of the 15-m-thick sequence is characterized by the occurrence of acteonellid assemblages (Fig. 2.1, 2.2). The upper part of the Karababa Formation comprises mainly monospecific acteonellids, Trochactaeon (Trochactaeon) giganteus subglobosus (Münster, Reference Münster and Goldfuss1844) at the Saytepe locality. At this site, the thickness of the shell beds was measured, and the preserved parts of the shells were internal molds. The available top bedding surface of the acteonellid shell beds at this site was recorded in a packstone–grainstone bed (Fig. 2.1).
Material and methods
Preserved specimens of Trochactaeon described herein were collected from above the rudist-bearing limestone assigned to the Karababa Formation (C-member) in the Gölbaşı valley near Besni (37.75472°N, 37.7513938°E; topographical map sheet 1:25,000; M39-c) in the Adıyaman region. The studied stratigraphic section was measured on a bed-by-bed scale, for a total thickness of 70 m (Fig. 2). Stratigraphic and microfacies analyses were performed at the Sedimentology Laboratory of the Department of Geological Engineering, Middle East Technical University (METU), Ankara, Turkey. Microfacies determinations were carried out by a point-counting/visual estimation method on thin sections with a James Swift point-counting apparatus and an Olympus CX31 polarizing microscope, based on the principles of microfacies analysis of Flügel (Reference Flügel2010).
The Trochactaeon bed yielded enough isolated and complete specimens to illustrate the diversity of their internal morphology. The descriptions and measurements in the systematic paleontology section follow Kollmann (Reference Kollmann1967) and Sohl and Kollmann (Reference Sohl and Kollmann1985). The classification used in this work follows Bouchet et al. (Reference Bouchet, Rocroi, Hausdorf, Kaim, Kano, Nützel, Parkhaev, Schrödl and Strong2017). The synonymies contain only the most recent references, as well as those of importance to the discussion. All linear measurements are given in millimeters.
The rudists are embedded in limestone, therefore they were identified in the field and from field photographs (Fig. 2). They have undergone some diagenetic alteration (e.g., dolomitization, recrystallization, dissolution, and compaction).
Repositories and institutional abbreviation
Most of the fossil specimens, as well as any additional material illustrated here, are stored in the Sedimentology Laboratory of the Department of Geological Engineering, Middle East Technical University (METU), Ankara, Turkey. Figured specimens are designated in the descriptions by the prefix METUGM (Middle East Technical University Geology Museum).
Systematic paleontology
Class Gastropoda Cuvier, Reference Cuvier1795
Subclass Heterobranchia Burmeister, Reference Burmeister1837
Superfamily Acteonoidea d'Orbigny, Reference d'Orbigny1843
Family Acteonellidae Gill, Reference Gill1871
Genus Trochactaeon Meek, Reference Meek1863
Subgenus Trochactaeon Meek, Reference Meek1863
Type species
Acteonella renauxiana d'Orbigny, Reference d'Orbigny1842, by original designation; Turonian, France.
Diagnosis
Subgeneric diagnosis rephrased from Kollmann (Reference Kollmann1967) and Sohl and Kollmann (Reference Sohl and Kollmann1985): the greatest shell width is above mid-height, and the spire comprises less than half of the total shell height. The aperture is narrow, parietal lip smooth; columella with three plaits, one as a high palatal ridge.
Trochactaeon (Trochactaeon) giganteus subglobosus (Münster, Reference Münster and Goldfuss1844)
Figures 3, 4.3
- Reference Münster and Goldfuss1844
Tornatella subglobosa Münster, p. 49, pl. 177, fig. 13a, b.
- Reference Pokorný1959
Acteonella (Trochactaeon) gigantea subglobosa Pokorný, p. 959, textfig. 1, pl. 1, fig. 3.
- Reference Kollmann1967
Trochactaeon (Trochactaeon) giganteus subglobosus Kollmann, p. 240, pl. 3. figs. 15, 19, pl. 8. figs. 53–55, pl. 9, figs. 56, 57.
- Reference Czabalay1973
Trochactaeon (Trochactaeon) giganteus subglobosus; Czabalay, p. 293, pl. 5, fig. 1, pl. 6, fig. 2.
- Reference Forner and Calzada1998
Trochactaeon (Trochactaeon) giganteus subglobosus; Forner and Calzada, p. 31, pl. 1, fig. 2.

Figure 3. (1–6) Trochactaeon (Trochactaeon) giganteus subglobosus (Münster, Reference Münster and Goldfuss1844), axial sections, METUGM 2218-01-06, specimens from Saytepe section. All scale bars represent 10 mm.

Figure 4. Morphologic features of Trochactaeon (Trochactaeon) shells in longitudinal cross-section, highlighting the plication (fold) of the palatal (outer lip) and the columellar folds.
Holotype
An internal mold (Nr.751/1), Santonian–lower Campanian (Kollmann, Reference Kollmann1967), Hieflau, Austria (Eastern Alps).
Description
Recrystallized, moderately large shells, approximately spherical shape, low-spired with apical angle >120° (Fig. 4). Aperture proportionally broad, columella thick, columellar lip only slightly expanded and bearing three plaits, low palatal ridge occurs within penultimate whorl, axial whorl section broad and inclined. Outer lip opisthocline at ~30° to shell axis.
Material and measurements
The largest of the measured specimens (measurements in mm) is shown in Figure 3.5 (H: 155, MW: 135) (H: Height, MW: Maximum width, in mm). A small specimen is shown in Figures 3.3 and 4.3 (H: 70.4; MW: 68.5). The other four specimens available for measurement are illustrated in Figures 3.1 and 4.3 (H: 110.8; MW: 96), Figure 3.6 (H: 70; MW: 75), Figure 3.4 (H: 71; MW: 62), and Figure 3.2 (H: 145; MW: 105).
Remarks
The majority of Cretaceous acteonellids from Europe, Caucasas, and Arabia (and in the present study) are preserved as recrystallized shells. The family Acteonellidae was created by Gill (Reference Gill1871) and its distinctive characters were presented and discussed in detail by Pčelintsev (Reference Pčelintsev1953), Kollmann (Reference Kollmann1967), Akopjan (Reference Akopjan1976), and Sohl and Kollmann (Reference Sohl and Kollmann1985). The internal characters are best seen in axial sections cut through the center of the columella and these include the most diagnostic features of acteonellids. The results of numerous twentieth century works saw a progressive accumulation of data on the recrystallized shells of the genus Trochactaeon. Kollmann (Reference Kollmann1967) and Sohl and Kollmann (Reference Sohl and Kollmann1985) discussed the taxonomy of the Trochactaeon group in detail and recognized four subgenera. The subgenus Trochactaeon differs from the other three subgenera by the greater width of the shell in relation to its height and the relatively large anterior expansion of the aperture.
The subgenus Neocylindrites differs from the other subgenera by its high subcylindrical shell and strong whorl overlap. In this regard, it is similar to the subgenus Mexicotrochactaeon, which is distinguished by presence of a parietal fold or ridge in addition to the columellar folds. The incision above the first columellar plate is rather narrow in all subgenera of Trochactaeon, but in the subgenus Mexicotrochactaeon the incision is broad and terminated by the parietal plait. The subgenus Sevanella differs from the other subgenera in having a deep spiral depression in the median part of its whorls and a sculpture of fold-like nodes. They are all heavily nodose shells with laterally depressed whorls. Based on the published descriptions, Trochactaeon (Trochactaeon) is proposed to have evolved from T. (Neocylindrites) during the upper part of the Cenomanian (Sohl and Kollmann, Reference Sohl and Kollmann1985). In general, Cenomanian species such as T. (Trochactaeon) matensis (Fittipaldi, Reference Fittipaldi1900) and T. (Trochactaeon) pseudocylindraceus (Pčelintsev, Reference Pčelintsev1953) remain morphologically close to T. (Neocylindrites), in which the whorls are only slightly inflated (Sohl and Kollmann, Reference Sohl and Kollmann1985).
Pokorný (Reference Pokorný1959) distinguished three subspecies of Trochactaeon giganteus: T. giganteus giganteus (Sowerby, Reference Sowerby1832), T. giganteus ventricosus Hojnos, Reference Hojnos1921, and T. giganteus subglobosus Münster, Reference Münster and Goldfuss1844. In their general review, Kollmann (Reference Kollmann1967) and Sohl and Kollmann (Reference Sohl and Kollmann1985) accepted only two of these: T. giganteus giganteus Sowerby and T. giganteus subglobosus Münster, and included T. giganteus ventricosus in the synonymy of T. giganteus subglobosus. The differences between Trochactaeon giganteus subglobosus and T. giganteus giganteus have been discussed in detail by Kollmann (Reference Kollmann1967). Trochactaeon giganteus giganteus has a large size, but never reaches the width found in Trochactaeon giganteus subglobosus. In large forms of T. giganteus giganteus, the side walls in the upper section of the shell are almost cylindrical adapically and gradually converge in the lower section, whereas in T. giganteus subglobosus they are strongly curved and almost spherical.
Most of the inner whorls have been crushed and a few also have been recrystallized. The Turkish specimens strongly resemble T. (T.) giganteus subglobosus as figured and described by Kollmann (Reference Kollmann1967) from the upper Coniacian to lower Campanian of the Eastern Alps and by Czabalay (Reference Czabalay1973) from the Ugod Formation of Hungary in having the same size, general internal outline, and columellar plications (columellar folds) (Fig. 4). The stratigraphically closest species with similar morphology is Trochactaeon (Trochactaeon) goldfussi (d'Orbigny, 1850) from the Santonian–Campanian Ugod Formation (Czabalay, Reference Czabalay1973, p. 294, pl. 5, fig. 1, pl. 6, fig. 2). This species differs from T. (T.) giganteus subglobosus by having weaker columellar folds.
Discussion
Paleoenvironmental and ecologic significance
Throughout the Cretaceous, several climatic shifts occurred, leading to a transition from a greenhouse to a cool greenhouse world. It is generally agreed that the warmest temperatures in the tropical realm occurred during the Late Cretaceous (Cenomanian–Coniacian), and that temperatures declined sharply during the Maastrichtian (Hay, Reference Hay2008). The Campanian (84.2–72.1 Ma) is the longest stage of the Late Cretaceous and a key interval, encompassing the transition from the ‘hot greenhouse’ of the Middle Cretaceous to the ‘cool greenhouse’ of the Maastrichtian (O'Connor et al., Reference O'Connor, Batenburg, Robinson and Jenkyns2020). Within the tropical belt, zones of slightly hypersaline, nutrient-depleted water facilitated development of the rudist facies and their typical gastropods (Kiel, Reference Kiel and Wagreich2002; Kollmann, Reference Kollmann2014). The life habit of the acteonellid gastropod genus Trochactaeon is generally accepted as mobile epifaunal (Sohl and Kollmann, Reference Sohl and Kollmann1985). A remarkable size increase in the comparatively short time range of this genus may reflect high nutrient environments due to favorable climatic conditions (Kollmann, Reference Kollmann2014).
The Karababa limestone typically consists a patchwork of slightly variant, highly fossiliferous facies, dominated by bio-pelmicrites/sparites (wacke- and packstones), bio-micrites, pellets, and pack- grainstones (Fig. 2.1). Characteristic facies fossils include bivalves (hippuritids and radiolitids), gastropods, echinoid fragments, ostracodes, dasyclad algae, and foraminifers. Hippuritids and radiolitids are usually found as isolated individuals in the Saytepe section (Fig. 2.2). They appear to have toppled over but may have been transported and deposited not far from their original environment. The paleoecological and facies characteristics indicate that the rudist-bearing limestones were deposited on a gently sloping mid-carbonate ramp on the northern shelf of the Arabian platform.
Trochactaeon (T.) giganteus subglobosus formed monospecific populations that lived in the upper part of the Saytepe section. Sanders et al. (Reference Sanders, Kollmann and Wagreich1997) and Kollmann (Reference Kollmann2005) have shown that Trochactaeon inhabited the proximal inner shelf, where it formed mostly monospecific assemblages. The nearly monospecific shell beds of the late Coniacian–early Santonian Trochactaeon assemblage from the Gosau deposits described by Kollmann (Reference Kollmann1965) have been interpreted as deposited in brackish to shallow-marine environments with fluctuating salinity. Salinity is an important issue because acteonellids sometimes have been stated to indicate brackish conditions. Acteonellids and nerineids are reported to have a certain tolerance for reduced salinity (Waite et al., Reference Waite, Wetzel, Meyer and Strasser2008). Trochactaeon (Trochactaeon) presumably had a broader salinity tolerance, ranging from 5–10 ppm to 30–40 ppm (Sohl and Kollmann, Reference Sohl and Kollmann1985).
At most levels, the benthic fauna is impoverished, with diversity values from very low to low. However, low diversity is a result of stress conditions, which can be created by a number of environmental parameters. The low species diversity indicates a somewhat restricted environment (Fürsich et al., Reference Fürsich, Werner and Schneider2009). The low diversity together with individual abundances in the molluscan assemblages are features that are characteristic of brackish-water faunas (Fürsich et al., Reference Fürsich, Freytag, Röhl and Schmid1995). The simple trophic structure of the studied molluscan assemblage with a predominance of T. (T.) giganteus subglobosus may confirm high stress conditions.
The Trochactaeon gastropod fauna can be well correlated with the fauna of the Gosau deposits in Austria, Hungary, and Romania, which occurred in the late Turonian to Campanian (Czabalay, Reference Czabalay1983; Wagreich and Faupl, Reference Wagreich and Faupl1994). Furthermore, associations of Trochactaeon with Radiolites and Nerinea in the Gosau Group at Brandenberg/Tyrol more likely indicate shallow-marine lagoonal conditions (Herm et al., Reference Herm, Kauffman and Wiedmann1979; Kowalke, Reference Kowalke2005). In addition, the Turonian deposits of Amman (Jordan), which are composed of bioclastic limestone with cherty bands and layers, host several acteonellid species including Trochactaeon (Mexicotrochactaeon) marjense Mustafa and Bandel, Reference Mustafa and Bandel1992, and T. (Neocylindrites) lamarcki (Sowerby, Reference Sowerby1832). According to Mustafa and Bandel (Reference Mustafa and Bandel1992), these species are typical of carbonate-dominated lagoonal settings. Hence, tropical lagoonal environments along the coasts of the Tethys Ocean seem to have been the preferred habitat of Acteonellidae (Mustafa and Bandel, Reference Mustafa and Bandel1992). On the other hand, Rashwan et al. (Reference Rashwan, El-Sabbagh, El-Hedeny, Vinn and Mansour2022) interpreted Trochactaeon shell concentrations as shallow nearshore warm-water accumulations in response to storm activity in the upper Turonian of Egypt.
Paleobiogeographical implications
Upper Cretaceous shallow-water carbonates from the Tethys Ocean were accreted in the Mediterranean area and constitute the best archive of the endemic or amphitropical fauna that prevailed in this vast ocean during the Late Cretaceous (Kauffman, Reference Kauffman and Hallam1973; Sohl, Reference Sohl1987; Christensen, Reference Christensen1997; Squires, Reference Squires2011, Reference Squires2018; Steuber et al., Reference Steuber, Scott, Mitchell and Skelton2016). Trochactaeon (Trochactaeon) is a widespread taxon recorded in Europe, Caucasia, Asia, America, south-southwestern Africa, and Arabia–northern Africa (Fig. 5). The stratigraphic range of T. (Trochactaeon) is also remarkable, covering nearly 35 million years from the Cenomanian to Maastrichtian (Fig. 5). The species of the nominate subgenus T. (Trochactaeon) rapidly and continuously diversified from the first appearance in the Cenomanian through Turonian stages (Sohl and Kollmann, Reference Sohl and Kollmann1985). The number of species/subspecies increased regularly from the Cenomanian to the Santonian (Fig. 5), while the highest numbers were reached in the Coniacian and Santonian stages (Sohl and Kollmann, Reference Sohl and Kollmann1985). Diversity decreased further throughout the Campanian and Maastrichtian stages (Kollmann, Reference Kollmann2014).
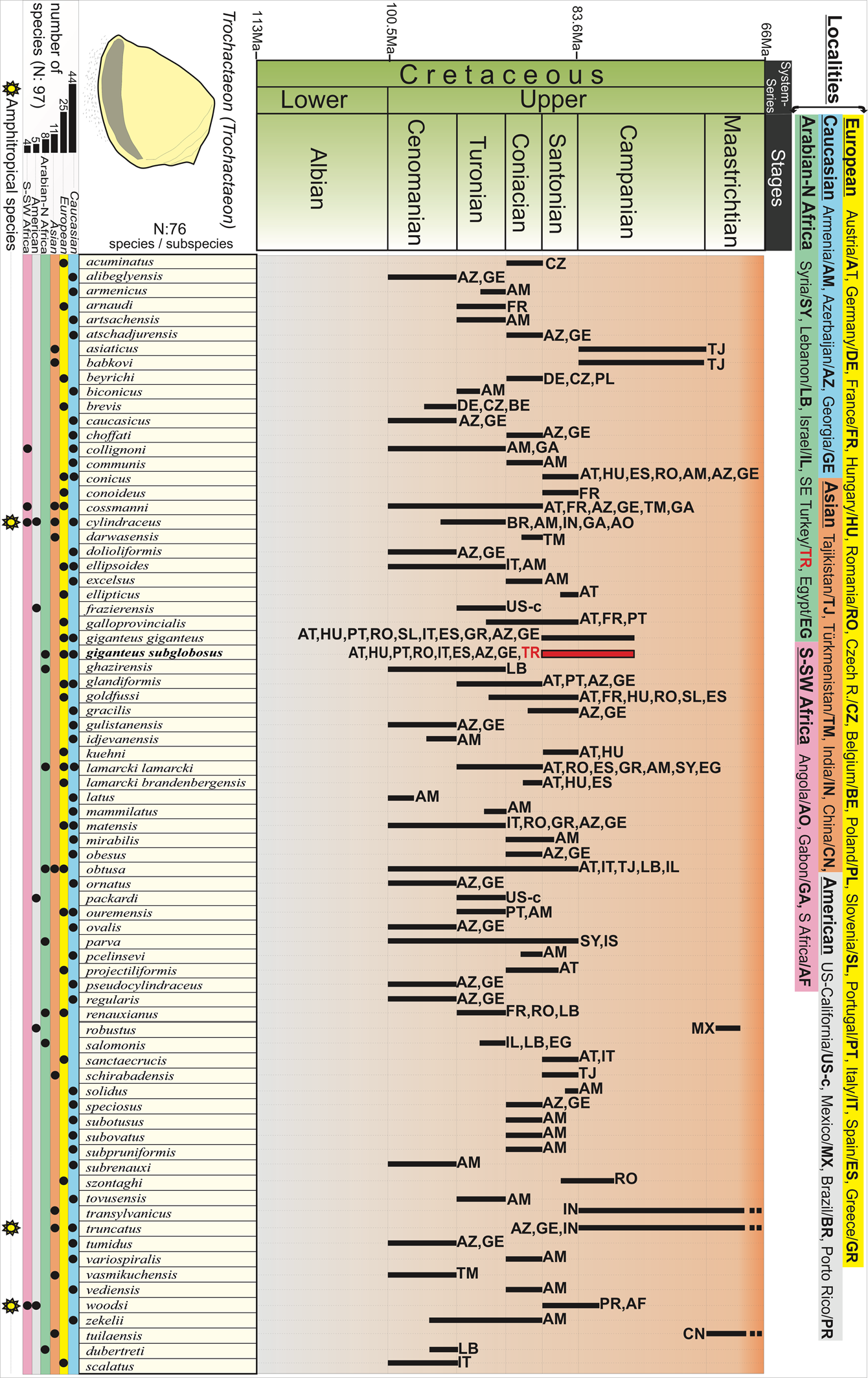
Figure 5. Stratigraphic range of Trochactaeon (Trochactaeon) species and subspecies during the Cretaceous (primary data sources are: Kollmann, Reference Kollmann1967; Czabalay, Reference Czabalay1973; Akopjan, Reference Akopjan1976; Sohl and Kollmann, Reference Sohl and Kollmann1985; Kowalke and Bandel, Reference Kowalke and Bandel1996; Forner and Calzada, Reference Forner and Calzada1998; Kollmann, Reference Kollmann2014).
Acteonellid assemblages have rarely been studied in the Late Cretaceous of the Arabian platform. The presence of T. (Trochactaeon) renauxianus (d'Orbigny, Reference d'Orbigny1842), T. (Trochactaeon) obtuse (Zekeli, Reference Zekeli1852), and T. (Trochactaeon) lamarcki lamarcki in the Cenomanian to Santonian shallow marine carbonate deposits of Syria, Lebanon, Israel, and Egypt extends the paleobiogeographical distribution of the subgenus into the Upper Cretaceous of Arabia–North Africa (Böhm, Reference Böhm1900; Blanckenhorn, Reference Blanckenhorn1927; Delpey, Reference Delpey1940; Sohl and Kollmann, Reference Sohl and Kollmann1985) (Fig. 5). The distribution of T. (T.) giganteus subglobosus was restricted to the Santonian to early Campanian of Austria, Hungary, Portugal, Romania, Italy, Spain, and the Caucasus and is regarded as a good indicator of a prevailing tropical climatic conditions (Pokorný, Reference Pokorný1959; Kollmann, Reference Kollmann1967; Czabalay, Reference Czabalay1973; Sohl and Kollmann, Reference Sohl and Kollmann1985; Forner and Calzada, Reference Forner and Calzada1998) (Fig. 6). The subspecies T. (T.) giganteus subglobosus recorded herein expands the early Campanian distribution of the subspecies to the southern Tethys, which is much farther south than previously reported.

Figure 6. (1) Geographic and (2) paleobiogeographic distributions of Trochactaeon (Trochactaeon) giganteus subglobosus (Münster, Reference Münster and Goldfuss1844). Main ocean currents are based on Poulsen et al. (Reference Poulsen, Barron, Arthur and Peterson2001) and Callapez et al. (Reference Callapez, Gil, Garcia-Hidalgo, Segura, Barroso-Barcenillia and Carenas2015); paleogeographic map after Blakey (Reference Blakey2012).
Trochactaeon (Trochactaeon) species are not recorded from the Coniacian to Santonian of the Americas (Fig. 5). In contrast, they occur in Europe, Caucasia, Asia, southern–southwestern Africa, and Arabia–northern Africa areas during this time (Fig. 5). Amphitropical distribution patterns are relatively common among diverse groups of tropical realm gastropods, particularly in carbonate platforms that were extensive in geographical latitude (i.e., South Africa and North/South America or Caucasia and Africa) during the Late Cretaceous (Squires, Reference Squires2011). Amphitropic Trochactaeon (Trochactaeon) species achieved such a distribution no earlier than the late Cenomanian (Fig. 5).
Cenomanian–Campanian strata are characterized by high biological diversity of the acteonellid, nerineid, oyster, inoceramid, belemnite, and echinoid assemblages (Pčelintsev, Reference Pčelintsev1953; Dobrov and Pavlova, Reference Dobrov, Pavlova and Moskvin1959; Akopjan, Reference Akopjan1976; Sohl and Kollmann, Reference Sohl and Kollmann1985; Christensen, Reference Christensen1997; Mamedalizade, Reference Mamedalizade2019). At that time, several shared species from the Caucasus migrated from northern regions to southern Europe via to the Crimea region. The oceanic circulation pattern within the Mediterranean area was probably dominated by a westward surface current directed toward the central Atlantic along its southern margin, and an eastward surface current along its northern margin (Poulsen et al., Reference Poulsen, Barron, Arthur and Peterson2001; Callapez et al., Reference Callapez, Gil, Garcia-Hidalgo, Segura, Barroso-Barcenillia and Carenas2015). These oceanic currents would have aided dispersal of the acteonellid subgenus Trochactaeon (Trochactaeon) and many other invertebrate groups across the shallow carbonate shelves along the margins of the Tethys (Fig. 6.2).
Conclusion
During the Late Cretaceous, the subgenus Trochactaeon (Trochactaeon) had a very wide geographic distribution. It has been recorded throughout Europe, Caucasia, Asia, North America, and part of the Arabian Plate. Thus, Trochactaeon (Trochactaeon) species or subspecies were mainly a Tethyan Realm subspecies, with a significant extension towards the southern Tethys along the northern Arabian plate margin. The new discovery in Turkey therefore extends the distribution of the subspecies T. (T.) giganteus subglobosus into the southern Tethys in the Late Cretaceous. Preserved specimens of Trochactaeon described herein lie near the top of the Karababa-C member with the rudist fauna suggesting a Campanian age for the Saytepe section. Discovery of T. (T.) giganteus subglobosus in the Karababa Formation greatly expands the paleobiogeographic range of the subspecies and allows us to propose an early Campanian age for the upper part of this formation. Future work on the Karababa Formation offers great potential for the discovery of additional exceptionally preserved fossils in the Upper Cretaceous of SE Turkey that will help further refine the paleobiogeographic and stratigraphic distributions of acteonellid gastropods.
Acknowledgments
Financial support for this study was provided by a national research project of TUBITAK-111Y138. We sincerely thank Çalık Petrol (Oil and Gas Exploration Company, Ankara) for providing us with valuable map data and previous field study. We would also like to thank R.W. Scott and A.M. Murray for discussion and constructive comments, which have helped to improve the paper. Equally, we are grateful to Associate Editor S. Schneider and two anonymous reviewers for all comments and suggestions.
Declaration of competing interests
The authors declare none.






