



Introduction
Shortfin mako shark, Isurus oxyrinchus (Rafinesque, 1810) (Order Lamniformes; Family Lamnidae) has a circumtropical distribution in tropical and temperate seas of the Pacific, Atlantic and Indian Oceans (Compagno et al., Reference Compagno, Dando and Fowler2005). It is a large species (up to 400 cm total length, TL), with oceanic and coastal, as well as epipelagic and mesopelagic habits, extending from the surface to ~4000 m depth (Compagno, 1995; Compagno et al., Reference Compagno, Dando and Fowler2005; Robertson and Allen, Reference Robertson and Allen2015; Gibson et al., Reference Gibson, Streich, Topping and Stunz G2021).
It is the most active and fastest-swimming shark species (Compagno et al., Reference Compagno, Dando and Fowler2005). It feeds on prey including shrimps, squid, a range of fish (ranging from herrings to elasmobranchs and tunas) and marine mammals (Rosas-Luis et al., Reference Rosas-Luis, Pincay-Espinoza, Loor-Andrade, Carrera-Fernández, Kovács and Nagy2016a). It is a species of ecological importance as it may regulate populations of prey species at lower trophic levels.
Isurus oxyrinchus is caught in pelagic longline fisheries occurring in the main ocean basins, where it has often been either a target or marketable bycatch species (Compagno, 1995, Reference Compagno2001). It is also caught in some gillnet fisheries and is one of the most appreciated target species by sports fishermen. Its meat is of high commercial value and is traded fresh, frozen, smoked, salted or desiccated for human consumption. Other body parts (fins, liver, skin and jaws) have also been utilized. Globally, there is a record of landings of 257,811 t during the period 1980–2020 (FAO, 2022). In Ecuador, it is taken in both targeted fisheries and as a bycatch and is captured both by artisanal and industrial fleets. The main landing ports in Ecuador are Manta and Daniel López (Manabí province), Santa Rosa and Anconcito (Santa Elena province). The type of boats used are fiberglass boats (handmade) and wooden mother ships (industrial). The fishing gear used are shallow-set longlines and gillnets. Its meat is used for local consumption and sold fresh or frozen (Martínez-Ortiz and García-Domínguez, Reference Martínez-Ortiz and García-Domínguez2013). During the period 2010–2012, approximately 29,551 t were landed by the artisanal fleet operating from Ecuadorian ports that fish for large pelagic fish (Coello and Herrera, Reference Coello and Herrera2018).
Based on the life-history characteristics of I. oxyrinchus, such as its very slow growth (k = 0.05–0.12), average sexual maturity (females = 273–278 cm LT and males = 180–215 cm LT), long gestation periods between 15 and 18 months, 3-year reproductive cycle and small litter size (4–25 embryos per litter) (Stillwell, Reference Stillwell and Gruber1990; Mollet et al., Reference Mollet, Cliff, Pratt and Stevens2000; Compagno, Reference Compagno2001; Joung and Hsu, Reference Joung and Hsu2005; Ribot-Carballal et al., Reference Ribot-Carballal, Galván-Magana and Quiñónez-Velázquez2005; Conde-Moreno and Galván-Magaña, Reference Conde-Moreno and Galván-Magaña2006; Liu et al., Reference Liu, Sibagariang, Joung and Wang2018), it could be categorized as having a low biological productivity compared to many other elasmobranchs. Most populations of cartilaginous fish can resist low mortality levels before they present symptoms of stock depletion and collapse (Camhi et al., Reference Camhi, Fowler, Musick, Bräutigam and Fordham1998; Musick, Reference Musick and Musick1999; Cortés, Reference Cortés2000), declining faster than other bony fish (Sminkey and Musick, Reference Sminkey and Musick1995, Reference Sminkey and Musick1996). Additionally, the landings observed by the authors in the field during the last 20 years in Ecuador indicate that the fishing exploitation levels could have affected its populations. However, the lack of biological and ecological studies does not allow us to identify its current population status in Ecuadorian waters. Also, it is important to highlight that the International Union for the Conservation of Nature (IUCN) has assessed shortfin mako as Endangered globally, and with decreasing population growth (Rigby et al., Reference Rigby, Barreto, Carlson, Fernando, Fordham, Francis, Jabado, Liu, Marshall, Pacoureau, Romanov, Sherley and Winker2019).
Diet and feeding habit analyses of a species are important to evaluate its ecological role and trophic position in the ecosystem (Allan and Castillo, Reference Allan and Castillo2007). Stomach content analysis of fish provides information on the feeding patterns and the quantitative evaluation of its feeding habits. It is an important aspect for informing ecosystem models and the ecosystem approach to fisheries management. The diet of fish represents the integration of many relevant ecological components, which include behaviour, body condition, habitat use and energy consumption, as well as intraspecific and interspecific interactions (Sagar et al., Reference Sagar, Gop, Nair and Nair2018). The diet analysis of the shortfin mako shark includes information from Yatsu (Reference Yatsu1995), Velasco-Tarelo (Reference Velasco-Tarelo2005), Mucientes-Sandoval and Saborido-Rey (Reference Mucientes-Sandoval and Saborido-Rey2008), Vetter et al. (Reference Vetter, Kohin, Preti, McClatchie and Dewar2008), López et al. (Reference López, Meléndez and Barría2009), Preti et al. (Reference Preti, Soykan, Dewar, Wells, Spear and Kohin2012), Rosas-Luis et al. (Reference Rosas-Luis, Pincay-Espinoza, Loor-Andrade, Carrera-Fernández, Kovács and Nagy2016a), Klarian et al. (Reference Klarian, Canales-Cerro, Barría, Zárate, Concha, Hernández, Heidemeyer, Sallaberry-Pincheira and Meléndez2018) and Márquez-García (Reference Márquez-García2018) in the Pacific Ocean; Stillwell and Kohler (Reference Stillwell and Kohler1982), Vaske-Jr and Ricón-Filho (Reference Vaske-Jr and Ricón-Filho1998), Maia et al. (Reference Maia, Queiroz, Correia and Cabral2006), Wood et al. (Reference Wood, Wetherbee, Juanes, Kohler and Wilga2009), Gorni et al. (Reference Gorni, Loibel, Goitein and Amorim2012, Reference Gorni, Goitein and Amorim2013) and Biton-Porsmoguer et al. (Reference Biton-Porsmoguer, Banaru, Boudouresque, Dekeyser, Viricel and Merchán2015, Reference Biton-Porsmoguer, Bănaru, Boudouresque, Dekeyser, Béarez and Miguez-Lozano2017) in the Atlantic Ocean; and Groeneveld et al. (Reference Groeneveld, Cliff, Dudley, Foulis, Santos and Wintner2014) in the Indian Ocean. However, data for the Tropical Eastern Pacific Ocean are limited.
Based on the importance of food ecology studies and the role played by this shark within tropical marine ecosystems in the Ecuadorian area, it is important to understand the relationship between predator and its prey to know how the flow of nutrients and energy occurs in this type of pelagic habitat. This study aimed to describe the composition of the diet and feeding habits of I. oxyrinchus in the Ecuadorian Pacific Ocean and its relationship with sex and biological cycle phases, as well as to estimate its trophic niche breadth, diet overlap and trophic level.
Materials and methods
Sampling
The fishing port of Santa Rosa is located in the Santa Elena province (02°12’56”S; 80°57’26”W), Ecuador that depends economically on fishing activity (Figure 1). It is the second most important fishing port in the country, after Manta, in number and volume of landings, which include crustaceans, cephalopods, small and large pelagic fishes, sharks, rays and demersal fishes.
Figure 1. Location of the study area (Santa Rosa) in the Province of Santa Elena, Ecuador.
Samples of I. oxyrinchus were collected from Santa Rosa fishing port from May to December 2004 after being landed by the artisanal fishery. The total length (LT), fork length (LF) and precaudal length (LPC) in cm (±0.1 cm) in a natural position were measured, and sex was recorded for each individual. Life stages were characterized through sexual maturity stages that were based on Clark and Von Schmidt (Reference Clark and Von Schmidt1965), Stevens (Reference Stevens1983), Mollet et al. (Reference Mollet, Cliff, Pratt and Stevens2000) and Costa et al. (Reference Costa, Braga, Arfelli and Andamorim2002). Sexual maturity stages were recorded based on the following criteria for females: the shape of the cloaca and whether it was closed or open; courtship marks on the sides of their bodies; the condition of the ovaries, oviducal gland and uterus; and the presence of embryos. For males, the size and condition of claspers (rotation, calcification, opening of the rhipidium and presence of semen); the condition of the testes and seminal vesicle, were examined. The immature individuals were considered juveniles, and mature specimens as adults (Table 1).
Table 1. Number of sampled individuals of the shortfin mako shark, Isurus oxyrinchus, in Santa Rosa, Ecuador, Pacific Ocean, from May to December 2004, by sexes and biological cycle phases

The stomach was also obtained, and its fullness evaluated based on reference values of the scale by Stillwell and Kohler (Reference Stillwell and Kohler1982): 0 (empty), 1 (25% full), 2 (50% full), 3 (75% full) and 4 (100% full). Subsequently, the stomach contents were placed in plastic bags and kept on ice for their transfer to the laboratory to freeze them.
Laboratory analysis
Samples were unfrozen to separate, count, measure (cm) and weigh (g) prey species found in the stomachs. Prey were grouped based on their digestion rate according to Olson and Galván-Magaña (Reference Olson and Galván-Magaña2002): state 1 (individuals that present all the complete morphological characteristics and are easily identifiable), state 2 (organisms without skin and eyes, and with muscle exposure), state 3 (headless specimens, some body parts present and axial skeleton) and state 4 (otoliths, skeletons and squid beaks).
The identification of prey items found in the stomachs of I. oxyrinchus was carried out based on specialized guides. The crustacean species were identified by their exoskeleton according to Hendrickx (Reference Hendrickx, Fischer, Krupp, Schneider, Sommer, Carpenter and Niem1995). The cephalopod species were identified using their mandible (‘beaks’) and bodies according to Ingrid et al. (Reference Ingrid, Iverson and Pinkas1971), Wolff (Reference Wolff1982, Reference Wolff1984), Clarke (Reference Clarke1986), Roper et al. (Reference Roper, Sweeney, Hochberg, Fischer, Krupp, Schneider, Sommer, Carpenter and Niem1995), Jereb and Roper (Reference Jereb and Roper2010) and Jereb et al. (Reference Jereb, Roper, Norman and Finn2014). The fish species (i.e. complete organisms, skeletons and otoliths) were identified following Clothier (Reference Clothier1950), Clothier and Baxter (Reference Clothier and Baxter1969), Fischer et al. (Reference Fischer, Krupp, Schneider, Sommer, Carpenter and Niem1995), García-Godos (Reference García-Godos2001), Jiménez and Béarez (Reference Jiménez and Béarez2004), Muñoz (Reference Muñoz2012), Robertson and Allen (Reference Robertson and Allen2015) and Vinueza (Reference Vinueza2015). The marine mammal species were identified according to Jefferson and Leatherwood (Reference Jefferson, Leatherwood, Fischer, Krupp, Schneider, Sommer, Carpenter and Niem1995).
Data analysis
The LT of females and males were compared through the non-parametric Mann–Whitney test (W) to investigate if there were significant differences between them. The same test was used for the comparison of the diet by sex and biological cycle phases (Daniel, Reference Daniel1991; Celis-De la Rosa and Labrada-Martagón, Reference Celis-De la Rosa and Labrada-Martagón2014). Organisms were grouped according to their biological cycle phases, regardless of their sex. In addition, length–frequency distributions were made by sex, using 10 cm length groups (modified from Holden and Raitt, Reference Holden and Raitt1975).
The number of stomachs analysed to adequately describe the diet of this species was established using the Pielou method (Hoffman, Reference Hoffman, Lipovsky and Simenstad1979). Likewise, the coefficient of variation was obtained for each stomach and plotted as a secondary axis; such coefficient was obtained by the relationship between the standard deviation and the average diversity of prey. For this purpose, the number of stomachs was estimated through the EstimateS V.8.0 software (Colwell, Reference Colwell2019), in which the number of stomachs analysed was subjected to 100 permutations to eliminate bias with α = 0.05. The selection of the respective stomach number was the one whose variation was observed to be 0.05.
The ecological indices used were the numeric methods (%N) of frequency of occurrence (%FOi) and gravimetry (%W) (Hyslop, Reference Hyslop1980). The importance that each species contributed to the diet of I. oxyrinchus was evaluated through the Prey-Specific Index of Relative Importance (%PSIRI) (Brown et al., Reference Brown, Bizzarro, Cailliet and Ebert2011):

where %PNi = prey-specific numeric abundance and %PWi = prey-specific weight abundance. These parameters were obtained from the following models:


The arrangement of the taxonomic order of the species consumed by the shortfin mako shark was completed based on Young et al. (Reference Young, Vecchione and Mangold2019) for cephalopods and Van der Laan et al. (Reference Van der Laan, Fricke and Eschmeyer2022) for bony fish.
The amplitude of the predator's diet was calculated through the Levin Index (Bi) (Krebs, Reference Krebs1985):

where Pij = proportion of prey j in the predator's diet i and n = number of prey species. The values of this index range from 0 to 1. Values lower than 0.6 indicate that the diet is dominated by few prey items, and thus, it would be a specialist consumer; while values equal to or greater than 0.6 suggest that consumers are generalists (Labropoulou and Eleftheriou, Reference Labropoulou and Eleftheriou1997).
The trophic overlap analysis (degree of food resource distribution) was estimated between sexes and maturity states through the Jaccard index (Krebs, Reference Krebs1999):

where J = Jaccard index, the same that employs the presence/absence of common prey in predators; a = number of common prey species in predators of both sexes or biological cycle phases; b = number of exclusive prey species of a predator (females or juvenile individuals); and c = number of exclusive prey species of the other predator (males or adult specimens). Values close to 0 indicate there is no overlap, while those close to 1 suggest that the use of food resources is identical. The program Past V.4.01 was used to determine this index (Hammer, Reference Hammer2020).
The trophic level was calculated from the equation proposed by Cortés (Reference Cortés1999):

where TLk = trophic level of the predator, n = number of prey species, Pjx = relative proportion of prey items that conform the consumer's diet and TLj = trophic level of the prey. The trophic levels of the prey consumed employed in the TLk estimates of I. oxyrinchus were obtained from Cortés (Reference Cortés1999) and Froese and Pauly (Reference Froese and Pauly2022). To categorize the trophic level of predators and prey, the scale proposed by Odum (Reference Odum1971) was used: TL-I: primary producers, TL-II: herbivores (fed on level I), TL-III: primary carnivores (consumed level II), TL-IV: secondary carnivores (ingested level III) and TL-V: tertiary carnivores (fed on level IV).
Given that some earlier studies had described the diet of I. oxyrinchus in relation to LF or LPC, the following length conversion relationships were used for comparison with LT:



Results
Characteristics of the sample
A total of 142 individuals were analysed, comprising 81 females (57.0%) and 61 males (43.0%). Females ranged from 104 to 295 cm LT ($\bar{x}$ = 170.4 ± 32.2), while males ranged from 127 to 245 cm LT ($\bar{x}$
= 170.4 ± 32.2), while males ranged from 127 to 245 cm LT ($\bar{x}$ = 171.7 ± 25.9). The largest female was 50 cm larger than the largest male but no significant differences were observed by sex (W = 5872; p > 0.05) (Figure 2). Furthermore, 118 specimens (83.1%) were juveniles and 24 (16.9%) were adults. Adult individuals were observed from 181 cm LT onwards (males) and 216 cm LT onwards (females).
= 171.7 ± 25.9). The largest female was 50 cm larger than the largest male but no significant differences were observed by sex (W = 5872; p > 0.05) (Figure 2). Furthermore, 118 specimens (83.1%) were juveniles and 24 (16.9%) were adults. Adult individuals were observed from 181 cm LT onwards (males) and 216 cm LT onwards (females).

Figure 2. Size frequency distribution of the shortfin mako shark, Isurus oxyrinchus, landed in the Santa Rosa fishery port from May to December 2004.
Of the 142 stomachs analysed, 106 contained food (75%) and 36 were empty (25%). Approximately half of the studied stomachs were in state 1 (n = 67; 47.2%), followed by state 0 (n = 36; 25.4%), state 2 (n = 18; 12.7%), state 4 (n = 13; 9.2%) and state 3 (n = 8; 5.6%). Regarding the state of digestion of prey that composed the diet (n = 289), more than half were in a completely digested state (state 4; n = 176; 60.9%), followed by state 3 (advanced digestion; n = 87; 30.1%), and digestion states 1 and 2 (n = 13; 4.5%).
Based on the accumulative prey curve, the number of stomachs analysed in this study was sufficient to characterize the diet of I. oxyrinchus, given that the asymptote was reached at stomach 69. The variability of prey items decreased noticeably from stomach 54 (Figure 3).

Figure 3. General cumulative curve of the prey species consumed by the shortfin mako shark, Isurus oxyrinchus, in Santa Rosa, Province of Santa Elena, Ecuador, Pacific Ocean. Arrow indicates the number of stomachs where the curve reached the asymptote.
Food composition
A total of 24 prey species or food components were identified, including shrimp remains, 12 cephalopods, 10 bony fish and one marine mammal. A total of 289 individuals that composed the diet of I. oxyrinchus were counted and weighed 392,079.46 g. According to the PSIRI (%), the most relevant prey species were Humboldt squid, Dosidicus gigas (42.57%); purpleback squid, Sthenoteuthis oualaniensis (21.04%); and halfbeak fish from the family Hemiramphidae (11.85%) (Table 2).
Table 2. Prey consumed by the shortfin mako shark, Isurus oxyrinchus, and its respective indices

N, number; %N, percentage of number; %Ni, percentage of prey-specific abundance; FOi, frequency of occurrence; %FOi, percentage of frequency of occurrence; W, weight; %W, percentage of weight; %Wi, percentage of prey-specific weight; %PSIRI, percentage of prey-specific index of relative importance. The taxonomic order of the prey species is based on Young et al. (Reference Young, Vecchione and Mangold2019) for cephalopods and Van der Laan et al. (Reference Van der Laan, Fricke and Eschmeyer2022) for fish.
With regards to feeding by sex, the diet of females was composed of 20 species distributed in 157 individuals with a total mass of 219,332.19 g, which included shrimp remains, ten squids, eight teleosts and a marine mammal. The dominant prey species according to the PSIRI (%) were D. gigas (40.67%), S. oualaniensis (23.61%) and halfbeaks (13.34%). Likewise, males had a trophic spectrum of 17 species represented by 132 specimens with a total weight of 172,117.93 g. The most relevant prey based on the PSIRI (%) were also D. gigas (44.93%), S. oualaniensis (17.79%) and halfbeaks (9.97%) (Figure 4). There were no significant differences between the composition of the diet of I. oxyrinchus by sex (W = 589; p > 0.05).
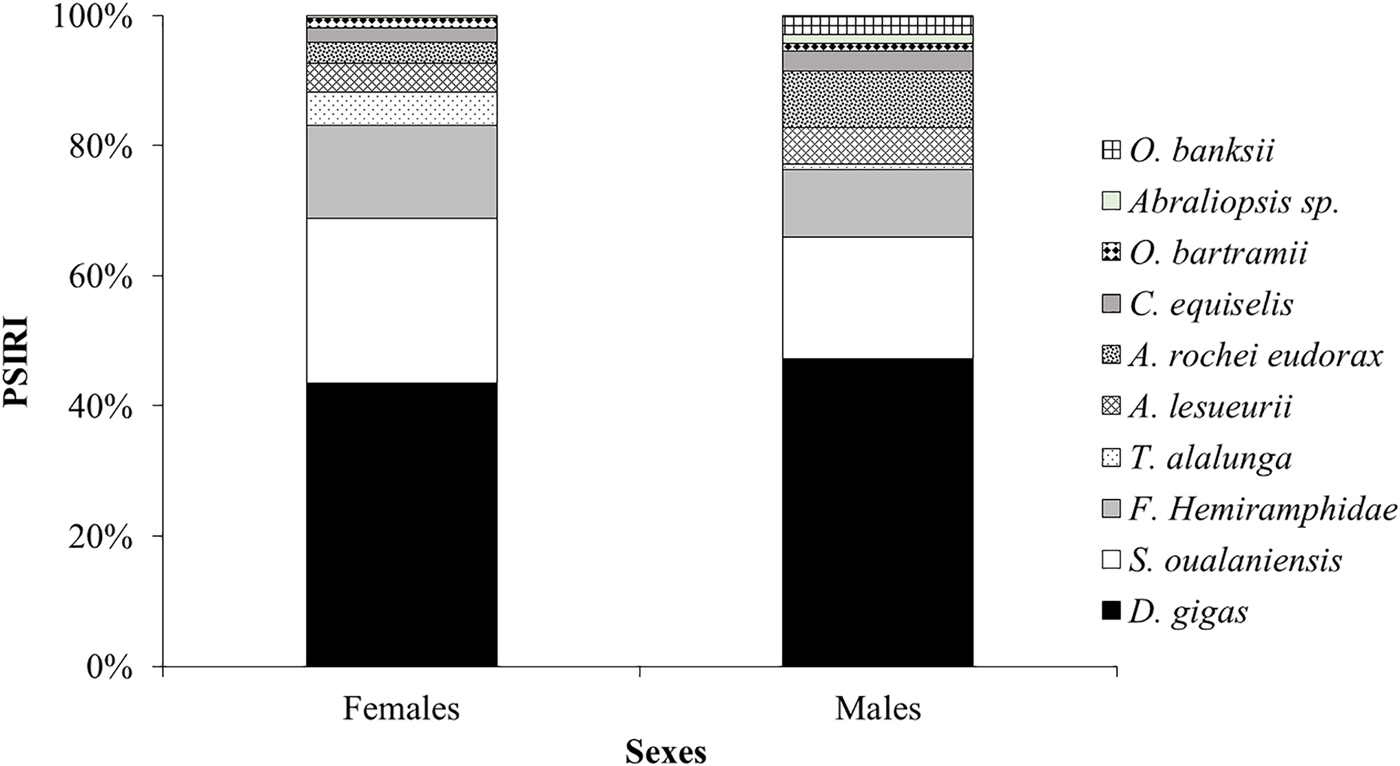
Figure 4. Values of prey-specific index of relative importance (%PSIRI) for females and males of Isurus oxyrinchus for each main species and others that compound its diet in the Ecuadorian Pacific Ocean.
The diet of juvenile sharks was comprised of 22 species distributed in one shrimp, ten cephalopods, ten bony fish and one marine mammal, equal to 244 individuals with a total weight of 307,335.35 g. The most abundant species according to the PSIRI (%) were D. gigas (42.78%), S. oualaniensis (19.08%) and halfbeaks (12.43%). Adult sharks had a trophic spectrum of 12 species, including eight squid and four teleosts represented by 46 specimens with a total weight of 82,892.92 g. The most relevant species based on the PSIRI (%) were D. gigas (42.49%) and S. oualaniensis (30.54%) (Figure 5). No significant differences were registered between the diet of I. oxyrinchus by life stages (W = 648; p > 0.05).
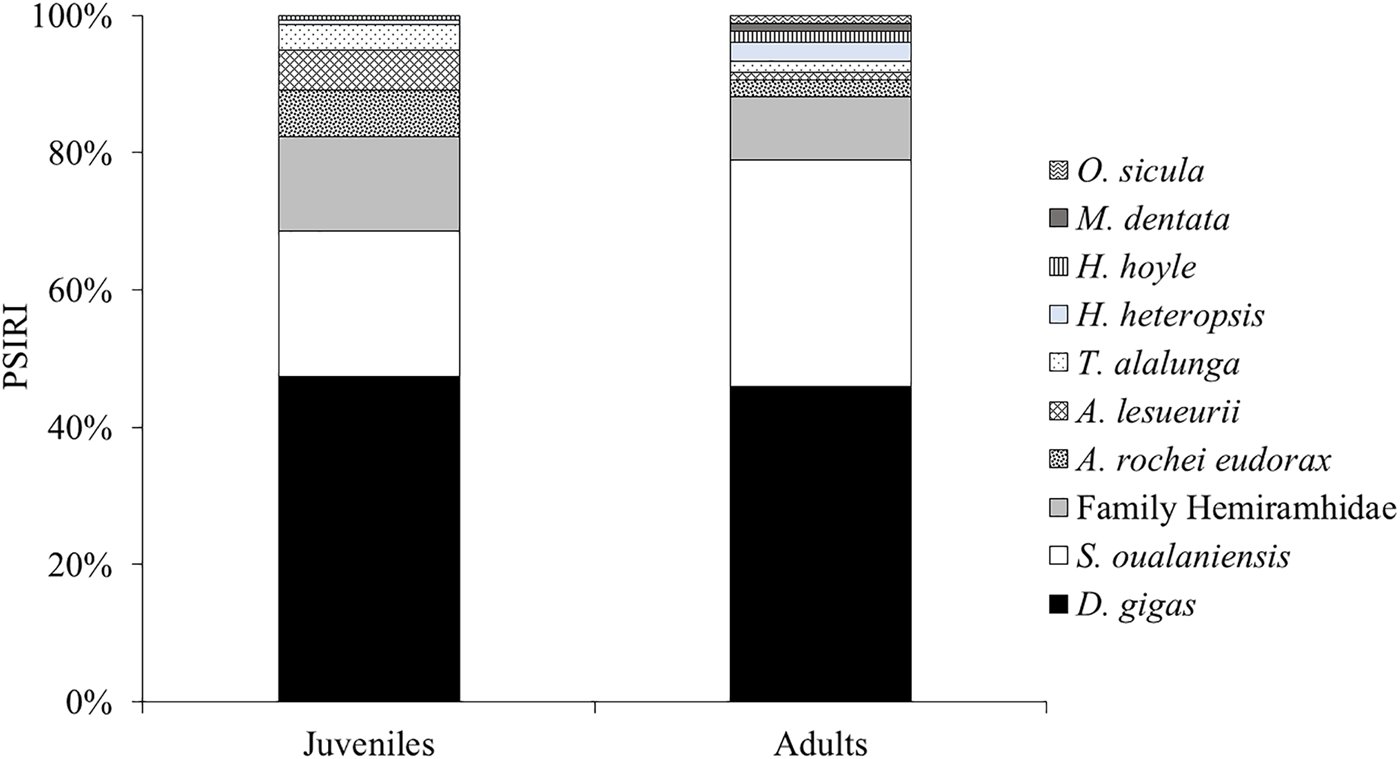
Figure 5. Values of prey-specific index of relative importance (%PSIRI) of juvenile and adult individuals of Isurus oxyrinchus for each main species and others that compound its trophic spectrum in waters of Ecuador.
Feeding habits
Breadth of trophic niche. Isurus oxyrinchus was a specialist predator that preferred three prey species out of the available 24 prey (Bi = 0.25). This behaviour remained the same in the rest of the analysed categories. Males (Bi = 0.34) presented a slightly higher value than females (Bi = 0.29). Similarly, adults (Bi = 0.37) registered a higher value than juveniles (Bi = 0.27).
Trophic overlap. The trophic overlap was moderate between females and males (J = 0.54), as well as for juveniles and adults (J = 0.42), that is, their food components were consumed regardless of their sex or life stages, so there was no defined pattern in the use of food resources.
Trophic level. The trophic level for this species was TLk = 4.47, which indicated that I. oxyrinchus was a top predator of the secondary carnivore type. The trophic levels by sex (TLk = 4.46 in females and TLk = 4.49 for males), as well as by biological cycle phases (TLk = 4.49 in juvenile sharks and 4.40 in adults) were similar.
Discussion
Food composition
The prey observed in this study included cephalopods, teleosts and marine mammals, with a high preference for squid, which agrees with the findings of previous studies. The diet of I. oxyrinchus along Ecuadorian Pacific Ocean has been poorly studied (Pincay-Espinoza, Reference Pincay-Espinoza2014; Rosas-Luis et al., Reference Rosas-Luis, Pincay-Espinoza, Loor-Andrade, Carrera-Fernández, Kovács and Nagy2016a, Reference Rosas-Luis, Navarro, Loor-Andrade and Forero2017). However, some trophic studies in the Pacific and other oceans suggest that this species is piscivorous (Stillwell and Kohler, Reference Stillwell and Kohler1982; Vaske-Jr and Ricón-Filho, Reference Vaske-Jr and Ricón-Filho1998; Maia et al., Reference Maia, Queiroz, Correia and Cabral2006; López et al., Reference López, Meléndez and Barría2009; Wood et al., Reference Wood, Wetherbee, Juanes, Kohler and Wilga2009; Preti et al., Reference Preti, Soykan, Dewar, Wells, Spear and Kohin2012; Biton-Porsmoguer et al., Reference Biton-Porsmoguer, Banaru, Boudouresque, Dekeyser, Viricel and Merchán2015, Reference Biton-Porsmoguer, Bănaru, Boudouresque, Dekeyser, Béarez and Miguez-Lozano2017; Klarian et al., Reference Klarian, Canales-Cerro, Barría, Zárate, Concha, Hernández, Heidemeyer, Sallaberry-Pincheira and Meléndez2018) and teuthophagous (Mucientes-Sandoval and Saborido-Rey, Reference Mucientes-Sandoval and Saborido-Rey2008; Vetter et al., Reference Vetter, Kohin, Preti, McClatchie and Dewar2008; Gorni et al., Reference Gorni, Goitein and Amorim2013; Rosas-Luis et al., Reference Rosas-Luis, Loor-Andrade, Carrera-Fernández, Pincay-Espinoza, Vinces-Ortega and Chompoy-Salazar2016b). Such observations suggest that the diet of I. oxyrinchus is related to geographic location, oceanic productivity and prey availability.
Even though I. oxyrinchus had a high preference for specific groups of prey (e.g. fish and/or cephalopods), it presents a wide trophic spectrum composed of birds (López et al., Reference López, Meléndez and Barría2009; Rosas-Luis et al., Reference Rosas-Luis, Pincay-Espinoza, Loor-Andrade, Carrera-Fernández, Kovács and Nagy2016a), marine mammals (Stillwell and Kohler, Reference Stillwell and Kohler1982; Mucientes-Sandoval and Saborido-Rey, Reference Mucientes-Sandoval and Saborido-Rey2008; López et al., Reference López, Meléndez and Barría2009; Wood et al., Reference Wood, Wetherbee, Juanes, Kohler and Wilga2009; Preti et al., Reference Preti, Soykan, Dewar, Wells, Spear and Kohin2012; Biton-Porsmoguer et al., Reference Biton-Porsmoguer, Banaru, Boudouresque, Dekeyser, Viricel and Merchán2015, Reference Biton-Porsmoguer, Bănaru, Boudouresque, Dekeyser, Béarez and Miguez-Lozano2017; Rosas-Luis et al., Reference Rosas-Luis, Pincay-Espinoza, Loor-Andrade, Carrera-Fernández, Kovács and Nagy2016a; Klarian et al., Reference Klarian, Canales-Cerro, Barría, Zárate, Concha, Hernández, Heidemeyer, Sallaberry-Pincheira and Meléndez2018), sea turtles (Biton-Porsmoguer et al., Reference Biton-Porsmoguer, Banaru, Boudouresque, Dekeyser, Viricel and Merchán2015, Reference Biton-Porsmoguer, Bănaru, Boudouresque, Dekeyser, Béarez and Miguez-Lozano2017), other elasmobranchs (Applegate, Reference Applegate1966; Bass et al., Reference Bass, D'Aubrey and Kistnasamy1975; Stillwell and Kohler, Reference Stillwell and Kohler1982; Maia et al., Reference Maia, Queiroz, Correia and Cabral2006; López et al., Reference López, Meléndez and Barría2009; Wood et al., Reference Wood, Wetherbee, Juanes, Kohler and Wilga2009; Groeneveld et al., Reference Groeneveld, Cliff, Dudley, Foulis, Santos and Wintner2014; Rosas-Luis et al., Reference Rosas-Luis, Pincay-Espinoza, Loor-Andrade, Carrera-Fernández, Kovács and Nagy2016a) and crustaceans (Maia et al., Reference Maia, Queiroz, Correia and Cabral2006; Mucientes-Sandoval and Saborido-Rey, Reference Mucientes-Sandoval and Saborido-Rey2008; López et al., Reference López, Meléndez and Barría2009; Wood et al., Reference Wood, Wetherbee, Juanes, Kohler and Wilga2009; Biton-Porsmoguer et al., Reference Biton-Porsmoguer, Banaru, Boudouresque, Dekeyser, Viricel and Merchán2015, Reference Biton-Porsmoguer, Bănaru, Boudouresque, Dekeyser, Béarez and Miguez-Lozano2017; Rosas-Luis et al., Reference Rosas-Luis, Pincay-Espinoza, Loor-Andrade, Carrera-Fernández, Kovács and Nagy2016a). This wide trophic spectrum indicates that the use of habitat behaves similarly regardless of its geographic area. Despite this, the selection of prey depends, to a large extent, on their availability (López et al., Reference López, Meléndez and Barría2009).
The similarity in the feeding behaviour of I. oxyrinchus in the different study areas could be related to habitat changes as a result of large vertical migrations (Wootton, Reference Wootton1990; López et al., Reference López, Meléndez and Barría2009), which can reach depths of up to 800 m during the day (Bress, Reference Bress1993; Loefer et al., Reference Loefer, Sedberry and McGovern2005; Field et al., Reference Field, Baltz, Philips and Walker2007; Vetter et al., Reference Vetter, Kohin, Preti, McClatchie and Dewar2008; Abascal et al., Reference Abascal, Quintans, Ramos-Cartelle and Mejuto2011). During this time, the species shows a high preference for depths below 300 m. Conversely, this species is located between the surface and 300 m of depth during the night (Vetter et al., Reference Vetter, Kohin, Preti, McClatchie and Dewar2008).
The bathymetric preferences of I. oxyrinchus indicate that this species has mesopelagic habits, which is supported by its high preference for the consumption of squids of the family Ommastrephidae (Stillwell and Kohler, Reference Stillwell and Kohler1982; Vetter et al., Reference Vetter, Kohin, Preti, McClatchie and Dewar2008; López et al., Reference López, Meléndez and Barría2009; Preti et al., Reference Preti, Soykan, Dewar, Wells, Spear and Kohin2012; Rosas-Luis et al., Reference Rosas-Luis, Pincay-Espinoza, Loor-Andrade, Carrera-Fernández, Kovács and Nagy2016a, Reference Rosas-Luis, Loor-Andrade, Carrera-Fernández, Pincay-Espinoza, Vinces-Ortega and Chompoy-Salazar2016b; this study). These squids occur from the surface to bathypelagic zones (0–2000 m of depth), with a high nocturnal activity between the surface and 200 m (Jereb and Roper, Reference Jereb and Roper2010). Additionally, it consumes epipelagic, mesopelagic and bathypelagic fish such as longfin cubehead, Cubiceps pauciradiatus, which is distributed vertically from 50 to 1000 m (Cervigón, Reference Cervigón1994; López et al., Reference López, Meléndez and Barría2009); escolar, Lepidocybium flavobrunneum (200–1000 m depth; Shcherbachev, Reference Shcherbachev1987; Riede, Reference Riede2004; Gorni et al., Reference Gorni, Goitein and Amorim2013); Atlantic saury, Scomberesox saurus (0–30 m depth; Wisner, Reference Wisner, Quero, Hureau, Karrer, Post and Saldanha1990; Biton et al., Reference Biton-Porsmoguer, Banaru, Boudouresque, Dekeyser, Viricel and Merchán2015); bluefish, Pomatomus saltatrix (0–200 m depth; Stillwell and Kohler, Reference Stillwell and Kohler1982; FAO-FIGIS, 2005); and snake mackerel, Gempylus serpens (0–600 m depth; Cervigón, Reference Cervigón1994; Mucientes-Sandoval and Saborido-Rey, Reference Mucientes-Sandoval and Saborido-Rey2008; McMillan et al., Reference McMillan, Griggs, Francis, Marriott, Paul, Mackay, Wood and Sui Hand Wei2011). Thus, the depth ranges of the prey consumed by I. oxyrinchus coincide with the depths that this species frequents at different times of the day, allowing for prey selection and habitat use.
Furthermore, ontogenic shifts are influenced by habitat use, which is a characteristic documented in other shark species such as sickle fin smooth-hound, Mustelus lunulatus (Méndez-Macías et al., Reference Méndez-Macías, Velázquez-Chiquito, Estupiñán-Montaño and Galván-Magaña2019); blue shark, Prionace glauca (Estupiñán-Montaño et al., 2019); scalloped hammerhead, Sphyrna lewini (Estupiñán-Montaño et al., Reference Estupiñán-Montaño, Galván-Magaña, Elorriaga-Verplancken, Zetina-Rejón, Sánchez-González, Polo-Silva, Villalobos-Ramírez, Rojas-Cundumí and Delgado-Huertas2021a, Reference Estupiñán-Montaño, Tamburin and Delgado-Huertas2021b); I. oxyrinchus (Maia et al., Reference Maia, Queiroz, Correia and Cabral2006; Mucientes-Sandoval and Saborido-Rey, Reference Mucientes-Sandoval and Saborido-Rey2008; Preti et al., Reference Preti, Soykan, Dewar, Wells, Spear and Kohin2012; Malpica-Cruz et al., Reference Malpica-Cruz, Herzka, Sosa-Nishizaki and Escobedo-Olvera2013; Klarian et al., Reference Klarian, Canales-Cerro, Barría, Zárate, Concha, Hernández, Heidemeyer, Sallaberry-Pincheira and Meléndez2018; Tamburín et al., Reference Tamburín, Kim, Elorriaga-Verplancken, Madigan, Hoyos-Padilla, Sánchez-González, Hernández-Herrera, Castillo-Géniz, Godinez-Padilla and Galván-Magaña2019); and I. paucus (Estupiñán-Montaño and Delgado-Huertas, Reference Estupiñán-Montaño and Delgado-Huertas2022). This condition can be explained by the results obtained in a stable isotope study at different maturity stages in shortfin mako shark (Tamburín et al., Reference Tamburín, Kim, Elorriaga-Verplancken, Madigan, Hoyos-Padilla, Sánchez-González, Hernández-Herrera, Castillo-Géniz, Godinez-Padilla and Galván-Magaña2019). These latter authors found that young-year-old embryos and juveniles of I. oxyrinchus (80–100 cm LT) show isotopic signals of oceanic origin as a product of maternal transfer processes. Moreover, the study showed that larger individuals (>100 cm LT) increase the consumption of oceanic prey, suggesting this species moves from coastal to oceanic areas changing habitats as it grows (Tamburín et al., Reference Tamburín, Kim, Elorriaga-Verplancken, Madigan, Hoyos-Padilla, Sánchez-González, Hernández-Herrera, Castillo-Géniz, Godinez-Padilla and Galván-Magaña2019), where it consumes larger prey with a higher caloric intake (Pope et al., Reference Pope, Brown, Duffy and Michaletz2001).
Similar to other studies, the low incidence of dolphins observed in the present study demonstrates a low preference for this prey group in the diet of I. oxyrinchus (Wood et al., Reference Wood, Wetherbee, Juanes, Kohler and Wilga2009; Biton-Porsmoguer et al., Reference Biton-Porsmoguer, Banaru, Boudouresque, Dekeyser, Viricel and Merchán2015, Reference Biton-Porsmoguer, Bănaru, Boudouresque, Dekeyser, Béarez and Miguez-Lozano2017; Rosas-Luis et al., Reference Rosas-Luis, Pincay-Espinoza, Loor-Andrade, Carrera-Fernández, Kovács and Nagy2016a; Klarian et al., Reference Klarian, Canales-Cerro, Barría, Zárate, Concha, Hernández, Heidemeyer, Sallaberry-Pincheira and Meléndez2018). Therefore, its intake could rather be the product of an opportunistic strategy by this predator (Maia et al., Reference Maia, Queiroz, Correia and Cabral2006; Mucientes-Sandoval and Saborido-Rey, Reference Mucientes-Sandoval and Saborido-Rey2008).
Feeding habits
A total of 24 prey species composed the I. oxyrinchus’ diet in the Ecuadorian Pacific. This is similar to previous studies that describe the diet of this species with a trophic spectrum of 17–24 prey items (Mucientes-Sandoval and Saborido-Rey, Reference Mucientes-Sandoval and Saborido-Rey2008; López et al., Reference López, Meléndez and Barría2009; Biton-Porsmoguer et al., Reference Biton-Porsmoguer, Bănaru, Boudouresque, Dekeyser, Béarez and Miguez-Lozano2017; this study). Furthermore, the feeding behaviour of I. oxyrinchus observed in the present study is of a specialist type for all the categories analysed, due to the preferential consumption of D. gigas, S. oualaniensis and halfbeaks (Hemiramphidae). This resembles to that reported by other authors, which the feeding strategy of this species is considered specialist (Field et al., Reference Field, Baltz, Philips and Walker2007; Zeidberg and Robinson, Reference Zeidberg and Robison2007; Lopez et al., Reference Lopez, Barría and Meléndez2012), but as somewhat opportunistic and/or generalist in some cases (Velasco-Talero, Reference Velasco-Tarelo2005; López et al., Reference López, Meléndez and Barría2009; Maia et al., Reference Maia, Queiroz, Correia and Cabral2006; Pincay-Espinoza, Reference Pincay-Espinoza2014).
It is important to highlight the dominance of D. gigas in the I. oxyrinchus’ diet may be a response to the population increase of this squid species throughout the EPO (Field et al., Reference Field, Baltz, Philips and Walker2007; Zeidberg and Robinson, Reference Zeidberg and Robison2007). Therefore, it is evident that I. oxyrinchus’ foraging behaviour would be conditioned to (i) the migratory nature that allows it to forage in different types of habitats (Wootton, Reference Wootton1990; López et al., Reference López, Meléndez and Barría2009; Gibson et al., Reference Gibson, Streich, Topping and Stunz G2021); (ii) the availability of prey and its abundance (López et al., Reference López, Meléndez and Barría2009; Pincay-Espinoza, Reference Pincay-Espinoza2014) related to spatiotemporal variability (Maia et al., Reference Maia, Queiroz, Correia and Cabral2006; Mucientes-Sandoval and Saborido-Rey, Reference Mucientes-Sandoval and Saborido-Rey2008; Pincay-Espinoza, Reference Pincay-Espinoza2014; Biton-Porsmoguer et al., Reference Biton-Porsmoguer, Banaru, Boudouresque, Dekeyser, Viricel and Merchán2015). For example, in North America, a high contribution of Pacific saury, Cololabis saira, in the diet of I. oxyrinchus has been recorded, which has been associated to the fact that this shark takes advantage of the high aggregations of C. saira during their reproductive season, given this species uses the California current to lay its eggs (Froese and Pauly, Reference Froese and Pauly2022); therefore, I. oxyrinchus takes advantage of this phenomenon to obtain easy and very abundant food (Juanes et al., Reference Juanes, Hare and Miskiewicz1996; Salerno et al., Reference Salerno, Burnett and Ibara2001). Also, ontogenetic changes may occur associated with prey consumption and habitat use (Maia et al., Reference Maia, Queiroz, Correia and Cabral2006; Mucientes-Sandoval and Saborido-Rey, Reference Mucientes-Sandoval and Saborido-Rey2008; Preti et al., Reference Preti, Soykan, Dewar, Wells, Spear and Kohin2012; Malpica-Cruz et al., Reference Malpica-Cruz, Herzka, Sosa-Nishizaki and Escobedo-Olvera2013; Klarian et al., Reference Klarian, Canales-Cerro, Barría, Zárate, Concha, Hernández, Heidemeyer, Sallaberry-Pincheira and Meléndez2018; Tamburín et al., Reference Tamburín, Kim, Elorriaga-Verplancken, Madigan, Hoyos-Padilla, Sánchez-González, Hernández-Herrera, Castillo-Géniz, Godinez-Padilla and Galván-Magaña2019). This information would explain the specialist, opportunistic and/or generalist nature of this species throughout its distribution.
The diet analysis by sex and biological cycle phases indicated that there was a moderate similarity between categories, suggesting a certain degree of food competition between them. However, these estimates differ from that reported by other authors which report high values of similarity indices, Cλ Sexes = 0.97 (Maia et al., Reference Maia, Queiroz, Correia and Cabral2006); 0.87 (Pincay-Espinoza, Reference Pincay-Espinoza2014) and Cλ Maturity stages = 0.71–0.98 (Maia et al., Reference Maia, Queiroz, Correia and Cabral2006); 0.52–0.85 (Pincay-Espinoza, Reference Pincay-Espinoza2014).
The different estimates of trophic overlap obtained in this and other studies could indicate that I. oxyrinchus have segregation processes related to the habitat use and behaviour (Wearmouth and Sims, Reference Wearmouth and Sims2008), which would be explained by the greater trophic spectrum of females (20 prey species) compared to that of males (17). Therefore, we hypothesize that female I. oxyrinchus could prefer to use oceanic and coastal areas, potentially then interacting with more demersal prey in coastal areas, whereas males would prefer to use oceanic areas. This habitat use by the sexes of I. oxyrinchus could be conditioned by the availability and abundance of prey in the area, environmental and oceanographic factors, among others. Therefore, more studies are needed to validate this hypothesis, because of that, these results should be interpreted with caution.
Sharks are generally considered top predators in marine ecosystems, although this characterization is not always correct for all shark species, since their functional role in ecosystems is conditioned by factors such as maximum body length, geographic distribution, trophic breadth, vulnerability to predation by other sharks (Roff et al., Reference Roff, Doropoulos, Rogers, Bozec, Krueck, Aurellado, Priest, Birrell and Mumby2016) and feeding habits (Cortés, Reference Cortés1999). Based on the last aspect, stomach content and stable isotope studies have estimated that I. oxyrinchus occupies medium and high trophic positions between 3.60 and 4.96 (Cortés, Reference Cortés1999; Estrada et al., Reference Estrada, Rice, Lutcavage and Skomal2003; Rosas-Luis et al., Reference Rosas-Luis, Pincay-Espinoza, Loor-Andrade, Carrera-Fernández, Kovács and Nagy2016a; Biton-Porsmoguer et al., Reference Biton-Porsmoguer, Bănaru, Boudouresque, Dekeyser, Béarez and Miguez-Lozano2017).
The trophic level for I. oxyrinchus indicated that this species was a top predator of the secondary carnivore type. This high value (trophic position) is the result of the high consumption of prey that occupy different positions in the marine trophic chain of the Ecuadorian Pacific, as is the case of D. gigas (TL = 4.14), S. oualaniensis (TL = 4.09), Ancistrocheirus lesueurii (TL = 4.13), A. rochei eudorax (TL = 4.13) and hemiramphids (TL = 2.82) (Froese and Paul, Reference Froese and Pauly2022). In addition to these food preferences, the large body size of I. oxyrinchus, maximum 4 m TL (Compagno, Reference Compagno2001), its wide range of horizontal (~2500 km) and vertical (~4000 m depth) movements (Gibson et al., Reference Gibson, Streich, Topping and Stunz G2021) and its diet breadth are additional characteristics that allow this species to position itself at the top of the marine food chain (Roff et al., Reference Roff, Doropoulos, Rogers, Bozec, Krueck, Aurellado, Priest, Birrell and Mumby2016) and thus, consume prey from different habitats as well as from various trophic levels.
According to the previous classification and the position estimate of I. oxyrinchus obtained in this study, this species is considered a top predator that fulfils the functions of a carnivore. These results are similar to other species of same family Lamnidae, as the white shark, Carcharodon carcharias, TL = 4.1–5.4 (Kerr et al., Reference Kerr, Andrews, Cailliet, Brown and Coale2006; Hussey et al., Reference Hussey, McCann, Cliff, Dudley, Wintner, Fisk and Domeier2012) and longfin mako shark, Isurus paucus, TL = 3.5–5.7 (Estupiñán-Montaño and Delgado-Huertas, Reference Estupiñán-Montaño and Delgado-Huertas2022). Therefore, this species plays an important role in the health maintenance of the Ecuadorian Pacific marine ecosystem, and may help to regulate lower trophic levels through top-down control (Navia et al., Reference Navia, Cortés and Mejía-Falla2010; Bornatowski et al., Reference Bornatowski, Navia, Braga, Albilhoa and Corrêa2014).
Data
The authors confirm that the data supporting the findings of this study are available within the article.
Acknowledgements
The authors thank the Escuela de Biología of the Facultad de Ciencias Naturales of the Universidad de Guayaquil for allowing us to use its laboratories for the analysis of the samples. C. E. M. thanks the Fundación Alium Pacific. F. G.-M. thanks the Instituto Politécnico Nacional for the fellowships granted (COFAA and EDI).
Author contributions
All authors contributed to conceptualization, formal analysis, investigation, methodology, visualization, writing – original draft, writing – review and editing
Financial support
This research received no specific grant from any funding agency, commercial or not-for- profit sectors.
Competing interests
None.







