Introduction
The Gobiidae is the largest fish family with about 1915 valid species (Fricke et al., Reference Fricke, Eschmeyer and Van der Laan2022) and also the most speciose fish group in the Mediterranean Sea with 77 species (Kovačić, Reference Kovačić2020; Kovačić et al., Reference Kovačić, Renoult, Pillon, Svensen, Bogorodsky, Engin and Louisy2022; Özden et al., Reference Özden, Seyhan Öztürk, Irmak, Oruç and Engin2022). The members of sand goby group (McKay and Miller, Reference McKay and Miller1997) (Knipowitschia, Orsinigobius, Economidichthys, Pomatoschistus, Ninnigobius) are characterized by their small size and mostly euryhaline or freshwater occurrence, while their close relatives ‘expanded sand goby group DBSL’ (McKay and Miller, Reference McKay and Miller1997; Agorreta et al., Reference Agorreta, San Mauro, Schliewen, Van Tassell, Kovačić, Zardoya and Ruber2013) (Deltentosteus, Buenia, Speleogobius, Lebetus) are known as strictly marine taxa and may occur at greater depths (Kovačić et al., Reference Kovačić, Ordines and Schliewen2017, Reference Kovačić, Ordines and Schliewen2018). These genera also contain new species described in the past two decades, and the biodiversity of the DBSL sand goby group has increased to ten valid species (Kovačić, Reference Kovačić2020).
The distribution of the recently described Buenia species, Buenia massutii Kovačić, Ordines and Schliewen, Reference Kovačić, Ordines and Schliewen2017, was restricted to Atlantic Ocean and western Mediterranean and it was reported only from Balearic Islands (type locality) (Kovačić et al., Reference Kovačić, Ordines and Schliewen2017), various locations in the Alboran Sea (Ordines et al., Reference Ordines, Kovačić, Vivas, García-Ruiz and Guijarro2019) and Gulf of Cádiz/north-eastern Atlantic Ocean (Ordines et al., Reference Ordines, Ramírez-Amaro, Burgos, Baro, Kovačić and Sobrino2020). The species was also recorded for the Adriatic Sea (Dulčić et al., Reference Dulčić, Lepen Pleić, Zorica, Bušelić, Šestanović and Kovačić2022). The other new Buenia species, Buenia lombartei Kovačić, Ordines and Schliewen, Reference Kovačić, Ordines and Schliewen2018, is only known from two individuals collected from the continental slope of its type locality, Balearic Islands/western Mediterranean (Kovačić et al., Reference Kovačić, Ordines and Schliewen2018).
The other recently described sand goby species, Pomatoschistus nanus Engin and Seyhan Öztürk, Reference Engin and Seyhan Öztürk2017, was described from the northern coast of the Levantine Sea/eastern Mediterranean and it is the Mediterranean's smallest fish. The fish differs from its congeners by the smallest adult size, deeper bathymetric range (29–41 m) and distinctive morphological characters (Engin and Seyhan Öztürk, Reference Engin and Seyhan Öztürk2017). P. nanus is still known only from its type locality (Antalya/northern Levantine Sea).
The aim of the present paper was to report using morphological and molecular evidence: (1) the easternmost record of B. massutii from two different localities in the eastern Mediterranean Sea, and (2) the second record of P. nanus from the two different locations in the Aegean Sea, known only from type locality since its description.
Materials and method
Sampling
Underwater observations and samplings were performed by two divers using both SCUBA and an electronic closed-circuit rebreather during the different underwater surveys in 2019–2022. Three B. massutii specimens were collected from Erdek/Akçaada/Sea of Marmara at 30 m depth and a specimen was collected from Taksi Island (Gulf of Çandarlı)/Aegean Sea at 65 m depth. The Erdek specimen of B. massutii was released and photographed at a depth of ~3 m while under anaesthesia. Additionally, P. nanus specimens were sampled and photographed from Datça/south-eastern Aegean Sea (three specimens) and İbrice (Gulf of Saros)/north-eastern Aegean Sea (one specimen) at 15–35 m, respectively. Specimens were euthanized by anaesthetic overdose, and then fixed in 4% formalin for morphological identification after having pectoral-fin clips fixed in 96% ethanol for genetic analysis. All specimens were deposited in the Fish Collection Centre of Izmir Katip Celebi University (P. nanus: IKC.PIS.1263–1266; B. massutii: IKC.PIS.1267–1270). Sampling locations and previous records of the species are shown in Figure 1.

Figure 1. Distribution records for Buenia massutii and Pomatoschistus nanus. Empty squares and circle indicate the previous records of B. massutii and P. nanus, respectively. Full squares and circles indicate new occurrences of B. massutii and P. nanus, respectively.
Morphological analysis
Morphological analysis follows Engin and Seyhan Öztürk (Reference Engin and Seyhan Öztürk2017) and Schliewen and Kovačić (Reference Schliewen and Kovačić2008), with the following meristic characters and abbreviations: A, anal fin; C, caudal fin; D1, first dorsal fin; D2, second dorsal fin; P, pectoral fin; V, pelvic disc; LL, scales in lateral series; TR, scales in transverse series. Meristic methods: TR is counted from the anterior origin of the anal fin obliquely upwards and rearwards to the base of D2; LL is counted from pectoral–fin axil along lateral midline, including the scales on the C itself; in D2 and A the last bifid ray is counted as a single ray. Terminology of lateral-line system follows Miller (Reference Miller, Whitehead, Bauchot, Hureau, Nielsen and Tortonese1986) based on Sanzo (Reference Sanzo1911).
Molecular analysis
Total genomic DNA was isolated from the fin clips according to the manufacturer's instructions with a DNA isolation kit (AMBRD Laboratories, Istanbul, Turkey). FishF1/FishR1 primers described in Ward et al. (Reference Ward, Zemlak, Innes, Last and Hebert2005) were used to amplify the DNA barcode region, cytochrome c oxidase I (COI). The thermocycler profile consisted of an initial step of 1 min at 94°C, five cycles of 94°C for 30 s, 50°C for 40 s and 72°C for 1 min, followed by 35 cycles of 94°C for 30 s, 54°C for 40 s and 72°C for 1 min, with a final extension at 72°C for 10 min. Polymerase chain reaction products were checked by 2% agarose gel. Sequencing was carried out at Macrogen Inc., Seoul, South Korea.
Nucleotide sequences were aligned using ClustalW (Thompson et al., Reference Thompson, Higgins and Gibson1994) implemented in MEGA version X (Kumar et al., Reference Kumar, Stecher, Li, Knyaz and Tamura2018) and edited with BioEdit (Hall, Reference Hall1999). Reference sequences belonging to sand gobies and DBSL group were downloaded from GenBank which were mainly provided by Knebelsberger and Thiel (Reference Knebelsberger and Thiel2014), Kovačić et al. (Reference Kovačić, Ordines and Schliewen2018) and Seyhan Öztürk and Engin (Reference Seyhan Öztürk and Engin2019) to investigate genetic relationships. Aligned sequences were compared with existing data and submitted to GenBank with accession numbers: OP580161–OP580167. Maximum-likelihood (ML) and neighbour joining (NJ) analyses were performed with the MEGA version X using the K2P + Gamma + I model due to the lowest Bayesian information criterion values. Uncorrected P and Kimura two-parameter (K2P) distances were used to compare the barcode data. A bootstrap test with 1000 replicates was performed to verify the robustness of the tree.
Results
Morphology
Pomatoschistus nanus Engin and Seyhan Öztürk, Reference Engin and Seyhan Öztürk2017
Material: IKC.PIS.1263–1265; two females and male, 13.82, 13.94 and 14.48 mm SL, from Datça, south-eastern Aegean Sea, 36°40′58″N, 27°34′30″E, 15 m depth. IKC.PIS.1266; female, 13.39 mm SL, from İbrice, north-eastern Aegean Sea, 40°36′02″N, 26°32′404″E, 35 m depth.
Identification: Specimens were identified as members of the Pomatoschistus genus according to the following features: suborbital papilla of longitudinal row a around the lower edge of the orbit; transverse rows c and cp present; anterior and posterior oculoscapular and preopercular canals present; single interorbital canal with a single anterior interorbital pore λ. The studied species IKC.PIS.1263–1266 were identified as P. nanus by following morphological characters: suborbital papilla of longitudinal row a anteriorly passing the pupil and with one transverse row atp. Six transverse suborbital rows. Predorsal area, first dorsal-fin base and breast naked. Anterior point of suborbital row b not reaching level of posterior point of the suborbital row d. Anterior and posterior oculoscapular canals and preopercular canals present but ω, β, δ pores missing. Two elongated brown and parallel stripes on the nape and one brown blotch beginning at the posterior edge of the eye that slightly extends to upper border of the preopercle.
Description: Body cylindrical, moderately elongate, with a slightly compressed caudal peduncle. Eyes dorsolateral. Anterior nostril short and tubular; posterior nostril pore-like and near orbit. Fins: D1 VI; D2 I/8; A I/9; P 15–16 (16: 2, 15: 2); D1 spines almost equal but second spine slightly longer. Wide interdorsal space between D1 and D2. P uppermost and lowermost rays not segmented and P uppermost rays within a membrane. Caudal fin slightly emarginated. Body with ctenoid scales, but predorsal area, first dorsal-fin base and breast naked. Scales in lateral series 31–32 (31: 3, 32: 1). Scales in transverse series 7. Head with anterior oculoscapular canal with pores: σ, λ, α, but ω and β pores missing in all specimens. Preopercular canal with pores γ and ɛ, pore δ missing in all specimens. Six transverse suborbital rows of sensory papilla. Except row cp, all transverse suborbital rows below the longitudinal row a. Longitudinal row d discontinuous and not reaching cp. Transparent body with longitudinal whitish stripe along the lateral midline and five elongate black blotches on this stripe in live coloration (Figure 2). Whitish stripe extending between eyes. The colour of preserved specimens with whitish trunk and scattered brown markings on head (Figure 3). Two elongated and parallel brown stripes on the nape. Pectoral, pelvic and caudal fins transparent. Body with ctenoid scale but predorsal area, opercle and cheek naked. Breast scaled with cycloid scales.

Figure 2. Live coloration of P. nanus, Datça/south-eastern Aegean Sea.

Figure 3. Preserved specimen of P. nanus. IKC.PIS.1265, 14.48 mm SL, male, Datça/south-eastern Aegean Sea.
Buenia massutii Kovačić, Ordines and Schliewen, Reference Kovačić, Ordines and Schliewen2017
Material: IKC.PIS.1267–1269; male and two females, 21.03, 22.62 and 18.28 mm SL, from Akçaada, Erdek, Sea of Marmara, 40°31′23″N, 27°42′24″E, 30 m depth. IKC.PIS.1270; male, 19.96 mm SL, from Taksi Island (Gulf of Çandarlı), eastern Aegean Sea, 38°51′46″N, 26°52′50″E, 65 m depth.
Identification: Specimens were identified as members of the Buenia genus according to the following features: suborbital row of sensory papillae a present; pelvic fins joined in V; V with anterior transverse membrane; anterior oculoscapular and preopercular head canals present; suborbital transverse rows of sensory papillae absent. The studied species IKC.PIS.1267–1270 were identified as B. massutii by the following morphological characters: posterior oculoscapular canal absent. Suborbital row c of 5 papillae. LL 26–28. P 16. The second spine of D1, as the longest spine, reaching to or behind posterior end of D2 in males when folded down, in females not reaching D2. Eye diameter 31.09–33.89% of head length. Caudal fin length 22.37–22.96% of SL.
Description: Body moderately elongated, laterally compressed towards the caudal fin. Head quite long, 31.16–31.67% of SL, snout short. Eyes large, dorsolateral, 31.09–33.89% of head length. Anterior nostril short and tubular; posterior nostril pore-like, mouth oblique. Fins. D1 VI (VI: 6); D2 I/7; A I/6–7 (6: 3, 7: 1); P 16; V I/5 + 5/I. Second spine of the D1 is the longest one and reaching to or behind posterior end of D2 in males when folded down. C rounded. Head with anterior oculoscapular canal semi-closed with pores σ, λ, κ, ω, α, ρ and additional pores or open furrows from interorbital part to pore ρ. Pores σ, λ, κ are present with defined pores. Posterior oculoscapular canal absent. Preopercular canals with pores γ, δ, ɛ and several additional pores. Suborbital longitudinal row of sensory papillae a present with two papillae below the posterior edge of pupil and rows a, c and cp without transverse proliferation. Longitudinal row b short with four papillae anteriorly beginning below rear border of eye. Live coloration: head and body yellowish to brownish with scattered yellowish, orange and dark brown pigments (Figure 4). Predorsal area light brownish with scattered orange chromatophores. A blotch with dark brown dots extending downwards from eye to behind posterior end of mouth, opercle and preopercle. Rare dark brown dots and distinctive orange mark in the upper posterior end of opercle. Underside of lower jaw and prepelvic area whitish. Eyes green golden to brass, with dark green-blue to black pupil. Melanophores on body concentrated mostly along lateral midline forming four dark brown vertical midline stripes extending from shortly above midline downwards across lower lateral side and fifth midlateral horizontal mark on the edge of caudal peduncle and caudal fin. Orange-brown longitudinal lateral stripe passing shortly above the lateral midline through upper ends of dark brown vertical stripes. Ventral side more whitish and not heavily pigmented like upper part of the body. Lower lateral half, in addition to dark brown vertical midline stripes, also with orange dots and bluish silver iridophores. Orange pigmentation with scattered black dots visible on D1 base, mostly anteriorly. Rarely scattered orange pigments on D2, C, A and P. Membrane between C rays origin pigmented dark. The colour of preserved specimens light brown to whitish (Figure 5). A distinctive dark stripe under the eye reaching to rear edge of mouth. Four visible blotches on the lateral midline and the caudal mark.

Figure 4. Live coloration of B. massutii, Akçaada/Erdek/Sea of Marmara.

Figure 5. Preserved specimen of B. massutii. IKC.PIS.1270, 19.96 mm SL, male, Taksi Island (Gulf of Çandarlı)/eastern Aegean Sea.
Molecular analysis
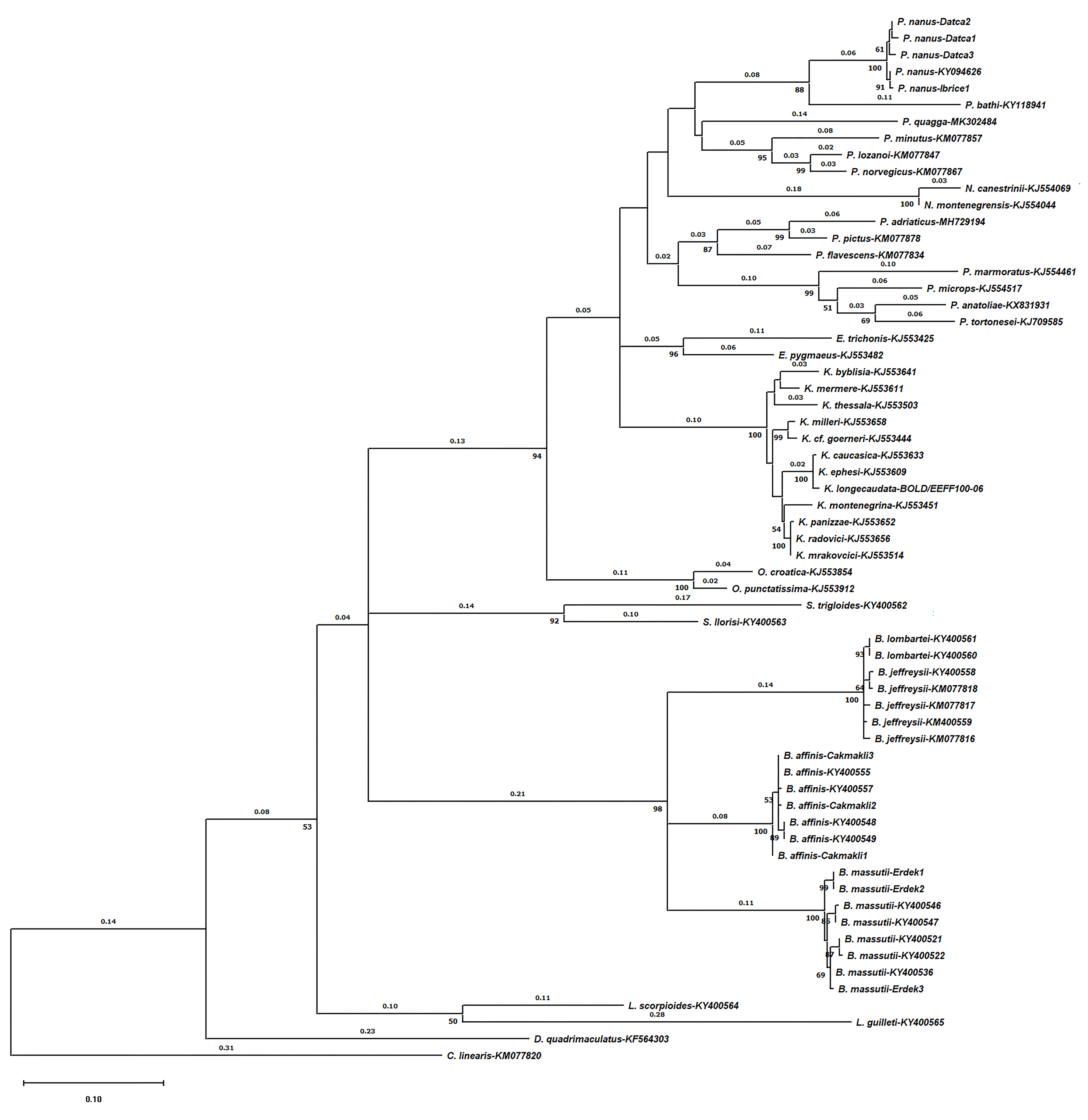
A total of 521 bp of COI fragments of sand gobies and their close relatives DBSL group were analysed. COI clades for sand gobies and DBSL genera were well supported in the ML and NJ phylogenetic analysis (Figure 6 and Supplementary material).

Figure 6. NJ tree based on sand gobies and expanded sand goby group ‘DBSL’ species. The tree is drawn to scale, with branch lengths (above the branches) measured in the number of substitutions per site. Numbers next to branches indicate bootstrap values >50 for 1000 replicates. Taxa in the red rectangular indicate newly sequenced material in this study.
In fact, the lowest interspecific distance between species was observed between Buenia jeffreysii and B. lombartei with 0.8% uncorrected P-distance and K2P-distances (SE: 0.003) which is similar to those reported in Kovačić et al. (Reference Kovačić, Ordines and Schliewen2018). The subsequent lowest genetic distance was observed between B. massutii and Buenia affinis with 11.4% (SE: 0.014) and 13.8% (SE: 0.020) uncorrected P-distance and K2P-distances, respectively. The highest genetic distance was observed between Crystallogobius linearis and B. massutii with 22.5% (SE: 0.019) and 33.7% (SE: 0.040) uncorrected P-distance and K2P-distances, respectively. The molecular analysis also revealed that, except for the identical B. lombartei sequences the low intraspecific distances were observed between B. affinis specimens with 0.4% (SE: 0.002) followed by P. nanus 0.7% (SE: 0.003), B. jeffreysii 0.8% (SE: 0.003) and B. massutii 1.1% (SE: 0.003) for both uncorrected P-distance and K2P-distances those contrasted with large interspecific distance. All these values confirm our morphological identification of B. massutii and P. nanus specimens.
Discussion
The molecular and morphological studies conducted in this study confirmed the presence of B. massutii in the eastern Mediterranean basin and the presence of P. nanus in the eastern Aegean Sea. These records also are the easternmost record including Sea of Marmara for B. massutii and northernmost record for P. nanus.
B. massutii specimens were captured at 30 m depth on a coarse substrate containing biogenic calcareous fragments and shell gravel with coralline deposits. This habitat type is different from the sandy bottom with red algae beds reported by Kovačić et al. (Reference Kovačić, Ordines and Schliewen2017), but it is in concordance with those reported by Ordines et al. (Reference Ordines, Kovačić, Vivas, García-Ruiz and Guijarro2019). Besides, present bathymetric range is shallower than the depth range of B. massutii reported for western Mediterranean and north-eastern Atlantic populations (57–125 m).
The Sea of Marmara, which is under the influence of the Mediterranean and Black seas, is a unique ecosystem with various physico-chemical environments and habitats. The low salinity waters originating from the Black Sea are located in the upper water layer of 15–20 m, and the high salinity water layer originating from the Mediterranean entering the Dardanelles Strait at the bottom (Beşiktepe et al., Reference Beşiktepe, Sur, Özsoy, Latif, Oğuz and Ünlüata1994). The cold and oxygen-rich Mediterranean waters entering the Sea of Marmara from the Aegean Sea sink to their own density levels here (Beşiktepe et al., Reference Beşiktepe, Özsoy and Ünlüata1993). While the nutrient-rich Black Sea waters, which form the surface waters, increase fisheries production, the high salinity waters of the Mediterranean Sea in the lower layer contribute to the increase of biological diversity. These two different water bodies are separated by a strong thermocline, pycnocline and halocline layer at ~20–25 m (Beşiktepe et al., Reference Beşiktepe, Özsoy and Ünlüata1993). The stratified Black Sea water on the surface is rich in nutrients and chlorophyll-a, which significantly limits the penetration of light under 20 m. Accordingly, it exhibits the cold and high salinity mesophotic conditions of the Mediterranean Sea below 20 m (Yılmaz et al., Reference Yılmaz, Baştürk, Tuğrul, Saydam and Ünlüata1990). These complex hydrographic features also allow the presence of the deeper Mediterranean biodiversity at more shallow waters in Sea of Marmara.
The present record of B. massutii represents the first confirmed alive coloration of this species (Figure 4). The photo matches in the coloration pattern and in the body shape of Buenia cf. massutii specimen in Figure 49 in Kovačić et al. (Reference Kovačić, Renoult, Pillon, Svensen, Bogorodsky, Engin and Louisy2022). The present more intensive coloration (Figure 4) can be attributed to shallower depth of photographing with more lights and more contrasting surroundings, as well as to the specimen stress caused by capturing and anaesthetization.
The bathymetric range of P. nanus specimens which were sampled from Datça/south-eastern Aegean Sea station (15 m) also was shallower than the depths reported by Engin and Seyhan Öztürk (Reference Engin and Seyhan Öztürk2017) in its type locality (29–41 m). The description of P. nanus was based on adult and breeding specimens which have 14.9–16.9 mm SL (Engin and Seyhan Öztürk, Reference Engin and Seyhan Öztürk2017). However, specimens which were sampled from Datça were not adults and had 13.9–14.4 mm SL, which might indicate that younger specimens of P. nanus have shallower bathymetric range. This bathymetric range also overlaps with the end of bathymetric distribution of Pomatoschistus bathi which prefers the same substrate with P. nanus.
Conclusion
The findings in this study reveal not only the new geographic distribution data for P. nanus and B. massutii, but also describe for the first time alive coloration of B. massutii and report new ecological data for both recently described species, expanding the known habitat tolerance ranges of these species.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0025315423000292
Acknowledgements
We are very thankful to our advisor Dr Semih Engin for his precious comments. We acknowledge the Izmir Katip Çelebi University Scientific Research Project Coordination Unit for funding the presently reported study in the frames of the grant no. 2022-GAP-SUÜF-0018 and TÜBİTAK grant no. 121 Y 553.
Author's contribution
Dilruba Seyhan Öztürk: conceptualization, investigation, methodology and writing original draft. Adnan Çağlar Oruç: Investigation, methodology and figure editing.
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.