


Introduction
Most bird species inhabiting Mediterranean steppe-like habitats are of conservation concern (BirdLife International, 2004). For most of these species population sizes and recent declines are well studied, and declines are known to be due to farming-related pressures and changes in land use (Knick & Rotenberry, Reference Knick and Rotenberry1995; Le Cuziat et al., Reference Le Cuziat, Lacroix, Roche, Vidal, Médali, Orhant and Béranger2005). However, despite being of conservation concern in Europe, most of these species are categorized globally as Least Concern (IUCN, 2007), with important populations believed to be present in the steppe-like habitat of North Africa (Cramp, Reference Cramp1988). However, the precise ranges, population sizes and, most importantly, population trends of these North African bird populations are practically unknown. Given the accelerated pace of development in these countries (Green et al., Reference Green, El Hamzaoui, Aziz El Agbani and Franchimont2002), it likely that these bird species are experiencing pressures similar to that which they experience in Europe. Knowledge of population trends of many of these steppe species in North Africa is thus required to establish their conservation status at the global scale.
Dupont's lark Chersophilus duponti is a typical example. The species is considered one of the least abundant passerine species in Europe (Garza et al., Reference Garza, Suárez, Tella, Madroño, González and Atienza2004). Its continental distribution is limited to the shrub steppes of the Mediterranean region of Spain, and its breeding population is 1,300–1,900 territories (Garza et al., Reference Garza, Traba and Suárez2003, Reference Garza, Suárez, Herranz, Traba, García de la Morena and Morales2005; Tella et al., Reference Tella, Vögeli, Serrano and Carrete2005). Dupont's lark is categorized as Endangered in Spain because of recent habitat loss driven by agricultural expansion, reforestation schemes and other land use changes (Garza et al., Reference Garza, Suárez, Tella, Madroño, González and Atienza2004).
Beyond Europe, the species’ population size and distribution are poorly known, partly because of its secretive habits. It is considered to be sparsely distributed and uncommon in most areas of its relatively small and fragmented range, restricted in North Africa to the north of the Maghreb (Morocco, Algeria and Tunisia), Libya and Egypt (Cramp, Reference Cramp1988; De Juana & Suárez, Reference De Juana, Suárez, Del Hoyo, Elliott and Christie2004). Within the African range, the nominal subspecies C. d. duponti is restricted to Morocco, northern Algeria and Tunisia, and is considered an evolutionary unit distinct from the European C. d. duponti (García et al., Reference García, Suárez, Garza, Calero-Riestra, Hernández and Pérez-Tris2008), whereas the subspecies C. d. margaritae occurs throughout the rest of the African species' range. Detailed distribution, population size and trends in these countries are unknown. Only Blondel (Reference Blondel1962) for Algeria and Suárez et al. (Reference Suárez, Fernández and De Lope1986) for Morocco have reported on the species’ abundance, all other available information being qualitative (see Thévenot et al., Reference Thévenot, Vernon and Bergier2003 for Morocco; Isenmann & Moali, Reference Isenmann and Moali2000 for Algeria; Isenmann et al., Reference Isenmann, Gaultier, El Hili, Azafzaf, Dlensi and Smart2005 for Tunisia). Despite the shortage of data, Dupont's lark was categorized globally as Near Threatened in 2005 (IUCN, 2007), reflecting the concern that its main North African populations may not have remained stable in recent decades (e.g. Isenmann et al., Reference Isenmann, Gaultier, El Hili, Azafzaf, Dlensi and Smart2005 for Tunisia).
The purposes of this study were to (1) delimit the current distribution of Dupont's lark in Morocco, one of the largest breeding populations in its North African range (De Juana & Suárez, Reference De Juana, Suárez, Del Hoyo, Elliott and Christie2004), (2) characterize its preferred habitat and estimate population density and size in Morocco, and (3) discuss the species' conservation status at the global level.
Methods
Geographical range, habitat preferences and occurrence
The whole distribution range of the species in Morocco (according to Thévenot et al., Reference Thévenot, Vernon and Bergier2003) was searched (Fig. 1), totalling c. 90,000 km2 in eastern Morocco (from latitude 34°23′ to 32°11′ N and from longitude 5°02′ to 1°50′ W) and the central Anti-Atlas (31°24′ to 30°22′ N and 7°46′ to 5°45′ W), where there have been some recent records (Scheepen & Meijer, Reference Scheepen and Meijer1996; Bergier, Reference Bergier2004). In eastern Morocco rangeland livestock is widespread along the flat high plateaux (rising to > 1,000 m), sparsely covered by steppe vegetation dominated by halfa grass Stipa tennacissima. In this area (which includes Aïn-Benimathar, Rekkam and Midelt-Missour) the range extends west to the High Moulouya (with a similar landscape, although some crops are cultivated along the Moulouya River) and south to the Saharan Atlas (north of Jbel Maïz; Heim de Balsac, Reference Heim de Balsac1928). In the south-west of the species' range (the central Anti-Atlas) steppe vegetation is dominant (mostly Artemisia spp. shrub), with interspersed crops in the valley bottoms (Thévenot et al., Reference Thévenot, Vernon and Bergier2003). Climate is arid with an average rainfall of 210 mm per year, and is higher in the north (300 mm) and lower in the south (120 mm).
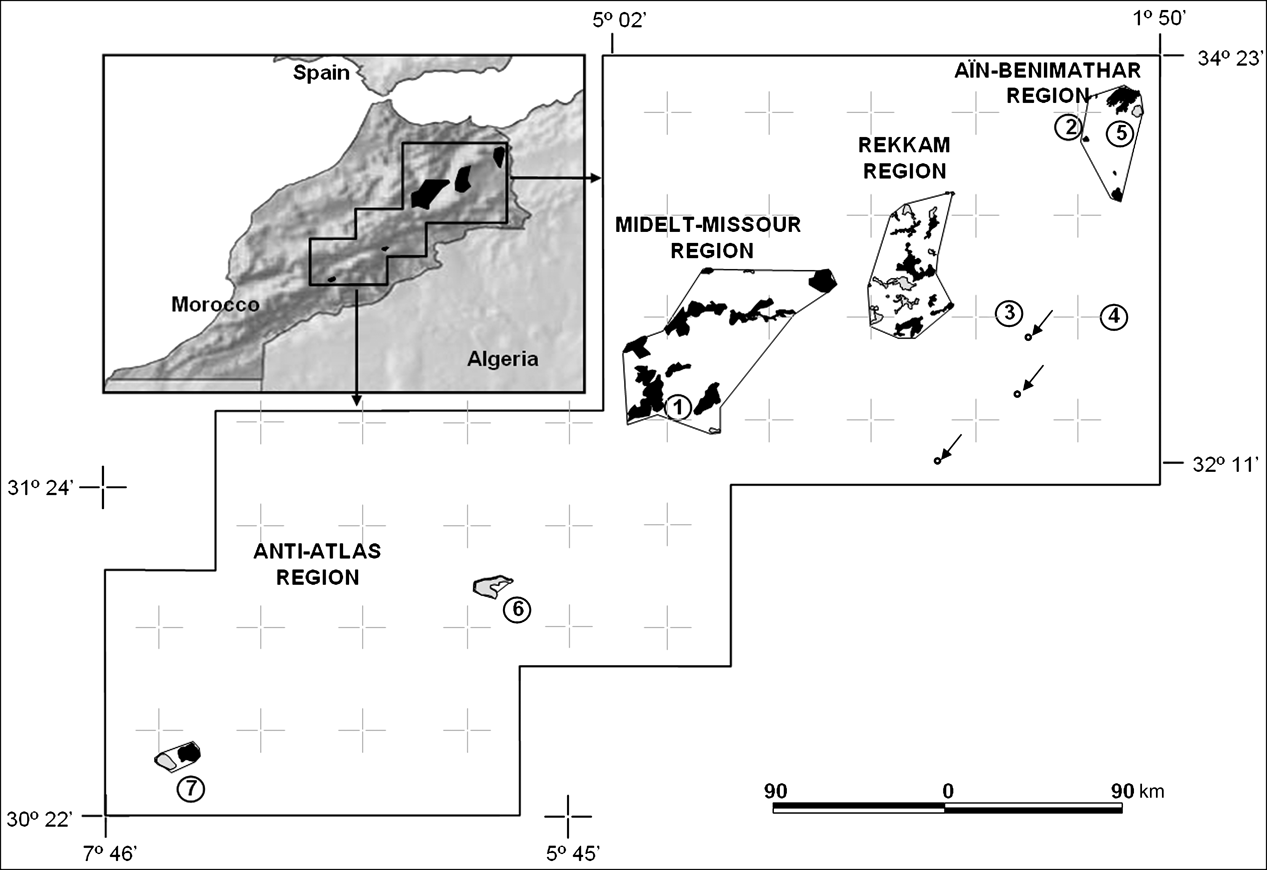
Fig. 1 Distribution of Dupont's lark in Morocco, showing the four main areas in which the species occurs, and the villages and areas mentioned in the text (1, Midelt; 2, Merija; 3, Matarka; 4, Tendrara; 5, Aïn-Benimathar; 6, Boumalne Dadès; 7, Tiz-n-Taghatine). Polygons show the species’ extent of occurrence in each region (see Methods), containing areas of occupancy (black areas) and areas of suitable habitat where the species was not recorded (grey areas). Small circles with arrows indicate the location of recent records (2003–2005) of the species during the breeding period (O. Fontaine & A. Qninba, pers. comm.). Inset shows the location of the main map in Morocco.
We visited all areas described as having suitable habitat for the species (Table 1) according to the described habitat preferences of the species in Morocco. We searched in dense halfa grass (> 15% cover), sparse halfa grass (< 15% cover), and Artemisia spp. shrub on hard or pebbly soils, avoiding sand (Heim de Balsac & Mayaud, Reference Heim de Balsac and Mayaud1962; Thévenot et al., Reference Thévenot, Vernon and Bergier2003) but we also visited other vegetation types that offer the minimum vegetation cover and height needed by the species (according to available information for the Spanish population; Garza & Suárez, Reference Garza and Suárez1990). Thus, we also searched in vegetation areas of Noaea mucronata (mixed sometimes with Anabasis articulata and Artrocnemum spp.), and species-mixed xerophytic vegetation of Hammada scoparia (sometimes with Lycium intrincatum and halfa grass), and including some areas with atypical vegetation such as Fredolia aretioides, or grassy-sandy areas. Only areas of flat or rolling terrain were searched because the species is absent from gradients steeper than 15%, at least in Spain (Seoane et al., Reference Seoane, Justribó, García, Retamar, Rabadán and Atienza2006).
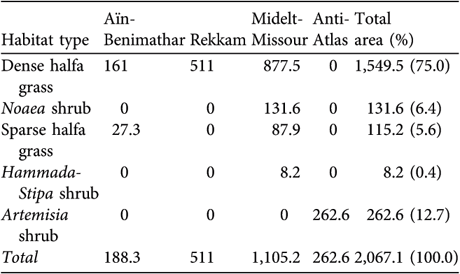
Table 1 Areas (km2) of the five habitat types in which we found Dupont's lark in each of the four regions where the species is known or believed to occur, and overall (Fig. 1; see text for further details of habitat types and regions).
We made six field visits between 2004 and 2006: December 2004 and December 2005 (10 and 15 days, respectively, by six people), March 2006 (12 days by six people), April 2005 (15 days by seven people), August 2005 (12 days by two people), and November 2005 (13 days by two people). In total the survey effort was 377 people days. Presence of the species was identified by visual or auditory detection of individuals, and eliciting territorial calls by broadcasting the males’ territorial songs and calls (Garza et al., Reference Garza, Traba and Suárez2003). The latter took place anywhere in the surveyed patches, depending on accessibility and the feasibility of walking through the patches.
The alarm calls of Dupont's lark can be detected year-round (Laiolo et al., Reference Laiolo, Vögeli, Serrano and Tella2005), which enabled us to detect presence by playback trials at any time. The position of each detected bird (using a global positioning system, GPS, error ± 10 m) and the habitat type (Table 1) in which it was found were recorded.
To estimate the extent of suitable habitat and the area in which the species is potentially distributed, we followed a three-stage method. Starting from the sites identified as containing the species, we used a GPS to geo-reference the boundaries of the habitat types present, up to a maximum distance of 5 km. Boundaries were assessed by driving through, and/or scanning the area with binoculars and telescopes, when possible, from any available vantage points. The boundaries of these units were then confirmed and extended over nearby areas (to a maximum of 30 km) by means of visual similarity analysis on satellite images (three Landsat ETM+ bands, GeoCoverTM, year 2000, pixel size 14.2 × 14.2 m). These homogeneous patches of habitat and their boundaries were rechecked and resampled in the following field visits, both to adjust geo-referencing of estimated boundaries and to check for the presence of the species. Survey coverage was thorough even for potentially less suitable habitat patches such as halfa grass with sparse cover, or species-mixed shrub vegetation.
Extent of occurrence and area of occupancy
All geo-referenced patches and bird locations were incorporated into the geographical information system Arcview v. 3.2 (ESRI, Redlands, USA). Following IUCN (2001) criteria, the extent of occurrence was measured as the area inside a minimum convex polygon (the smallest polygon in which no internal angle exceeds 180°) containing all the sites of recorded or inferred occurrence (i.e. all delimited patches with suitable habitat), and subtracting discontinuities of unsuitable habitat within the overall distribution of the species (IUCN, 2001). Area of occupancy was measured as the sum of the areas of delimited patches with suitable habitat within which the species was recorded.
Population density and size
The species’ density was estimated by mapping all males during line transect censuses. Although other procedures could provide more fine results at the local scale (Garza et al., Reference Garza, Traba and Suárez2003) we used this method because of the constraints imposed by the large area surveyed and the time available. Because the extent of the different habitats was not homogeneous for a given region, the number of censuses in each habitat type was determined in relation to areal extent. Length of transects were 1,200–4,700 m. Censuses were made only during the breeding season (field visits in March–April) and 30–60 minutes before sunrise, because Dupont's lark is most vocally active just before dawn (Aymí et al., Reference Aymí, Elliot and Martínez1994; De Juana & Suárez, Reference De Juana, Suárez, Del Hoyo, Elliott and Christie2004) and most birds sing when flying (Cramp, Reference Cramp1988), which enabled us to discriminate between individuals and record their exact position. All individuals were geo-referenced by GPS to calculate their perpendicular distance to the transect line, and to avoid duplicated records. For analyses we only took into account individuals detected within 200 m either side of the transect line (see Garza et al., Reference Garza, Traba and Suárez2003, 2005, for a review of the problems associated with distance estimation for this species). We have assumed there are no differences in detectability between habitats because (1) we were unable to find evidence of different detectabilities in relation to habitat type in the literature, even on related species, (2) the habitat structure (vegetation cover and height) is similar in the habitats surveyed (J.T. García & F. Suárez, unpubl. data) in these areas of Morocco, and (3) Dupont's lark's vocal activity during census time is maximal, and most singing birds were detected while flying.
Maximum, minimum and mean population sizes of Dupont's lark in each region were calculated as the product of the average density (and its 95% upper and lower confidence limits) observed in each habitat and the area of each habitat type (Table 1). Calculations were made first for each studied area, and mean, minimum and maximum values for the total population of the species in Morocco were then calculated as the sum of estimated values for the regions surveyed.
Results
Geographical range
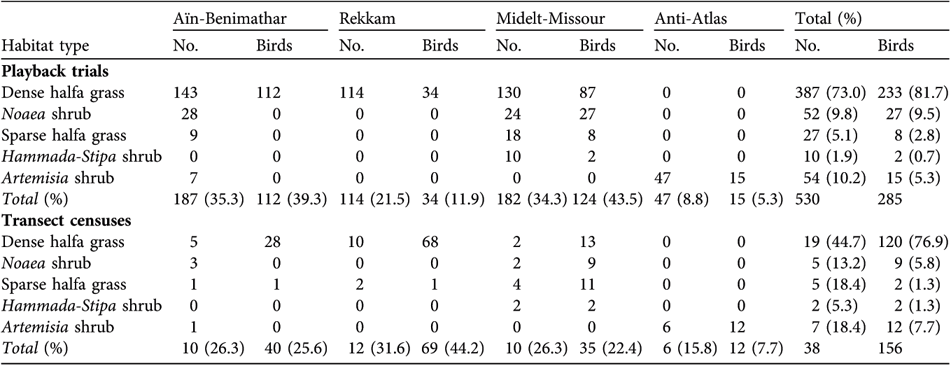
The total number of playback trials was 530, with a total trial time of 207 hours, and a total number of geo-referenced Dupont's larks of 285. The distribution of trials among regions and habitat types is summarized in Table 2. The Moroccan distribution of Dupont's lark covers a wide geographical range (Fig. 1). In eastern Morocco we found the species in three regions: the high plateaux, from 30 km north to 45 km south of Aïn-Benimathar (although breeding individuals were also detected west of Aïn-Benimathar, near Merija); the Rekkam Plateau, north to Debdou and south to Matarka; the High Molouya, between Midelt and Missour south-east and Zeida and Enjil north-west. In the central Anti-Atlas region the species was present near Tiz-n-Taghatine (15 Dupont's lark detected in 24 trials), but not in Boumalne Dadès (n = 23 trials). Because we searched this latter region less intensively it is possible that some remnant populations may still occur there, principally close to Boumalne Dadès, where the species has been detected previously (Thévenot et al., Reference Thévenot, Vernon and Bergier2003) and suitable habitat for the species exists. We did not detect the species either in the plains south of Tendrara (n = 92 trials) or in the plain of Tafrata (n = 10 trials).
Table 2 Number of playback trials and transect censuses, and Dupont's larks located, in each of the five habitat types in which we found the species in each of the four regions searched, and overall (Fig. 1; see text for further details).
Habitat
The vegetation of sites in which we found Dupont's lark was typical of climax steppe vegetation in Morocco, generally dominated by relatively dense stands of halfa grass (Table 1). Qualitative differences in floristic composition and coverage were nevertheless found between regions (J.T. García & F. Suárez, unpubl. data), with north-eastern sites dominated by dense halfa grass stands (especially in the Rekkam region), whereas non-degraded Artemisia shrub stands (with A. herba-alba and A. campestris) were scarce and almost exclusive to the Anti-Atlas. Dominant sparse halfa grass stands were found in the Aïn-Benimathar and Midelt-Missour regions, the latter presenting some Noaea mucronata shrub and mixed Hammada scoparia-Stipa tennacissima stands as well.
The area of suitable habitat for Dupont's lark in Morocco was estimated to be 2,067 km2 (Table 1). The main regions containing suitable habitat are Midelt-Missour (53.5%) and Rekkam (24.7%), and to a lesser extent the Anti-Atlas (12.7%) and Aïn-Benimathar regions (9.1%). Total extent of occurrence in Morocco was calculated to be 10,922.9 km2, comprising 37 patches of suitable habitat with an average and median surface of 66.05 km2 and 55.08 km2, respectively (Table 3).
Table 3 Descriptive parameters of the geographical range, and population estimate (in each of the five habitat types in which we found the species, and mean, maximum and minimum) of Dupont's lark in each the four regions in which the species was found (Fig. 1), and overall.
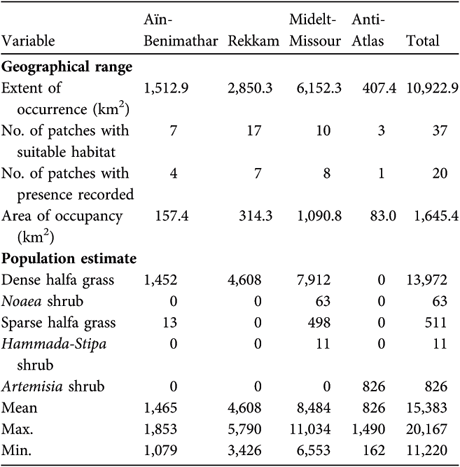
The presence of the species was confirmed in 54% of suitable habitat patches, covering at least 79.6% of the total area of suitable habitat estimated for the whole study area (Table 3). The size of patches differed significantly between occupied and unoccupied patches, being significantly higher in the former (mean 82.27 ± SE 18.37 and 24.80 ± SE 7.83 km2, respectively: general linear model F 1,35 = 7.33, P = 0.010). Area of occupancy was calculated to be 1,645.4 km2 in 20 sub-areas with an average and median surface of 82.27 km2 and 63.44 km2, respectively (Table 3).
Dense stands of halfa grass appear to be the main habitat for the species in Morocco, containing 91% of the total estimated population. Habitat dominated by Artemisia shrub (5.4%) and sparse stands of halfa grass (3.3%) were next in importance, and Noaea shrub (0.4%) and mixed Hammada-Stipa shrub (0.1%) appeared to be less important as habitat for the species.
Population density
After removing unclear records (n = 23), possible duplicate individuals (n = 31), and birds detected at long distances (n = 54), we had 156 geo-referenced individuals from transect censuses. A total of 82.7 km was censused, covering 3,308 ha (1,696 ha in 2005, and 1,312 ha in 2006). Density estimates (including six transects with no records of the species) ranged from 0.00 to 2.40 birds per 10 ha, averaging 0.60 ± SD 0.52 birds per 10 ha. Dupont's lark density varied among habitat types (one way ANOVA F 4,33 = 6.08, P < 0.001), even when excluding censuses without positive responses (F 4,27 = 2.90, P = 0.040), being highest in dense halfa grass (all censuses, 0.902 ± SD 0.514 birds per 10 ha,) and Noaea shrub habitat (0.567 ± SD 0.267 birds per 10 ha), and lowest in sparse halfa grass (0.048 ± 0.067 birds per 10 ha) and Hammada-Stipa shrub habitat (0.137 ± SD 0.087 males per 10 ha). Density in areas dominated by Artemisia shrub was intermediate (0.315 ± SD 0.341 birds per 10 ha).
Population size
Our estimate of the total population size of Dupont's lark in Morocco is 15,400 males (Table 3). Highest estimated populations were in the regions of Midelt-Missour (56.0% of the total population), and Rekkam (29.4%), with Aïn-Benimathar region (9.3%) and the Anti-Atlas (5.3%) having smaller populations (Table 3).
Discussion
To our knowledge this is the first time that the population size of Dupont's lark has been estimated outside its European range (Thévenot et al., Reference Thévenot, Vernon and Bergier2003). Our figure of 15,400 singing males in Morocco must, however, be considered as the minimum population, and interpreted with caution, because of two potential biases. Firstly, the extent of suitable habitat for the species is difficult to evaluate because of the detailed scale that is needed to reflect the relevant parameters appropriately, such as vegetation cover and composition, soil type and slope. Our method, based on successive approximations combining analysis of satellite images and intensive field work, appeared to be reliable, as confirmed by the discovery of individuals in apparently unusual habitats, such as mixed Hammada-Stipa shrub. Nevertheless, reported observations south of Matarka, in the furthest Rekkam region (O. Fontaine, pers. comm.; Fig. 1), in the north of Jbel Maïz (Heim de Balsac, Reference Heim de Balsac1928) and in the north of Jbel Grouz (A. Qninba, pers. comm.; Fig. 1), and near Boumalne Dadès, in the Anti-Atlas (Thévenot et al., Reference Thévenot, Vernon and Bergier2003; Fig. 1), suggest that the species' distribution may extend beyond the areas we delimited in our study. However, densities will probably be low in these marginal sites because habitat (dominated by Lyceum intrincatum and Fredolia aretioides, or with sparse Artemisia spp.) searched there during our summer visit seemed poorly suited to the species. Hence, it is possible that the total Moroccan population could exceed our initial estimate by at most a few hundred males. Secondly, our estimate of density, which is lower than found in Spain (c. 0.7 birds per 10 ha in Morocco, and c. 1.0 birds per 10 ha in Spain; Garza et al., Reference Garza, Suárez, Herranz, Traba, García de la Morena and Morales2005), has a higher variance because we censused over an area of occupancy of near 2,000 km2 of suitable but heterogeneous habitat. For example, the coefficient of variation of estimated densities in Spain is 19.9% (data from Tella et al., Reference Tella, Vögeli, Serrano and Carrete2005) whereas it is 84.3% for Morocco. This is why there is a wide range (11,220–20,167) in our estimate of the number of singing males in Morocco.
Our results also confirmed that halfa grass steppe is the main habitat for Dupont's lark in Morocco, supporting previous information on the species' habitat preferences (Thévenot et al., Reference Thévenot, Vernon and Bergier2003). Differences in the size of occupied and unoccupied patches suggest that the species is sensitive to habitat fragmentation as we did not find it in smaller areas. The distribution of Dupont's lark is probably influenced by human activities. Overgrazing, land reclamation and burning are the main causes of the contraction and degradation of steppe habitat in Morocco (Benabid, Reference Benabid2000), affecting primarily dense halfa grass. Regression of these steppes has occurred since the beginning of the 20th century in Morocco (and also in Algeria and Tunisia), with the area of steppe dominated by halfa grass undergoing a sharp decline (Quézel, Reference Quézel2000). All these factors increase habitat fragmentation but how they affected Dupont's lark in the past, and will do so in the future, is difficult to discern, especially given the shy and cryptic nature of the species.
In the mid 1980s the species was continuously distributed throughout the plains of eastern Morocco bordering Algeria (Suárez et al., Reference Suárez, Fernández and De Lope1986), at which time these areas were characterized by a heterogeneous mosaic of dominant halfa grassland and Noaea shrub under low to medium livestock pressure, interspersed with extensive cereal plots. From our observations this landscape seems to have suffered three major changes, mainly because of human activities: (1) an expansion of intensively managed plantations of fodder shrubs (e.g. Atriplex spp.) to supply livestock, (2) a sharp decline in the total halfa grassland area, with many remaining stands severely affected by drought and by locust Schistocerca gregaria plagues in certain years (Ullman, Reference Ullman2006), and (3) an increase in the cover of sandy soils at the expense of hard and pebbly surfaces (in comparison to observations provided by Suarez et al., Reference Suárez, Fernández and De Lope1986). As a consequence, the area of habitat suitable for Dupont's lark in the high plateaux has diminished markedly in the last 2 decades. The area of halfa grass between Aïn-Benimathar in the north and Tendrara in the south (a heterogeneous area c. 80 × 30 km) has almost disappeared since 1985 and remnants are severely degraded, with a high proportion of dead plants (68.6 ± 35.5% of halfa plants dead, n = 15 sampled halfa grass patches). Decline of Dupont's lark populations in this area is therefore unsurprising. South to Tendrara, where the species had an abundance in 1985 similar to that in Aïn-Benimathar today (2.55 and 2.07 birds per 10 ha, respectively; Suárez et al., Reference Suárez, Fernández and De Lope1986) the species is virtually absent today. It is likely that other species linked to halfa grass, such as the scrub warbler Scotocerca inquieta, which is almost non-existent today (J.T. García & F. Suárez, unpubl. data) but was frequent in the 1980s (Suárez et al., Reference Suárez, Fernández and De Lope1986), have also suffered consequent population declines in eastern Morocco during this period.
However, we have no evidence for similar large losses of habitat in the remaining distribution area of Dupont's lark in Morocco. Although no former records are available for comparative purposes from Midelt-Missour or Rekkam, we have not detected plantations of fodder crops or such high halfa grass mortality in these areas as in Tendrara (34.4 ± SD 9.7%, n = 41, and 11.9 ± SD 16.4%, n = 21, of plants dead from Midelt-Missour and Rekkam, respectively). However, in these areas we found a higher presence of complementary suboptimal habitats, such as Noaea and Hammada-Stipa shrubs, on which the species is able to persist, albeit at lower densities than in halfa grass. It is necessary, however, to develop effective management to prevent habitat degradation and population fragmentation that, in turn, may cause irreversible loss of habitat and significant long-term consequences for the conservation of Dupont's lark, as reported in other steppe-land bird species (Knick & Rotenberry, Reference Knick and Rotenberry1995; Le Cuziat et al., Reference Le Cuziat, Lacroix, Roche, Vidal, Médali, Orhant and Béranger2005).
According to our conservative calculations, the Moroccan population is > 7 times larger than the European population, with c. 15,400 singing males in Morocco compared to 2,000 in Spain (Garza et al., Reference Garza, Traba and Suárez2003, Reference Garza, Suárez, Herranz, Traba, García de la Morena and Morales2005). The Moroccan and Spanish populations together probably amount to an important part of the global population of Dupont's lark, although the extent of the potential breeding range of the species in other North African countries could be even larger than in Morocco. Research is needed in Algeria, where the species' presence is probably related to halfa grass and Artemisia shrub coverage in the high plateaux and the north-western edge of the Sahara (Isenmann & Moali, Reference Isenmann and Moali2000), and in Tunisia where, in spite of a recently reported decline because of overgrazing and agricultural development (Isenmann et al., Reference Isenmann, Gaultier, El Hili, Azafzaf, Dlensi and Smart2005), a significant area of the species' preferred habitats remains. Data are also needed for Libya and Egypt, where scattered and minor populations may remain (De Juana & Suárez, Reference De Juana, Suárez, Del Hoyo, Elliott and Christie2004).
In summary, we found that Dupont's lark has a smaller population and a reduced breeding area in one region of its Moroccan range in comparison to quantitative records from 20 years ago, although this does not seem to be a general pattern in the lark's Moroccan range. More accurate information on the range and population size of the species elsewhere in North Africa is needed. Irrespective of the species’ global conservation status, it is essential to study the medium and long-term dynamics of the species' habitat and the species' reproductive and population ecology across its North African range. We are now extending our field work to Tunisia, where preliminary data has shown that the population of Dupont's lark is low (Authors, unpubl. data), and we are planning future field work to assess the species in Algeria. The population of Dupont's lark in Morocco is still large but it may be only a matter of time before the species experiences critical declines similar to that of the European population. Finally, it is likely that this urgency is not exclusive to Dupont's lark and can be extended to other species of open Mediterranean habitats showing strong declines in Europe (e.g. the black-bellied and pin-tailed sandgrouses Pterocles orientalis and Pterocles alchata, Suárez et al., Reference Suárez, Martínez, Herranz and Yanes1997) that have important, but as yet unquantified, North African populations (De Juana, Reference De Juana, Del Hoyo, Elliott and Sargatal1997).
Acknowledgements
We would like to thank the Moroccan government for fieldwork permits, and M. Radi and M. Znari (Université Cadi Ayyad, Marrakech) for help with arrangements. E. Ramírez, R. del Pozo, A. Ramírez, J. Viñuela and the Colectivo Ornitológico Cigüeña Negra helped us during fieldwork. O. Fontaine and A. Qninba provided records of the south-eastern distribution of the species. J.L. Tella and an anonymous referee commented on and helped improve the manuscript. This study was funded by the Spanish Ministry of Environment (MMA) and the Spanish Agency for International Cooperation. Thanks are due to J.J. Areces and B. Heredia (MMA), who provided enthusiasm and institutional support.
Biographical sketches
Jesús T. García researches the genetics, ecology and conservation of steppe birds, particularly intra- and interspecific competition, and also host-parasite interactions in arid environments. Francisco Suárez has carried out research on the relationships between agriculture, land-use changes and larks, and is coordinating a project on Dupont's lark in Spain. Vicente Garza works on the ecology and conservation of raptors and steppe birds and is also participating in the Dupont's lark project. Jorge H. Justribó works on ecology and conservation of steppe passerines and raptors and environmental impact assessment, and is also taking part in the Dupont's lark project. Juan J. Oñate works on the analysis of the design and impact of agricultural policies on biodiversity components, with a special focus on the EU's Common Agricultural Policy and pseudo-steppe birds. Israel Hervás is currently participating in a research project on the ecology of steppe birds in Spain. María Calero works on the relationship between blood parasites and ecology of steppe birds. Eladio L. García de la Morena's work focuses on ecology and conservation of steppe birds and the environmental impact of human infrastructures.




