



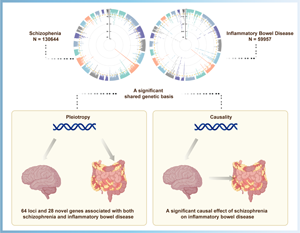
Introduction
Schizophrenia (SCZ) is one of the most severe psychiatric disorders affecting approximately 1% general population globally and is listed as the top disability disease in mental disorders across the global burden of disease study (Ferrari et al., Reference Ferrari, Santomauro, Herrera, Shadid, Ashbaugh, Erskine and Whiteford2022; Jauhar, Johnstone, & McKenna, Reference Jauhar, Johnstone and McKenna2022). A growing body of epidemiological evidence shows the widespread comorbidity among SCZ patients, especially for inflammatory bowel disease (IBD), (e.g. Bernstein et al., discovered the risk of SCZ in IBD patients was 1.64 folds higher than non-IBD patients (Bernstein et al., Reference Bernstein, Hitchon, Walld, Bolton, Sareen and Walker2019), and in Sung's study (Sung et al., Reference Sung, Zhang, Wang, Bai, Tsai, Su and Chen2022), there was a 3.28 times greater risk of IBD among SCZ patients than non-SCZ patients), highlighting a crucial role of the brain–gut axis (BGA) (Gracie, Hamlin, & Ford, Reference Gracie, Hamlin and Ford2019). SCZ patients were found to have a disruption of the intestinal epithelial barrier and reflection of inflammatory cytokines (Gao et al., Reference Gao, Fan, Yang, Ma, Zhao, He and Zhu2022; Murphy et al., Reference Murphy, Walker, O'Donnell, Galletly, Lloyd, Liu and Weickert2022; Szabo et al., Reference Szabo, O'Connell, Ueland, Sheikh, Agartz, Andreou and Djurovic2022). Therefore, clinicians have to face a critical issue of how to safely and effectively treat individuals with both SCZ and IBD.
A recent study has reported strong positive genetic correlations between SCZ and ulcerative colitis (UC) as well as Crohn's disease (CD), indicating that the underlying biological mechanism of the BGA is partially attributed to shared genetic architecture (Tylee et al., Reference Tylee, Lee, Wendt, Pathak, Levey, De Angelis and Polimanti2022). Both SCZ and IBD are highly heritable and heterogeneous diseases, with twin studies estimating family heritability of 79% for SCZ, and 67–75% for IBD (Gordon, Trier Moller, Andersen, & Harbord, Reference Gordon, Trier Moller, Andersen and Harbord2015; Hilker et al., Reference Hilker, Helenius, Fagerlund, Skytthe, Christensen, Werge and Glenthøj2018). Genome-wide association studies (GWASs) of SCZ and IBD have reported several susceptibility loci related to immune-related signals (Jostins et al., Reference Jostins, Ripke, Weersma, Duerr, McGovern, Hui and Cho2012; Liu et al., Reference Liu, van Sommeren, Huang, Ng, Alberts, Takahashi and Weersma2015; Trubetskoy et al., Reference Trubetskoy, Pardiñas, Qi, Panagiotaropoulou, Awasthi and Bigdeli2022). Moreover, through a cross-tissue transcriptome-wide association study, Uellendahl-Werth et al. have identified several shared susceptibility genes between SCZ and IBD, including NR5A2, SATB2, and PPP3CA (Uellendahl-Werth et al., Reference Uellendahl-Werth, Maj, Borisov, Juzenas, Wacker, Jørgensen and Ellinghaus2022). These findings reflect genetic overlap existed between SCZ and IBD, but the magnitude of shared genetic components and the causality relationship between them remains unclear.
By leveraging the hitherto largest GWAS summary statistics of SCZ and IBD, we conducted a comprehensive genetic pleiotropic analysis to uncover shared loci, genes, or biological processes underpinning the observed co-morbidity between SCZ and IBD. First, we performed Linkage disequilibrium score regression (LDSC) to assess the genetic correlation between these two traits. Second, we conducted a cross-trait meta-analysis by multi-trait analysis of GWAS (MTAG) to uncover risk loci shared between the two traits. After that, a summary-data-based Mendelian randomization (SMR) analysis was done to discover novel pleiotropic genes between SCZ and IBD. Then, co-expressed and pathway enrichment analyses were performed to obtain biological insights for those novel pleiotropic genes. Finally, Mendelian randomization analyses were conducted to examine the causal correlation between SCZ and IBD, UC, or CD. This study aims to provide novel insights into the biological mechanisms and therapeutic targets underlying these two disorders.
Methods
Study design and GWAS datasets
The overall study design is depicted in Fig. 1. The GWAS summary statistics of SCZ were derived from European ancestry with 53 386 cases and 77 258 controls (Trubetskoy et al., Reference Trubetskoy, Pardiñas, Qi, Panagiotaropoulou, Awasthi and Bigdeli2022). Individuals with schizophrenia or schizoaffective disorder were included as cases. For IBD, including UC and CD, the GWAS summary datasets were extracted from a genome-wide association study of predominantly European ancestry, which included 25 042 cases and 34 915 controls for IBD, 12 366 cases, and 33 609 controls for UC and 12 194 cases and 28 072 controls for CD, respectively (de Lange et al., Reference de Lange, Moutsianas, Lee, Lamb, Luo, Kennedy and Barrett2017). Cases were diagnosed using endoscopic, histopathologic, and radiologic criteria. The summary-level data used in this study were publicly available, and all GWAS received ethical approval from the relevant institutional review boards, informed consent from participants, and strict quality control, the sources of these data are provided in online Supplementary Table S1. Ethical approval was not required for this study as it was accessed from summary-level data.

Figure 1. Overall study design.
Abbreviations: SCZ, schizophrenia; IBD, inflammatory bowel disease; UC, ulcerative colitis; CD, Crohn's disease; MR, Mendelian randomization; KEGG, Kyoto Encyclopedia of Genes and Genomes.
Statistical analysis
Genetic correlations analysis
We used the Linkage disequilibrium score regression to estimate the heritability of SCZ and each of IBD, UC, and CD. Heritability, taking the value 0–1, is the magnitude of the influence of inherited components on a trait and is defined as the ratio of the genetic variance to the variance of the trait. Generally, greater than 30% is considered as high heritability, 10–30% as moderate heritability, and less than 10% as low heritability. The LD score of a SNP can be defined as the sum of the SNP and other SNPs in a certain area, which measures the magnitude of genetic variability labeled by a SNP. Before performing the calculations, we needed to reformat the GWAS summary data of SCZ and IBD (including UC and CD) into pre-calculated LD scores referencing the European population data from the 1000 Genomes Project. SNPs with MAF < 1%, INFO score < 0.3, or were located in MHC regions (chr6: 28 477 797–33 448 354) were excluded (Yang et al., Reference Yang, Musco, Simpson-Yap, Zhu, Wang, Lin and Zhou2021). By using LDSC software, we assess the genetic correlation between SCZ and each of IBD, UC, and CD. LDSC estimates rg using the slope of the regression of Z scores on the LD-score (Bulik-Sullivan et al., Reference Bulik-Sullivan, Finucane, Anttila, Gusev, Day, Loh and Neale2015; Wu et al., Reference Wu, Xiao, Han, Zhang, Zhao, Hao and Jiang2022).
For LDSC analysis without the intercept constrained, p values < 1.25 × 10−2 (Bonferroni correction, 0.05/4) of rg estimates (taking values ranging from −1 to 1) were considered significant. Then we performed single-trait constrained intercept of heritability LDSC as sensitivity analysis under the assumption of no population stratification. Constrained intercept LDSC reduces the standard error of estimation compared to unconstrained LDSC, thus indirectly assessing the impact of inflated GWAS statistics. Nevertheless, if the intercept is misconstrained, it could also provide biased or misleading estimates of genetic correlation (Yang et al., Reference Yang, Musco, Simpson-Yap, Zhu, Wang, Lin and Zhou2021).
Cross-trait meta-analysis
We performed a cross-trait analysis via multi-trait analysis of GWAS (Python 2.7) to identify shared risk SNPs underlying the joint phenotypes (SCZ-IBD, SCZ-UC, SCZ-CD). MTAG is a generalized inverse variance weighted meta-analysis of multiple correlated traits, which obtains summary statistics from single-trait GWASs and outputs trait-specific association statistics, and p values from the meta-analysis can be used like those from single-trait GWASs. There is a central assumption of MTAG that all SNPs between traits have the same variance–covariance matrix. However, even if the assumption is not met, based on the analysis by the previous study (Turley et al., Reference Turley, Walters, Maghzian, Okbay, Lee, Fontana and Benjamin2018), MTAG is a consistent estimator with effect estimates that always have lower genome-wide mean squared errors than the corresponding single-trait GWAS estimates. We calculated maxFDR (the upper bounds on the false discovery rate) to test the hypothesis of equal variance-covariance for the shared SNP effect sizes under these traits. SNPs with p meta < 5 × 10−8 & single trait p < 0.05 were defined as significant shared risk SNPs. The combined variance effect predictor (VEP) was used to annotate the MTAG-identified independent variants (LD clumping r2 < 0.01 and a distance of 1000 kb, according to the 1000 Genomes European reference panel) with detailed functions.
Summary-data-based Mendelian randomization
The SMR analysis was conducted to discover pleiotropic genes in the association of SCZ with IBD, UC, or CD based on the results of the cross-trait meta-analysis. SMR mainly uses GWAS summary data with data from expression quantitative trait loci (eQTL) studies for the purpose of identifying genes whose expression levels are associated with complex traits due to pleiotropy (Zhu et al., Reference Zhu, Zhang, Hu, Bakshi, Robinson, Powell and Yang2016). In the current study, the eQTL data was obtained from gene expression data of 670 healthy individuals' whole blood tissues from the GTEx V8 database. Assessment of causality or pleiotropy of significant SMR associations by heterogeneity in dependent instruments (HEIDI) outlier test. Genes with FDR-corrected p value < 0.05 & p HEIDI > 0.05 were deemed significant. If the gene was not overlapped with the loci reported significantly by previous GWAS of SCZ and IBD (p < 5 × 10−8) and had not been reported as a SCZ or IBD functional gene by GWAS Catalog (https://www.ebi.ac.uk/gwas/) and PubMed (https://pubmed.ncbi.nlm.nih.gov/) retrievals, it was announced as a novel gene.
Co-expressed and KEGG pathway enrichment analysis
To obtain biological insights into novel pleiotropic genes, we calculated the co-expression of the novel genes identified between SCZ and IBD in healthy human whole blood tissues. Whole blood tissue gene expression data encompassing the expression of 56 200 genes in 758 healthy individuals were obtained from the GTEx V8 database. These 56 200 genes were then compared to the target genes using a linear regression model for genome-wide expression correlation analysis. Genes with p < 0.05 were considered to be significant and chosen for subsequent gene set enrichment analysis: Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis, conducted by the R package ‘clusterProfiler’.
Mendelian randomization
The selection of instrumental variables (IVs) in the MR analysis required the satisfaction of the following three assumptions, that genetic variants are strongly associated with exposure factors (association assumption) but not with confounding factors (independence assumption) and that they are associated with outcomes only through exposure (exclusivity assumption) (Emdin, Khera, & Kathiresan, Reference Emdin, Khera and Kathiresan2017). To satisfy the association assumption, genetic instruments for SCZ, IBD, UC, and CD were extracted from the above-mentioned GWAS summary data with a p value < 5 × 10−8. For Actinobacteria, the genetic instruments were derived from a GWAS including 8956 German individuals with p value < 1 × 10−5 (Rühlemann et al., Reference Rühlemann, Hermes, Bang, Doms, Moitinho-Silva, Thingholm and Franke2021), and for BMI, these were obtained from a meta-analysis with 700 000 participants and conducted by Genetic Investigation of Anthropocentric Traits (GIANT) consortium with p value < 5 × 10−8 (Yengo et al., Reference Yengo, Sidorenko, Kemper, Zheng, Wood and Weedon2018). To meet the independence assumption, the linkage disequilibrium (LD) r2 was set to be <0.001 in a 10 000 kb window. If an instrumental SNP was not available in the outcome dataset, a search was conducted through the LDlink website (https://ldlink.nih.gov) for a proxy SNP in LD with the target SNP (r 2 > 0.8). We also calculated the F statistic to evaluate the strength of the association between instrumental and exposure variables with the following formula: F = R 2 × (N − 2)/(1 − R 2) and excluded SNPs with F value less than 10 (weak instrumental variable). The R2 in the equation above denotes the exposure variance interpreted by each SNP separately, which was obtained by performing the following formula:

where β and EAF are the effect value and frequency of the effect allele, respectively, the N is the sample size of the SNP.
A bidirectional Mendelian randomization analysis was done to investigate whether there was a potential causal relationship of SCZ with each IBD, UC, and CD, or vice-versa. The inverse variance weighted (IVW) method was performed as the major MR analysis, and in the case that all these alternatives of IVs were valid, it combined the Wald ratio estimates of causal effects gained from the different variables to estimate the causal effects between the two traits. MR-Egger regression (Bowden, Davey Smith, & Burgess, Reference Bowden, Davey Smith and Burgess2015), weighted model-based estimate (Hartwig, Davey Smith, & Bowden, Reference Hartwig, Davey Smith and Bowden2017), and weighted median (Bowden, Davey Smith, Haycock, & Burgess, Reference Bowden, Davey Smith, Haycock and Burgess2016) were used to test the robustness of our findings under the loose model assumptions. To test for heterogeneity and pleiotropic effects, we conducted additional Cochran's Q test as well as MR pleiotropy residual sum and outlier (MR-PRESSO) test. We further examine the robustness of the results by performing a leave-one-out analysis by deleting each variant in turn from the MR analysis. The p value less than 8.33 × 10−3 (Bonferroni correction, 0.05/6) was considered statistically significant.
Multivariable Mendelian randomization (MVMR) is an extension of two-sample Mendelian randomization that allows simultaneous estimation of the causal effect of each risk factor on the outcome through a set of genetic variants associated with several measured risk factors (Burgess & Thompson, Reference Burgess and Thompson2015). Actinobacteria, one taxon of intestinal bacterial flora whose numbers are described by abundance, is a continuous variable (Rühlemann et al., Reference Rühlemann, Hermes, Bang, Doms, Moitinho-Silva, Thingholm and Franke2021). Several studies have shown both BMI and actinobacteria were associated with exposure (SCZ) and outcome (IBD) (Alexander et al., Reference Alexander, Ang, Nayak, Bustion, Sandy, Zhang and Turnbaugh2022; Gu et al., Reference Gu, Luo, Kim, Paul, Limketkai, Sauk and Singh2022; Ni et al., Reference Ni, Xu, Yan, Han, Zhang, Wei and Zhang2022; Yu et al., Reference Yu, Fu, Yu, Tang, Sun, Wang and Lu2023). Therefore, in the MVMR analysis, we added BMI and actinobacteria as risk factors to estimate the direct effect of SCZ, BMI, and actinobacteria on IBD risk. Duplicate and related IVs (r 2 > 0.001,10000 kb) were excluded, and then the IV effect values and standard errors in each summary statistic were extracted from the GWAS summary data of exposure and outcome for MVMR analysis. Notably, effect alleles should be consistent in the MVMR analysis. The bidirectional MR analysis and MVMR analysis were conducted by R packages ‘TwoSample MR’ and ‘Mendelian randomization’, respectively. The parts of the manuscript on MR analyses adhere to the STROBE-MR guideline (Skrivankova et al., Reference Skrivankova, Richmond, Woolf, Yarmolinsky, Davies, Swanson and Richards2021).
Results
SNP heritability and genetic correlation estimated by LDSC
We detected moderate to high heritability estimates for SCZ and IBD: 36% for SCZ, 30% for IBD, 24% for UC, and 42% for CD, respectively (online Supplementary Table S2). Significant positive genetic correlations (without intercept constrained) were found in all trait pairs (SCZ-IBD: rg = 0.14, p = 3.65 × 10−9; SCZ-UC: rg = 0.15, p = 4.88 × 10−8; SCZ-CD: rg = 0.12, p = 2.27 × 10−6). For comparison, a significant positive genetic correlation was observed between UC and CD, with rg = 0.63 (p = 3.26 × 10−73), details are seen in Table 1 and Fig. 2. These estimates were slightly declined between all trait pairs with constrained the LDSC intercept (SCZ-IBD: rg = 0.11, p = 1.50 × 10−9; SCZ-UC: rg = 0.11, p = 6.71 × 10−8; SCZ-CD: rg = 0.10, p = 7.19 × 10−7; UC-CD: rg = 0.42, p = 1.10 × 10−36).
Table 1. Genetic correlation estimated by LDSC

Abbreviations: LDSC, linkage disequilibrium score regression; SCZ, schizophrenia; IBD, inflammatory bowel disease; UC, ulcerative colitis; CD, Crohn's disease; SNP, single nucleotide polymorphism; λGC, genomic inflation factor.

Figure 2. Genetic correlation between SCZ and IBD, SCZ and UC, SCZ and CD, UC, and CD.
Abbreviations: SCZ, schizophrenia; IBD, inflammatory bowel disease; UC, ulcerative colitis; CD, Crohn's disease; LDSC, linkage disequilibrium score regression.
Multi-trait analysis of GWAS
After pruning, there were 82, 61, and 79 genome-wide significantly independent SNP associated with SCZ and IBD, UC, and CD, which were located at 64, 52, and 66 loci, respectively. The most significant SNP was rs35848276 at 6p22.1 for the joint-phenotype SCZ-IBD (p meta = 3.34 × 10−33) (online Supplementary Table S3). The maxFDR values of SCZ with IBD and its subtypes were about 4.52 × 10−7, suggesting that the possible deviation caused by violation of MTAG assumptions can be negligible. Detailed functional annotations of each significant variant identified by MTAG are shown in online Supplementary Table S4.
Summary-data-based mendelian randomization
On the basis of the findings of the cross-trait meta-analysis, between SCZ and IBD, SMR confirmed 66 genes, 28 of which were notified as novel pleiotropic genes (Table 2). SMR identified 80 potentially functional genes between SCZ and UC, of which 40 were recognized as novel genes, and 59 functional genes between SCZ and CD, of which 28 were deemed as novel genes (online Supplementary Table S5). Notably, 11 novel genes (including KAT5, RABEP1, ELP5, CSNK1G1, and TMED4, etc.), were identified in all joint phenotypes, details about the genes are shown in Table 2.
Table 2. Shared functional genes between SCZ and IBD identified by SMR

Abbreviations: SCZ, schizophrenia; IBD, inflammatory bowel disease; SMR, summary-data-based Mendelian randomization analysis; BP, base pair location; SNP, single nucleotide polymorphism; A1, effect allele; A2, reference allele; HEIDI, heterogeneity in dependent instrument.
a The boldened genes are novel genes.
Co-expression and KEGG pathway enrichment analysis
KEGG pathway enrichment analysis suggested that the 11 novel genes identified in all joint phenotypes and their respective co-expressed genes were mainly enriched in core immune-related signal transduction and cerebral disorder-related pathways, such as T-cell receptor signaling, Alzheimer's disease, Parkinson's disease, and neurotrophin signaling pathway (online Supplementary Fig. S1).
Mendelian randomization analysis
In total, there were 155, 17, 543, 117, 60, and 88 independent SNPs were selected as IVs for SCZ, Actinobacteria, BMI, IBD, UC, and CD, respectively (online Supplementary Tables S6–S9). The F statistic values ranged from 29.46 to 175.27 for SCZ, 19.52 to 25.83 for Actinobacteria, 28.44 to 1270.71 for BMI, 29.86 to 500.59 for IBD, 30.46 to 408.12 for UC and 30.14 to 489.55 for CD, respectively (online Supplementary Table S10). In univariable MR analysis, genetically liability to SCZ was associated with an increased risk of IBD (OR 1.11 per logOR change in SCZ, 95% CI 1.07–1.15, p = 1.85 × 10−6), UC (OR 1.11, 95% CI 1.06–1.16, p = 5.05 × 10−5), and CD (OR 1.13, 95% CI 1.07–1.19, p = 1.04 × 10−4), respectively (Table 3). The results were similar across different MR methods. However, no causal effect between genetic liability to IBD on SCZ was observed (online Supplementary Table S11).
Table 3. Mendelian randomization between SCZ and IBD as well as its subtypes with IVW method

Abbreviations: IVW, inverse-variance weighted; SCZ, schizophrenia; IBD, inflammatory bowel disease; UC, ulcerative colitis; CD, Crohn's disease; UVMR, univariable Mendelian randomization; MVMR, multivariable Mendelian randomization analysis.
a MVMR1, multivariable MR for schizophrenia and actinobacteria with risk of IBD and its subtypes; MVMR2, multivariable MR for schizophrenia and BMI with risk of IBD and its subtypes.
Although there was significant heterogeneity present, no horizontal pleiotropy was detected by the MR-Egger intercepts (all p > 0.05) (online Supplementary Tables S12 and S13). The MR-PRESSO distortion test results indicated that the effect estimates stayed stable with the adjustment of these outliers (all p > 0.05). Meanwhile, the sensitivity analysis, and leave-one-out analysis, confirm the robustness of our results (online Supplementary Table S14). The SNP effect magnitude and direction of SCZ with IBD and its subtypes are shown in online Supplementary Fig. S2.
Multivariable MR taking into consideration actinobacteria generated similar results, although the OR estimate was slightly larger, indicating a causal effect of SCZ on the risk of IBD independent of Actinobacteria (OR 1.11, 95% CI 1.06–1.16, p = 1.34 × 10−6), as well as its subsets UC (OR 1.11, 95% CI 1.06–1.17, p = 2.28 × 10−5) and CD (OR 1.12, 95% CI 1.06–1.19, p = 1.67 × 10−4). After accounting for BMI, the causal relationship also existed between genetically determined SCZ and IBD (OR 1.11, 95% CI 1.04–1.18, p = 1.84 × 10−3) or CD (OR 1.17, 95% CI 1.08–1.27, p = 1.11 × 10−4) (Table 3).
Discussion
By leveraging the GWAS summary statistics, this study systematically interrogates the overlap genetic architecture underlying SCZ and IBD. Our results supported the positive genetic correlation of SCZ with IBD, as well as with UC and CD. Using cross-trait meta-analysis, we identified several pleiotropic loci and genes for two illnesses, with some of the shared genes might contribute to therapeutic target discovery. In addition, MR analysis supported a causal role of genetic liability to SCZ on IBD risk. All these findings provide novel insight into the presence of comorbidity of IBD in SCZ patients from a genetic perspective.
Overall, the SNP-based heritability estimates were moderate to high for SCZ and IBD. A global genetic correlation of SCZ with IBD and its subtypes was observed, indicating that genetic components contribute to the comorbidity of SCZ and IBD, which either directly affects both traits via variants (pleiotropy), or through the causal effect of one trait on the other. These positive genetic correlations were generally in line with a previous study using another SCZ summary statistics from a mixed population (Tylee et al., Reference Tylee, Lee, Wendt, Pathak, Levey, De Angelis and Polimanti2022). In Tylee's study, besides SCZ, they also discovered significant genetic correlations between other psychiatric disorders (major depressive disorder and anxiety) and IBD. Moreover, we applied intercept-constrained LDSC to estimate the effect of population stratification. The estimated value of genetic correlation (rg) with constrained intercept was slightly lower than those with unconstrained intercepts in all joint phenotypes, consistent with a previous study (Yang et al., Reference Yang, Musco, Simpson-Yap, Zhu, Wang, Lin and Zhou2021). Recently, the cross-trait meta-analysis with enhanced statistic power has been widely used to decipher pleiotropic genetic loci or genes for SCZ and its high comorbidities. For example, Cai et al., reported 25 new independent SNPs responsible for the co-occurrence of SCZ and type 2 diabetes (Cai et al., Reference Cai, Sun, Liu, Chen, He and Wei2022).
Genetic correlation analysis estimated the magnitude of pleiotropy or causal overlap between SCZ and IBD, whereas MR analyses inferred the causal associations. Consistent with two previous bi-directional MR studies (Qian et al., Reference Qian, He, Gao, Fan, Zhao, Ma and Yang2022; Yang et al., Reference Yang, Yang, Zhang, Zhang, Chen and Zuo2023), we uncovered significant causal effects between SCZ and IBD, UC, and CD, but not vice versa. Although the general conclusion was consistent with our work, there were considerable differences in the process of data acquisition and methodology selection. To reduce the bias from population stratification, we chose the released largest SCZ GWAS summary statistics from European ancestry (Trubetskoy et al., Reference Trubetskoy, Pardiñas, Qi, Panagiotaropoulou, Awasthi and Bigdeli2022), and obtained summary statistics for IBD from the largest GWAS meta-analysis of mainly European ancestry (de Lange et al., Reference de Lange, Moutsianas, Lee, Lamb, Luo, Kennedy and Barrett2017). Moreover, we performed MVMR analysis to investigate whether the causality between SCZ and IBD remained after correcting for the effects of BMI and Actinobacteria. We found that the effect of SCZ on IBD risk attenuated after taking actinobacteria into account, suggesting that actinobacteria may be a mediator for the effect of SCZ on IBD risk. Of note, genetically predicted SCZ confers a causal effect on CD subtype in both univariable and multivariable MR, but not on UC subtype adjustment for BMI. A possible explanation for this is that building on the fact that BMI is a body-wide indicator, the symptoms of UC relate only to the mucosa and submucosa of the colon, whereas CD affects multiple regions of the gastrointestinal tract (Mentella, Scaldaferri, Pizzoferrato, Gasbarrini, & Miggiano, Reference Mentella, Scaldaferri, Pizzoferrato, Gasbarrini and Miggiano2020).
We identified 82 independent SNP, located at 64 loci between SCZ and IBD, suggesting shared biological mechanisms between the two traits. Functional annotation analysis highlighted that the loci of KIF21B, DRD2, and SHMT2 are involved in the potential pathogenesis. KIF21B is a kinesin protein in which mutation can lead to neurodevelopmental disorders (Asselin et al., Reference Asselin, Rivera Alvarez, Heide, Bonnet, Tilly, Vitet and Godin2020). Gialluisi et al., have identified KIF21B as a candidate gene for late-onset Parkinson's disease (Gialluisi et al., Reference Gialluisi, Reccia, Modugno, Nutile, Lombardi, Di Giovannantonio and Esposito2021). Aberrant synaptic pruning may cause brain disorders such as SCZ and anxiety disorders. DRD2, as a class of inhibitory dopamine receptors, activates cell-autonomous mechanisms of mTOR signaling to regulate synaptic pruning (Zhang et al., Reference Zhang, Lin, Huang, Zhao, Zhang and Yin2021). DRD2 also acts as a tumor suppressor, interacting with β-arrestin2, DDX5, and eEF1A2 to inhibit the activation of the NF-κB signaling pathway, as well as regulating the microenvironment to promote macrophage M1 polarization, triggering GSDME execution of pyroptosis and suppressing tumorigenesis (Lu et al., Reference Lu, Wu, Guo, Liang, Yin, Yin and Zhou2022). SHMT2, serine hydroxymethyltransferase 2, is associated with tumor progression and may drive colorectal cancer progression and metastasis through inhibition of β-catenin degradation (Liu et al., Reference Liu, Wang, Liu, Tan, Tao, Xiao and Zhao2021).
Indeed, our subsequent SMR analysis discovered 11 novel genes, such as KAT5, RABEP1, ELP5, CSNK1G1, and so on, associated with all joint phenotypes. KAT5, also known as TIP60, belongs to the MYST family of histone acetyltransferases (HATs) and is involved in a multitude of cellular pathways, including autophagy, DNA repair, RNA transcription, and learning memory, etc. KAT5 plays a critical role in the development, metabolism, and functioning of the nervous system and is involved in the processing of amyloid precursor proteins, making it a potent therapeutic target in Alzheimer's disease, as postulated by Li et al. (Li & Rasmussen, Reference Li and Rasmussen2020). RABEP1, RAB GTPase binding effector protein 1, located in internal vesicles and endosomes, engages in vesicle-mediated transport (Gong et al., Reference Gong, Hu, Jin, Yan, Liu, He and Wang2022). RABEP1 has been found to be strongly associated with Alzheimer's disease and serves as a candidate causative gene for this disorder (Novikova et al., Reference Novikova, Kapoor, Tcw, Abud, Efthymiou, Chen and Goate2021; Park et al., Reference Park, Pyun, Hodges, Jang, Bice and Kim2021). ELP5 is a core subunit of the elongator complex and is vital for maintaining its integrity and stability. The elongator complex plays an integral role in the cytotoxicity produced by gemcitabine, a first-line agent in chemotherapy for gallbladder cancer patients, by inducing apoptosis in gallbladder cancer cells. Inhibition of ELP5 expression may be responsible for the development of resistance to gemcitabine in patients with gallbladder cancer, leading to adverse effects (Xu et al., Reference Xu, Zhan, Jiang, He, Yang, Shen and Wang2019; Xu et al., Reference Xu, Jiang, Lin, Wang, Hu, Chen and Chen2021). CSNK1G1 generated a closed-loop structure of hsa_circ_101555 (circ101555) after back splicing. Silencing circ101555 expression in hepatocellular carcinoma cells and colorectal cancer cells was found to significantly inhibit the proliferation of carcinoma cells, induce their apoptosis, and impair their DNA repair ability (Chen et al., Reference Chen, Ren, Wan, Wang, Xue, Jiang and Zhi2019; Gu et al., Reference Gu, Zhang, Ran, Pan, Jia, Zhao and Yu2021).
Through the KEGG pathway enrichment analysis, we discovered that core immune-related signal transduction pathways (especially the ‘T-cell receptor signaling pathway’) were significantly correlated with the expression levels of the 11 novel genes and their respective co-expressed genes in whole blood. T cells are critical mediators of the adaptive immune response (Shah, Al-Haidari, Sun, & Kazi, Reference Shah, Al-Haidari, Sun and Kazi2021). Within the brain, T cells activate microglia (the brain's resident immune cells), inducing the production of pro-inflammatory cytokines and leading to neuroinflammation (Corsi-Zuelli & Deakin, Reference Corsi-Zuelli and Deakin2021; Debnath, Reference Debnath2015). Epidemiologic and GWAS studies support that immune system dysregulation underlies the core of neurodevelopmental and neurodegenerative processes in SCZ (Horváth & Mirnics, Reference Horváth and Mirnics2014; Purcell et al., Reference Purcell, Wray, Stone, Visscher, O'Donovan, Sullivan and Sklar2009). Increased neutrophil and monocyte counts, CD4/CD8 lymphocyte ratios, and levels of pro-inflammatory cytokines (IL1β, IL6, and TNFα) have been found in patients with psychotic episodes compared to patients in remission (Steiner et al., Reference Steiner, Frodl, Schiltz, Dobrowolny, Jacobs, Fernandes and Bernstein2020; Yuan, Chen, Xia, Dai, & Liu, Reference Yuan, Chen, Xia, Dai and Liu2019).
In clinical practice, whether the comorbidity of IBD in SCZ patients resulted from extrinsic or endogenous effects remains unclear. Our results suggest an intrinsic link underlying IBD and SCZ and provide new insight into the biological mechanisms. The shared genetic basis of these two complex disorders was attributed to pleiotropy and/or causality. The extrinsic effects are likely to exert major roles that are beneficial for psychiatrists and SCZ patients in clinical treatment. However, we acknowledged several limitations of the current study. First, given that all GWAS summary datasets were mostly derived from European populations, the findings might not be generalizable to other ethnicities. Given that the summary-level data of IBD are not purely European populations, this may had an impact on the results. However, in the LDSC analysis, we considered population stratification, which was shown to have less impact on the results of genetic correlation results. Second, the small sample size of GWAS summary data for actinobacteria may affect the results of our multivariable MR. Third, the current work is based on a series of comprehensive genetic analyses, actual functional experiments are needed to validate our findings. Finally, although genetic components partially explain the comorbidity between SCZ and IBD, its interaction with shared environmental factors is uncertain.
Conclusion
Our findings provide evidence of positive genetic correlation and shared loci/genes between SCZ and IBD, and suggested that genetic predisposition to SCZ was causally linked to increased risk of IBD. This work sheds novel insight into biological mechanisms and therapeutic targets underlying these two disorders.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0033291724000771
Availability of data and materials
All data generated or analyzed during this study are included in this published article and its supplementary information files. GWAS summary statistics for SCZ and IBD were available from the website of PGC (https://pgc.unc.edu/for-researchers/download-results/) and GWAS catalog (https://www.ebi.ac.uk/gwas/downloads/summary-statistics).
Acknowledgments
We thank all the study participants and research staff for their contributions and commitment to the present study.
Author contributions
JN and BD were responsible for the conception and design of the study. JN had full access to all the data in the study and took responsibility for the integrity of the data and the accuracy of the data analysis. JW, GYL, and TT performed the statistical analysis and drafted the manuscript. All authors were involved in drafting the article or revising it critically for important intellectual content, and all authors approved the final version to be published.
Funding statement
This study was supported by the National Natural Science Foundation of China (82103932), Research Fund of Anhui Institute of Translational Medicine (2023zhyx-C07), Key Project of Jiangsu Provincial Health Commission (ZD2021038) and Key Project for Social Development of Yangzhou (YZ2023147).
Competing interests
The authors declare that they have no competing interests.







