



Introduction
Titanoptera is an iconic insect group of the Triassic. The prominently large size (wingspan to 400 mm documented for Gigatitan vulgaris Sharov, Reference Sharov1968) and the raptorial forelegs equipped with stout spines make them the most impressive predatorial insects of the Triassic (Sharov, Reference Sharov1968; Grimaldi and Engel, Reference Grimaldi and Engel2005). Both sexes seemed to have had a ‘sound apparatus’ in the forewings, which might have produced deep and resonant acoustic signals in Triassic forests (Grimaldi and Engel, Reference Grimaldi and Engel2005; but see Schubnel et al., Reference Schubnel, Legendre, Roques, Garrouste, Cornette, Perreau, Perreau, Desutter-Grandcolas and Nel2021). Despite the fame, however, their phylogenetic affinity remains unstable. Although titanopterans are often considered closely related to orthopterans, their phylogenetic position has been the subject of debate. Sharov (Reference Sharov1968) argued that titanopterans originated from the Permian orthopteran family Tcholmanvissiidae Zalessky, Reference Zalessky1929, rendering this family paraphyletic. Subsequently, Gorochov (Reference Gorochov2001, Reference Gorochov2007) suggested the Carboniferous protorthopteran family Geraridae Scudder, Reference Scudder1885 as a sister group of Titanoptera. Béthoux (Reference Béthoux2007) reinvestigated the wing veins and concluded that the order Titanoptera is derived from the Tchomanvissidae. Recently, Huang et al. (Reference Huang, Schubnel and Nel2020), however, questioned the close relationship between the Tchomanvissidae and Titanoptera, stating that further investigation is required (but see Béthoux, Reference Béthoux2020). The incomplete understanding of the affinity is partly due to the small number of fossils with limited paleogeographic occurrences; to date, all titanopteran fossils have been known only from Central Asia (including European Russia) and Australia (Tillyard, Reference Tillyard1925; Riek, Reference Riek1954; Sharov, Reference Sharov1968).
The Nampo Group in the Chungnam Basin is situated in the southwestern part of the Korean Peninsula and consists of nonmarine fluvial, alluvial, and lacustrine successions (Egawa and Lee, Reference Egawa and Lee2009; Lee and Chung, Reference Lee and Chung2010). Although it is one of the largest Mesozoic basins in Korea, its depositional age has remained controversial. Paleontological evidence such as plant and choncostracan fossils have suggested Late Triassic to Early Jurassic as the age (Kim and Kimura, Reference Kim and Kimura1988; Kim, Reference Kim1989; Kim and Lee, Reference Kim and Lee2015). In contrast, radiometric dating from a volcanic deposit provided Jurassic ages (172 Ma) for the deposition of the Nampo Group (Koh, Reference Koh and Kee2006; Jeon et al., Reference Jeon, Cho, Kim, Horie and Hidaka2007). Kee et al. (Reference Kee, Kim, Kim, Hong, Kwon, Lee, Cho, Koh, Song, Byun, Jang and Lee2019) also regarded the Nampo Group as coeval to the Jurassic nonmarine deposits of the small-scale piggyback basins in other parts of the Korean Peninsula. Recently, Park and Noh (Reference Park and Noh2015) and Park et al. (Reference Park, Kwon, Kim, Hong and Santosh2018a) discovered that the formation process of the volcanics should be distinguished from the deposition of the Nampo Group and suggested that the Nampo Group was deposited during the Late Triassic to Early Jurassic, and subsequently, the Chungnam Basin further subsided at the Early–Middle Jurassic to accommodate the volcanic deposits (Park et al., Reference Park, Noh, Cheong, Kwon, Song, Kim and Santosh2019; Lee et al., Reference Lee, Park, Choi and Sim2021).
This study reports a new titanopteran genus from the Amisan Formation, Nampo Group, Chungnam Basin, Korea, which is the first discovery of a titanopteran fossil outside of Central Asia and Australia, contributing to a better understanding of the morphological diversity and paleogeographic distribution of this group of giant insects. In addition, because titanopteran fossils do not occur above the Triassic, the new discovery is relevant to the debate on the age of the Nampo Group.
Geological setting
The outcrops of the Chungnam Basin are distributed in three closely situated areas, namely (from west to east) the Ocheon, Oseosan, and Seongju areas (Fig. 1), which have even been recognized as isolated sediment fills in subbasins (Egawa and Lee, Reference Egawa and Lee2009). The Chungnam Basin includes the Nampo Group and the mid-Jurassic Oseosan Volcanic Complex that unconformably overlies the Nampo Group in the Oseosan area (Park et al., Reference Park, Kwon, Kim, Hong and Santosh2018a). The fluvial, alluvial, and lacustrine successions of the Nampo Group of the Chungnam Basin consist of five formations, in ascending order, the Hajo, Amisan, Jogyeri, Baegunsa, and Seongjuri formations (Choi et al., Reference Choi, Kim and Seo1987; Yang, Reference Yang and Kim1999; Egawa and Lee, Reference Egawa and Lee2009). The Hajo, Jogyeri, and Seongjuri formations are dominated by conglomerate and sandstone facies, whereas the fossiliferous Amisan and the Baegunsa formations comprise coal-bearing shale and sandstone, with a minor amount of conglomerate in the lower part of the Baegunsa Formation (Yang, Reference Yang and Kim1999). The Nampo Group is unconformably overlain by the Early–Middle Jurassic Oseosan Volcanic Complex, which consists of light gray to gray tuff and lapilli tuff, overlain by thick tuffaceous sedimentary rocks of gray to dark gray shale, sandstone, and conglomerate (Park et al., Reference Park, Kwon, Kim, Hong and Santosh2018a). The basement rocks of the Chungnam basin are largely of Neoproterozoic and Paleozoic metamorphic rocks (the Wolhyeonri complex) west of the Oseosan area, whereas Paleoproterozoic granite gneiss is prevalent to the east of the Oseosan area (Kee et al., Reference Kee, Kim, Kim, Hong, Kwon, Lee, Cho, Koh, Song, Byun, Jang and Lee2019) (Fig. 1). Various hypotheses have been proposed for the development of the Chungnam Basin, under the view that the Chungnam Basin is part of the Daedong Supergroup, which include development of half-grabens or a pull-apart basin from the late stage of the Triassic Songrim orogeny (Cluzel, Reference Cluzel1992), deposition in synorogenic (Songrim orogeny) piggyback basins (Chough et al., Reference Chough, Kwon, Ree and Choi2000), and syntectonic deposition during the early stages of the Jurassic Daebo orogeny (Jeon et al., Reference Jeon, Cho, Kim, Horie and Hidaka2007). Subsequently, Egawa and Lee (Reference Egawa and Lee2009) considered the ‘subbasins’ of the Chungnam Basin as piggyback basins formed during the Daebo orogeny. Park et al. (Reference Park, Noh, Cheong, Kwon, Song, Kim and Santosh2019) and Lee et al. (Reference Lee, Park, Choi and Sim2021) proposed a postcollisional extensional deformation in the Late Triassic for the basin development based on the assumption that an eastward extension of the Sulu collisional belt runs through this area (Oh, Reference Oh2006; Kwon et al., Reference Kwon, Sajeev, Mitra, Park, Kim and Ryu2009).
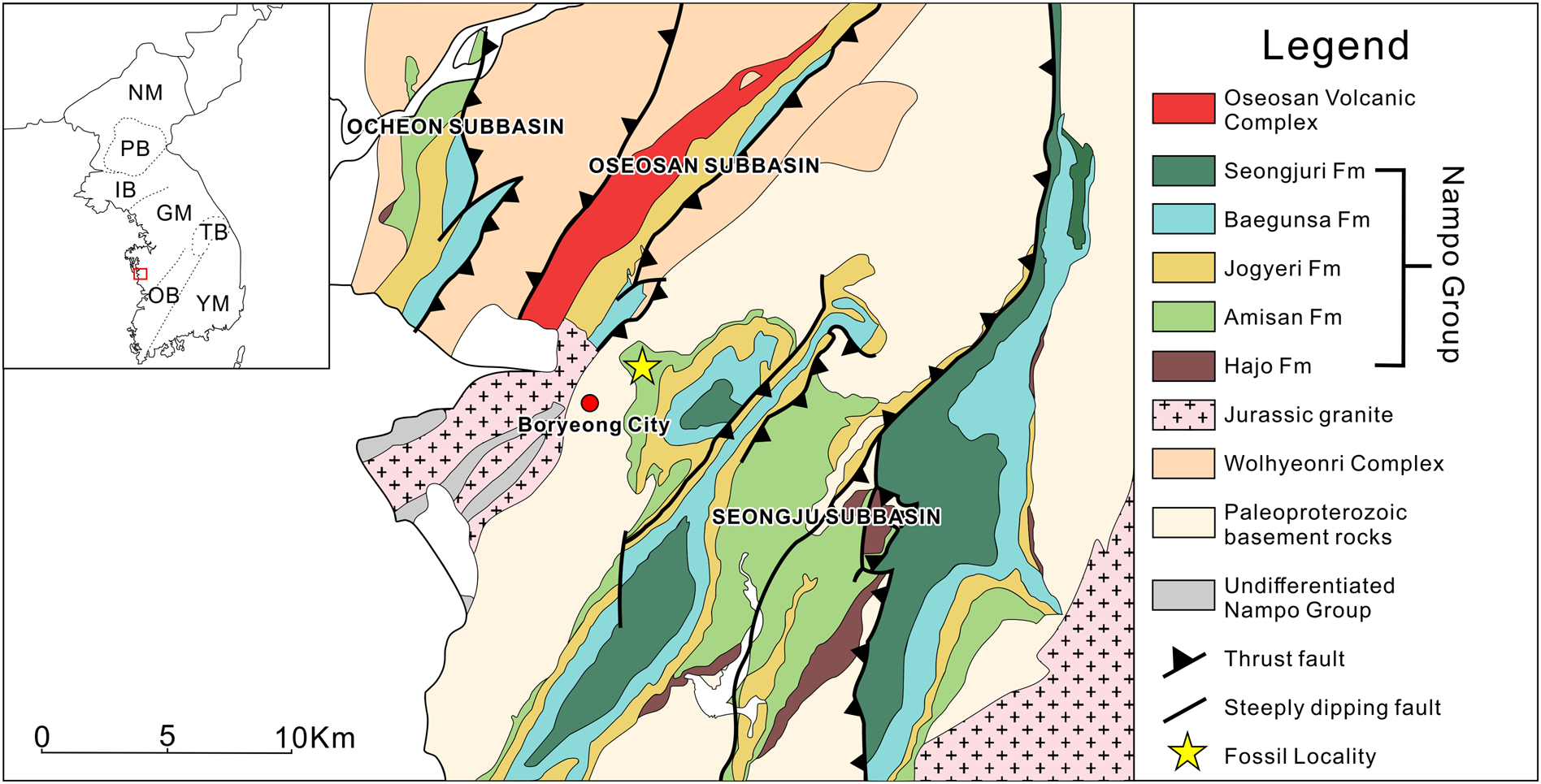
Figure 1. Geological map of the study area with its location on the Korean Peninsula, modified from Egawa and Lee (Reference Egawa and Lee2009), Kee et al. (Reference Kee, Kim, Kim, Hong, Kwon, Lee, Cho, Koh, Song, Byun, Jang and Lee2019), and Park et al. (Reference Park, Noh, Cheong, Kwon, Song, Kim and Santosh2019). GM = Gyeonggi Massif; IB = Imjingang Belt; NM = Nangrim Massif; OB = Okcheon Basin; PB = Pyeongan Basin; TB = Taebaeksan Basin; YM = Yeongnam Massif; star = fossil locality, Myeongcheon Section.
The Amisan Formation is known to 1,000 m thick and consists of the lower sandstone units, the lower shale unit, the middle sandstone units, the middle shale unit, and the upper sandstone unit (Yang, Reference Yang and Kim1999). Diverse fossils have been documented from the Amisan Formation, including plants (Kim, Reference Kim1976, Reference Kim1990, Reference Kim2001; Kimura and Kim, Reference Kimura and Kim1984; Kim et al., Reference Kim, Kim, Lee, Kim and Lee2002; Kim and Roh, Reference Kim and Roh2008), bivalves (Kim et al., Reference Kim, Lee and Choi2015), conchostracans (Kim and Lee, Reference Kim and Lee2015), insects (Nam and Kim, Reference Nam and Kim2014; Nam et al., Reference Nam, Wang, Ren, Kim and Szwedo2017), and fishes (Kim et al., Reference Kim, Lee, Park, Lee and Lee2020). Of these, plant and conchostracan fossils have been used for age-dating the Amisan Formation. Plate assemblages led Kimura and Kim (Reference Kimura and Kim1984) to suggest a Late Triassic age, but most of the genera later appeared to have ranges to the Lower Jurassic and even to the Cretaceous (Kim and Kimura, Reference Kim and Kimura1988; Kim, Reference Kim1990, Reference Kim2009, Reference Kim2013; Kim et al., Reference Kim, Kim, Lee, Kim and Lee2002; Lee et al., Reference Lee, Kim, Kim, Kim and Kim2004; Kim and Roh, Reference Kim and Roh2008). Conchostracan species, known from poorly preserved specimens, were also regarded as indicating Triassic age (Kim and Lee, Reference Kim and Lee2015), but the Mesozoic occurrence of the bivalve genus Margaritifera Schumacher, Reference Schumacher1816, the only known bivalve genus from the Amisan Formation, was then known to be restricted to the post-Triassic (Kim et al., Reference Kim, Lee and Choi2015). In addition, Jurassic radiometric ages of the Nampo Group (Koh, Reference Koh and Kee2006; Jeon et al., Reference Jeon, Cho, Kim, Horie and Hidaka2007) questioned the Triassic age derived from the paleontological evidence, calling for more convincing cues (Kim and Lee, Reference Kim and Lee2015).
Materials and methods
A single forewing with well-preserved venation was discovered in the middle shale unit of the Amisan Formation exposed in the Myeongcheon section (36°20’ 21”N, 126°37’34”E; Fig. 1). The sample was photographed submerged in water with a Canon EOS 60D using a Canon EF 100 mm f/2.8 USM macro lens. High-angle lighting was used for the maximal reflective image of the specimen. Images were cropped and enhanced by means of contrast and brightness in Adobe Photoshop CS6. For the relief-enhanced images, raw images for polynomial texture mapping (also known as Reflectance Transforming Imaging) were acquired by a self-crafted system that permits lighting from 50 different directions (see Béthoux et al., Reference Béthoux, Llamosi and Toussaint2016, Chen et al., Reference Chen, Gu, Yang, Ren, Blanke and Béthoux2021 for previous application of this method on insect fossils), and the same Canon EOS 60D camera setting mentioned above. The specimen was white-coated with magnesium oxide for this process. To enhance surface details of the fossil, 50 images were taken and converted into a PTM format file (see Supplemental Data), which was then run in RTI Viewer software (http://culturalheritageimaging.org/What_We_Offer/Downloads/).
The specimen was also gold-coated using a Cressington 108 Auto sputter coater with 10 mA for 80 s. A field-emission electron probe microanalyzer (JEOL JXA-8530F) and a low-vacuum field emission scanning electron microscope (JEOL JSM-7200F-LV) at the Korea Polar Research Institute were used to acquire X-ray elemental maps and surface analysis. X-ray elemental maps for C, N, O, Mg, Al, Si, P, S, Cl, K, Ca, Ti, Mn, Fe, Ni, Cu, Zn, and Ba were obtained using five wavelength dispersive spectrometry (WDS) and 19 energy dispersive spectrometry (EDS), with an acceleration voltage of 20 kV, beam current of 200 nA, beam size of 13 μm, dwell time of 10 ms, and step size of 13 μm. Raw data of X-ray elemental maps were imported and processed for brightness and contrast by ImageJ (Schneider et al., Reference Schneider, Rasband and Eliceiri2012).
General wing venation nomenclature follows that of Béthoux and Nel (2002; see also Desutter-Grandcolas et al., Reference Desutter-Grandcolas, Jacquelin, Hugel, Boistel, Garrouste, Henrotay, Warren, Chintauan-Marquier, Nel and Grandcolas2017). Interpretation of titanopteran-specific wing venation pattern follows Béthoux (Reference Béthoux2007). Anatomical abbreviations used in the text are: AA1 = first anal veins; AA2 = second anal veins; CuA = cubitus anterior; CuP = cubitus posterior; CuPa = anterior branch of CuP; CuPaα = anterior branch of CuPa; CuPaα• = first posterior branch of CuPaα; CuPaα* = second posterior branch of CuPaα; CuPaα° = anterior stem of CuPaα; CuPaβ = posterior branch of CuPa; CuPb = posterior branch of CuP; M = media vein; MA = media anterior, MP = media posterior; R = radius; RA = radius anterior; RP = radius posterior; ScA = subcosta anterior; and ScP = subcosta posterior.
Repository and institutional abbreviation
The specimen figured in this study is deposited in the Gongju National University of Education (GNUE).
Systematic paleontology
Superorder Archaeorthoptera Béthoux and Nel, Reference Béthoux and Nel2002
Titanoptera Sharov, Reference Sharov1968
Family Paratitanidae Sharov, Reference Sharov1968
Genus Magnatitan new genus
Type species
Magnatitan jongheoni n. gen. n. sp.
Diagnosis
As for the type species by monotypy.
Etymology
The generic name is derived from the Latin magna, referring to its large size, plus the general name of the group, titan.
Remarks
The two most important synapomorphies of titanopterans are CuPaα• + CuPaβ and CuPb having the same point of origin (Béthoux, Reference Béthoux2007), and the presence of strongly developed crossveins between MA and MP, both of which are conspicuous in the new specimen. The family Paratitanidae is characterized by the division of RA and RP near the distal half of the wing, and previously included two genera, Paratitan Sharov, Reference Sharov1968 and Minititan Gorochov, Reference Gorochov2007 from the Triassic of Kyrgyzstan. The location of the RA and RP division in the forewing justifies the assignment of Magnatitan n. gen. into the Paratitatnidae. The short length of the stem of M and fusion of CuA and anterior branch of CuPaα° make the new genus Magnatitan different from Minititan, and Paratitan in Paratitanidae. The presence of a single anterior veinlet in RA is distinct from the genus Paratitan with three or more anterior veinlets and a simple RA as in the genus Minititan.
Magnatitan jongheoni new genus new species
Figure 2
Holotype
GNUE 112001, an almost complete left forewing.

Figure 2. Holotype of Magnatitan jongheoni n. gen. n. sp., GNUE 112001: (1) overview of the specimen (rectangles, left to right, represent details shown in 1.3, 1.4, and 1.5, respectively); (2) interpretive drawing; (3) magnified image of the proximal part of the forewing; (4) wavelength dispersive spectrometry (WDS) silicon map, showing details around the division of CuPb; (5) PTM image showing CuA + CuPaα° and CuPaα°, and the cross-veins between; (6) PTM image, showing the distinctive cross-veins developed between MA and MP. AA1 = first anal veins; AA2 = second anal veins; CuA = cubitus anterior (in red); CuP = cubitus posterior; CuPaα = anterior branch of CuPa (in blue); CuPaα• = first posterior branch of CuPaα; CuPaα° = anterior stem of CuPaα; CuPaβ = posterior branch of CuPa (in orange); CuPb = posterior branch of CuP (in green); M = media vein; MA = media anterior (with its branches MA1 and MA2); MP = media posterior; R = radius (in pink); RA = radius anterior; RP = radius posterior; ScA = subcosta anterior; ScP = subcosta posterior; * (= CuPaα*) = second posterior branch of CuPaα; • (= CuPaα•) = first posterior branch of CuPaα. Large colored dots indicate fusion of veins.
Diagnosis
ScP with many strong veinlets, dichotomously ramified distally; RA extending a single anterior veinlet distally; stem of M short; MP with two branches; basal parts of CuPaα• and CuPaα* fused; basal part of CuPaβ fused with CuPb; CuPaβ + CuPb short; area between posterior wing margin widened.
Occurrence
The middle shale unit of the Amisan Formation, Upper Triassic, Myeongcheon Section, Seongju-myeon, Boryeong City, Chungcheongnam-do, Republic of Korea.
Description
Forewing large with likely pointed apex, anterior margin anteriorly bowed, posterior margin posteriorly bowed, preserved length 57.3 mm, width 19.0 mm; distal part of ScA preserved, weak; area between ScA and anterior margin narrow; ScP long, strong, anteriorly pectinate with many branches and dichotomously ramified distally; R convex; RA and RP divided near distal half of wing; RA with single, weak anterior veinlet; split of M and CuA preserved, stem of M 3.8 mm long; M gently twisted, MA1-MA2 split located distally with respect to RA-RP split; MP concave, with fork located between RA-RP split and MA1-MA2 split; area between MA and MP wide with dense, sturdy crossveins; CuA separated from M, free part of basal CuA short; basal part of CuPaα° divided into two branches; anterior branch of CuPaα° weak, fused with CuA like a crossvein; CuA + CuPaα° long, gently concave; posterior branch of CuPaα° posteriorly pectinated; fused CuPaα• + CuPaα* + CuP long; basal parts of CuPaα• and CuPaα* fused, CuPaα• + CuPaα* separated near widest area between MA and MP; basal part of CuPaβ fused with CuPb, CuPaβ + CuPb short; preserved AA1 and AA2 long without branches; crossveins dense, long, simple, forming network near wing margins and in broad intervenal areas.
Etymology
The species name honors the collector of the specimen, Professor Jongheon Kim (Kongju National University) who has made significant paleontological contributions from the Nampo Group over the last 30 years.
Remarks
Huang et al. (Reference Huang, Schubnel and Nel2020) argued that a divided CuPb is present in some members of Archaeorthoptera, Panorthoptera, and Titanoptera, and considered this feature in those groups as the result of convergent evolution, questioning the close relationship between the Titanoptera and the orthopteran family Tcholmanvissiidae. In contrast, Béthoux (Reference Béthoux2007, Reference Béthoux2020) reinterpreted the wing vein homology of those groups and proposed that the divided CuPb was not present in titanopterans, reinstating the close relationship of the Titanoptera and the Tcholmanvissiidae. Following the most recent interpretation by Béthoux (Reference Béthoux2020), we interpret that CuPaβ is fused into CuPb. In line with this, CuPaα• and CuPaα* also fuse with each other, as also appears in a species of Tcholmanvissiella Gorochov, Reference Gorochov1987 (see Béthoux, Reference Béthoux2007, Reference Béthoux2020).
In recent paleontological research using WDS analyses on fossiliferous shale, various body structures (e.g., brains in Cambrian stem-arthropod fossils [Park et al., Reference Park, Kihm, Woo, Park, Lee, Smith, Harper, Young, Nielsen and Vinther2018b] and wing venations and a subgenital plate in Cretaceous elcanid orthopteran fossils [Kim et al., Reference Kim, Lee, Nam and Park2021]) were highlighted in the carbon elemental maps by different carbon concentrations, indicating that the fossils were preserved as thin carbonaceous film on dark shale. The mineralogy of the new titanopteran fossil from the Amisan Formation in this study, however, seems different from those in the previous studies. Carbon distribution on the fossil surface is scarce (Fig. 3.3) and there is less silicon (Fig. 3.6) compared to the matrix. Instead, aluminum and potassium are enriched on the fossil surface (Fig. 3.11, 3.12).

Figure 3. Wavelength dispersive spectrometer (WDS) and energy dispersive spectrometer (EDS) images of the wing of Magnatitan jongheoni n. gen. n. sp. from the Amisan Formation: (1) normal image of the holotype, submerged in water; (2) PTM image; (3–6) WDS elemental maps of carbon (C), magnesium (Mg), iron (Fe), and silicon (Si), respectively; (7–12) EDS elemental maps of calcium (Ca), phosphate (P), oxygen (O), titanium (Ti), aluminum (Al), and potassium (K), respectively.
Along the thick wing veins, dendrite-like overgrowth textures are present (Fig. 3.1). Qualitative point analysis by EDS equipped at the low-vacuum field emission scanning electron microscope, however, has shown that the total EDS spectra of the wing membrane area, the wing veins, and the dendrite-like textures are comparable to each other. Similar results have been acquired from a XRD microstructural analysis applied to the wing and the rock matrix. In the elemental maps, a slight amount of magnesium and oxygen is present along the thick veins (Fig. 3.4, 3.9), whereas traces of titanium are recognized along the dendrite-like overgrowth (Fig. 3.10). There are prominent absences of iron, calcium, and potassium along the dendrite-like overgrowth (Fig. 3.5, 3.7, 3.12). At present, however, the mineralogy of the fossil is not clearly identified, requiring further research.
Discussion
First documentation of a titanopteran in East Asia
Compared to the large number of plant fossils (> 20 species reported), only a small number of invertebrate fossils have been documented from the Nampo Group, i.e., a single species of bivalve (Kim et al., Reference Kim, Lee and Choi2015), four species of conchostracans (Kim and Lee, Reference Kim and Lee2015), and two species of insects (Nam and Kim, Reference Nam and Kim2014; Nam et al., Reference Nam, Wang, Ren, Kim and Szwedo2017). The occurrence of the titanopteran fossil from the Amisan Formation, therefore, adds an important component to the Late Triassic ecology of the Nampo Group. Given its large size, Magnatitan jongheoni n. gen. n. sp. must have played an important role in the food web as a daunting predator on insects, invertebrates, and possibly small tetrapods, e.g., amphibians (Fig. 4). Previously, the occurrences of titanopterans have been restricted to Central Asia (Triassic Madygen Formation of Kyrgyzstan), Australia (Middle Triassic of New South Wales), and possibly European Russia (Permian of Vologda and Orenburg regions—Deinotitanidae, assigned to Titanoptera with some reservations; Gorochov in Aristov et al., Reference Aristov, Bashkuev, Golubev, Gorochov, Karasev, Kopylov, Ponomarenko, Rasnitsy, Rasnitsyn, Sinitshenkova, Sukatsheva and Vassilenko2013). A recent report documented a fragmentary titanopteran forewing from the Carboniferous of France (Schubnel et al., Reference Schubnel, Legendre, Roques, Garrouste, Cornette, Perreau, Perreau, Desutter-Grandcolas and Nel2021), but given the much earlier age and incomplete preservation of the fossil, further investigation is required. In Permo-Triassic paleogeography, Central Asia (including European Russia) was located north of the Tethys Ocean, Australia was located south of the ocean, and the two areas were connected by regions spreading along the western margin of the Tethys Ocean, e.g., northern Africa, Arabia, and India (Scotese, Reference Scotese2021). This implies that titanopterans might have also inhabited those regions along the western margin of the Tethys Ocean (Fig. 5). By the Late Triassic, the East Asian plates, including the Sino-Korean, South China, and Indochina cratons were accreted to form a large continental crust, located east of the Tethys Ocean (Scotese, Reference Scotese2021) (Fig. 5). Accordingly, the new discovery of titanopteran fossil from Korea could suggest a circum-Tethys Ocean distribution of titanopterans, at least in the Late Triassic. The possible widespread distribution of titanopterans implies a critical role played by this giant insect predator in Triassic terrestrial ecology. Future investigations on the Triassic nonmarine deposits in the circum-Tethys Ocean regions could lead to new discoveries of titanopterans.

Figure 4. Artistic reconstruction of Magnatitan jongheoni n. gen. n. sp., preying upon a small amphibian (artwork by Do-Yoon Kim).

Figure 5. A paleogeographic map of the Late Triassic with the occurrences of titanopterans (stars) and the inferred areas with possible distributions (marked by dotted lines) (modified from Scotese, Reference Scotese2021). AF = Africa; AR = Arabia; AT = Antarctica; AU = Australia; EU = Europe; IC = Indochina; IN = India; KOR = Korea; KRG = Kyrgyzstan; KZ = Kazakhstan; NA = North America; SA = South America; SC = South China; SKC = Sino-Korean (North China) Craton.
Triassic age of the Nampo Group
Titanopterans are known as predatory insects of the Triassic (Grimaldi and Engel, Reference Grimaldi and Engel2005), although an occurrence from the upper Permian has been documented (Gorochov, Reference Gorochov2007). The occurrence of a titanopteran, therefore, corroborates a Triassic age for the Amisan Formation. Previously, the family Paratitanidae to which the newly described genus belongs, included two genera, Paratitan and Minititan from the Middle–Upper Triassic Madygen Formation. The worldwide ‘coal accumulation gap’ interval is known in the Lower–Middle Triassic (Retallack et al., Reference Retallack, Veevers and Morante1996; Dai et al., Reference Dai, Bechtel, Eble, Flores, French, Graham, Hood, Hower, Korasidis and Moore2020; Shao et al., Reference Shao, Wang, Wang, Li, Wang, Li, Shao, Zhang, Gao and Dong2020), the cause of which is still debated (see Dai et al., Reference Dai, Bechtel, Eble, Flores, French, Graham, Hood, Hower, Korasidis and Moore2020). The frequent occurrences of coal-bearing layers in the Amisan and Baegunsa formations, therefore, constrains the age of these formations as post-Middle Triassic. Overall, the new discovery finally confirms a Late Triassic age for the Amisan Formation proposed previously by Kimura and Kim (Reference Kimura and Kim1984) and Kim and Lee (Reference Kim and Lee2015). In this regard, occurrences of the plant fossil genera Lobatannularia Kawasaki, Reference Kawasaki1927 and Sphenophyllum Brongniart, Reference Brongniart1828 from the Baegunsa Formation, which overlies the Amisan and the Jogyeri formations, could suggest that the Upper Triassic spans to the Baegunsa Formation. Kim and Lee (Reference Kim and Lee2015) and Kim and Kimura (Reference Kim and Kimura1988) even regarded the two genera as relic components from the Late Paleozoic Cathaysian flora of East Asia. If so, the uppermost unit of the Nampo Group, the Seongjuri Formation, remains the only unit with the possibility of stretching into the Lower Jurassic. The youngest detrital zircon data from the Seongjuri Formation shows a range from 197–175 Ma (Lee et al., Reference Lee, Park, Choi and Sim2021), supporting this possibility.
Data availability statement
Data available from the Dryad Digital Repository: https://doi.org/10.5061/dryad.q573n5tkz.
Acknowledgments
We are grateful to O. Béthoux and an anonymous reviewer for constructive comments that significantly improved the manuscript. Y. Oh helped us in the field. E. Shim helped by drawing the figures. This project was financially supported by the KOPRI project (PE22060). The authors declare that they have no known competing interests or personal relationships that could have appeared to influence the work reported in this paper.






 Open access
Open access