



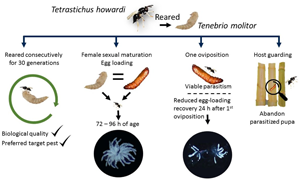
Introduction
Applied biological control (ABC) relies on the large-scale rearing of natural enemies that offer both quality (sex ratio, body size, female fecundity, fertility, parasitism/predation rate, etc.) and cost-effectiveness. The development of artificial diets and rearing methods for target pests (parasitoid hosts) is the first milestone for ABC (Leppla, Reference Leppla, King and Leppla1984). This step helped to mitigate contamination risks within the insectary, ensured a consistent supply of hosts/prey, and reduced rearing costs, among others (Leppla and Ashley, Reference Leppla and Ashley1989). Notably, the advancements in artificial diets and the refinement of rearing techniques have significantly contributed to ABC by enabling the effective use of parasitoids against various agricultural and veterinary pests (Morales-Ramos et al., Reference Morales-Ramos, Rojas, Coudron, Huynh, Zou, Shelby, Morales-Ramos, Rojas and Shapiro-Ilan2023).
Biological control plays a role in managing stalk (a.k.a. stem) borer pests (Lepidoptera: Crambidae) that affect crops like sugarcane, rice, and maize (Kondo et al., Reference Kondo, Manzano, Cotes, van Lenteren, Bueno, Luna and Colmenarez2020). Larvae of Crambidae such as of Diatraea spp. bore into host plant stems impairing control through insecticide applications. On the other hand, biological control agents can reach the pest within the plant stem, offering a sustainable and environmentally friendly approach to managing pests while reducing the reliance on chemical insecticides (van Lenteren et al., Reference van Lenteren, Bale and Bigler2003). Additionally, mass rearing and strategic release of beneficial organisms into agricultural ecosystems have been essential in protecting crop yield and enhancing food security (Settle et al., Reference Settle, Ariawan, Astuti, Cahyana, Hakim, Hindayana and Syahbudin1996). This approach enhances agricultural sustainability and reduces the ecological footprint of crop cultivation, underscoring its critical role in our efforts to meet growing global demands for food and bioenergy.
ABC has strengthened by the use of alternative hosts with reduced costs for different natural enemy species worldwide. For instance, Trichogramma spp. has been reared on eggs of Sitotroga cerealella (Olivier), Ephestia kuehniella (Zeller), and Ostrinia furnacalis (Guenée) (Pratissoli et al., Reference Pratissoli, Dalvi, Polanczyk, Andrade, Holtz and Nicoline2010; Li et al., Reference Li, Lei, Hua, Song, Wang, Ramirez-Romero, Dai, Li and Li2019; Moghaddassi et al., Reference Moghaddassi, Ashouri, Bandani, Leppla and Shirk2019). Furthermore, pupae of the yellow mealworm Tenebrio molitor L. have been used for rearing Sclerodermus guani Xiao & Wu to control the Japanese pine sawyer, Monochamus alternatus (Hope) (Coleoptera: Cerambycidae) (Hu et al., Reference Hu, Yang, Lin, Zhang and Dong2017). Lately, yellow mealworm pupa has been adopted to rear the parasitoids Trichospilus diatraeae Cherin & Margabandhu, Palmistichus elaeisis Delvare & LaSalle, and Tetrastichus howardi (Olliff) (Pereira et al., Reference Pereira, Pastori, Kassab, Torres, Cardoso, Fernandes, Oliveira, Kassab, Zanuncio, Parra, Pinto, Nava, Oliveira and Diniz2021), registered to release against lepidopteran defoliators of Eucalyptus and sugarcane stalk borers (AGROFIT, 2023).
T. howardi is a cosmopolitan, gregarious endoparasitoid that naturally parasitises different lepidopteran pests (Silva-Torres et al., Reference Silva-Torres, Pontes, Torres and Barros2010; Vargas et al., Reference Vargas, Pereira, Tavares and Pastori2011; Barbosa et al., Reference Barbosa, Kassab, Pereira, Rossoni, Costa and Berndt2015; Pereira et al., Reference Pereira, Kassab, Calado, Vargas, Oliveira and Zanuncio2015; Piñeyro et al., Reference Piñeyro, Pereira, Borges, Rossoni, Silva and Kassab2016). The life history and behavioural traits of T. howardi make it a candidate biocontrol agent against pupae of sugarcane stalk borers. Although currently used egg and larval parasitoids do not target pupae, parasitism does occur on the larvae of sugarcane borers (Vargas et al., Reference Vargas, Pereira, Tavares and Pastori2011; Pereira et al., Reference Pereira, Kassab, Calado, Vargas, Oliveira and Zanuncio2015). Released T. howardi find the sugarcane borer pupa inside the stem in the field (Kfir et al., Reference Kfir, Gouws and Moore1993; Barbosa et al., Reference Barbosa, Pereira, Motomiya, Kassab, Rossoni, Torres, Mussury and Pastori2019); they fulfil development and accomplish parasitism when reared at temperatures ranging from 18 to 34°C (Yan et al., Reference Yan, Su, Lyu, Lu, Tang, Zhuo and Yang2024). Additionally, they can be reared in a laboratory on pupae of different alternative host species (Barbosa et al., Reference Barbosa, Kassab, Pereira, Rossoni, Costa and Berndt2015; Piñeyro et al., Reference Piñeyro, Pereira, Borges, Rossoni, Silva and Kassab2016; Pereira et al., Reference Pereira, Pastori, Kassab, Torres, Cardoso, Fernandes, Oliveira, Kassab, Zanuncio, Parra, Pinto, Nava, Oliveira and Diniz2021), including pupae of T. molitor (Vargas et al., Reference Vargas, Pereira, Tavares and Pastori2011; Barbosa et al., Reference Barbosa, Pereira, Motomiya, Kassab, Rossoni, Torres, Mussury and Pastori2019; Tiago et al., Reference Tiago, Pereira, Kassab, Barbosa, Cardoso, Sanomia, Pereira, Silva and Zanuncio2019).
Even though T. howardi can be reared in different alternative hosts in the laboratory, T. molitor pupae are preferred for commercial rearing (Tiago et al., Reference Tiago, Pereira, Kassab, Barbosa, Cardoso, Sanomia, Pereira, Silva and Zanuncio2019; Machado et al., Reference Machado, Bermúdez, Vacari, Silva-Torres, Pereira and Torres2023). There are many advantages of using T. molitor pupae as a host for T. howardi: they are less demanding of dietary and rearing conditions, can be mass-reared easily, and are readily available commercially. In addition, the estimated cost of producing 1000 pupae of T. molitor, reared on starter poultry feed, is approximately 28.57 US dollars, resulting in an average production of 130.5 T. howardi parasitoids per pupa (≈130,000 parasitoids). This means the estimated cost per adult parasitoid is only 0.00022 US dollars (Machado et al., Reference Machado, Bermúdez, Vacari, Silva-Torres, Pereira and Torres2023). It's important to note that even though pupae from Lepidoptera species maintain a closer biological relationship with the target pest, the sugarcane borer, the alternative host T. molitor, a coleopteran species, serves as an excellent host for large-scale commercial rearing if it does not negatively impact the parasitoid's performance against the target pest.
T. howardi is a gregarious parasitoid belonging to the Eulophidae family. Gregarious eulophids exhibit the intriguing behaviour of remaining with the cryptic host after locating it, allowing for multiple oviposition events (Matthews et al., Reference Matthews, González, Matthews and Deyrup2009; Silva-Torres et al., Reference Silva-Torres, Pontes, Torres and Barros2010; Pereira et al., Reference Pereira, Guedes, Serrão, Zanuncio and Guedes2017). After the host location, a female T. howardi may engage in superparasitism to overcome larger hosts, potentially depleting her egg supply for further parasitism. A single female T. howardi attains better parasitism (% of parasitism and offspring production) attacking a median-sized host pupa (33.3–64.8 mg) (N.C. Bermudez, unpublished data), but also parasitises successfully larger hosts like sugarcane borers (≈160 mg) (Vargas, Reference Vargas2013). Beyond host size, external cues and the host's internal defence mechanisms can be affected by the natal host of the parasitoid (Silva-Torres et al., Reference Silva-Torres, Matthews, Ruberson and Lewis2005). To overcome host defence, female parasitoids may inject venom into the host after the initial oviposition. Venom injection enhances offspring survival by preventing egg encapsulation, melanisation, and counter-attacking the host's immune system (Pennacchio and Strand, Reference Pennacchio and Strand2006; Andrade et al., Reference Andrade, Serrão, Zanuncio, Zanuncio, Leite and Polanczyk2010). The learning process and memory acquisition, including pre-imaginal learning shortly after adult emergence, play a crucial role in host selection (Cortesero and Monge, Reference Cortesero and Monge1994; Gandolfi et al., Reference Gandolfi, Mattiacci and Dorn2003; Dauphin et al., Reference Dauphin, Coquillard, Colazza, Peri and Wajnberg2009). As a result, the parasitoid may develop a preference for the natal host, thus influencing pest control once released into the field. Thus, alternative hosts used for extended periods in rearing facilities may alter the parasitoid's ability to locate hosts and its overall parasitism performance in the field (Liu et al., Reference Liu, Wang, Zhao, Men and Wei2017).
This research aimed to examine behavioural characteristics of the parasitoid T. howardi and parasitism performance in response to continuous rearing on the alternative host T. molitor. The fitness and preference of T. howardi for Diatraea saccharalis pupae were evaluated after successive generations of parasitoid rearing on the alternative host. We also determined the egg load patterns in female T. howardi reared on D. saccharalis and T. molitor pupae and the number of ovipositions required to achieve viable parasitism. Additionally, the ability of female T. howardi to produce new eggs (egg load replenishment) after parasitism was evaluated. Finally, the influence of the natal host on the wing shape and parasitoid size variation were assessed as indirect parasitoid quality.
In this context, we tested four hypotheses: (i) rearing T. howardi up to 30 generations using the alternative host, pupae of T. molitor, would not affect its choice of the target host or influence parasitism performance; (ii) parasitism of sugarcane borer, a larger host pupa, will require T. howardi females successful oviposition, reducing the egg loading, hence restrain further host parasitism; (iii) T. howardi females would require a resting period after oviposition to re-establish egg loading; and (iv) T. howardi females, upon locating a sugarcane borer pupa inside the stalk, will choose to remain inside to avoid competition with other females and ensure successful parasitism.
Materials and methods
Insect population sources and rearing methods
A colony of the sugarcane stalk borer, D. saccharalis, was established in the ‘Laboratório de Controle Biológico de Insetos’ of the Universidade Federal Rural de Pernambuco, Recife, Pernambuco State, Brazil, using pupae donated by the ‘Associação de Plantadores de Cana da Paraíba (ASPLAN)’, located in the district of Pitanga da Estrada, Mamanguape, PB, Brazil. The colony was kept at 22 ± 1°C, 70 ± 10% relative humidity, and a photoperiod of 12:12 h L:D.
Larvae of D. saccharalis were fed with an artificial diet prepared after Hensley and Hammond (Reference Hensley and Hammond1968). The diet consisted of soybean meal, wheat germ, sugar, vitamin solution, Wesson salts, ascorbic acid, water, and anti-contaminants (Nipagin and antibiotics). The rearing procedures were according to Fonseca et al. (Reference Fonseca, Marques, Torres, Silva and Siqueira2015). Briefly, after hatching larvae were transferred to vials (7.5 × 14 cm2 in diameter × height) containing an artificial diet. After 30 days, larvae were transferred to plastic boxes (30 × 18 × 4 cm3, L × W × H), where they received this diet until pupation. Pupae were collected and transferred to acrylic boxes (8.5 × 8.0 cm2) lined with filter paper containing a wet cotton pad inside bottle caps until adult emergence. The adults were fed with a 10% honey-water solution, provided in moistened cotton in bottle caps. Adult rearing cages were cylindrical tubes of polyvinyl chloride measuring 22 × 20 cm2 (height × diameter) and lined with paper for oviposition. Eggs were immersed in a solution consisting of 1% copper sulphate and 3% formalin for 3 min, after which they were thoroughly rinsed with distilled water. After 5–6 days of incubation, eggs were placed in Petri plates (2 × 15 cm2, height × diameter), where larvae emerged to start a new rearing cycle.
The yellow mealworm, T. molitor, has been maintained in the laboratory as previously mentioned since 2016. Larvae and adults fed on a diet of 97% wheat flour and 3% yeast, as described in Torres et al. (Reference Torres, Zanuncio and Moura2006). In addition, slices of carrots, sweet potatoes, or sugarcane stalks are offered as part of the diet. Pupae served for parasitoid rearing and colony maintenance.
The parasitoid T. howardi colony initiated from parasitised D. saccharalis pupae donated by the ‘Laboratório de Controle Biológico de Insetos (LECOBIOL)’ located at the Universidade Federal da Grande Dourados, Dourados, MS, Brazil. After emergence, adult parasitoids were transferred to glass tubes (10 × 2 cm2, height × diameter) at a rate of seven females and a male per vial. The vials were sealed with plastic film and honey droplets were offered as food. The rearing procedure for T. howardi followed Vargas et al. (Reference Vargas, Pereira, Tavares and Pastori2011). Each vial with seven parasitoid females received one T. molitor pupa (≤48 h old) for 3 days. After that, the foundress females were discarded, and the pupae were reared until adult parasitoid emergence.
Sugarcane plants
Sugarcane plants (var. RB04-1443) were cultivated in cement rings (1.0 m diam. × 0.6 m height) filled with soil up to 20 cm to the surface. Plants were maintained in a greenhouse under natural conditions of 12–13 h photophase, 23–30°C, and were irrigated as needed. Harvesting begun when plants were 10 months old. Only the median portion of the stalks, which had a diameter of 3.5–4.0 cm, was used in the bioassays. Stalks were cleaned, transported to the laboratory, and prepared for the sugarcane borer larvae and parasitism by T. howardi.
Could the use of alternative host impair the parasitism performance of T. howardi?
This bioassay assessed the parasitoid performance after being continuously reared on T. molitor pupae. Initially, T. howardi was reared for five generations using pupae of the target pest, D. saccharalis. Thus, emerging adults were considered the F0 generation for the alternative host, T. molitor. Parasitoids reared on D. saccharalis pupae were compared to those reared exclusively on T. molitor pupae in future generations. Therefore, the performance of T. howardi was assessed at generations F5, F15, and F30 on T. molitor as the host. Because females of T. howardi at F0 were generated exclusively from D. saccharalis, they were allowed free choice of either host pupae and considered the control for future generations of continuous rearing. Thus, the following experiments were run: generation F0 – females emerged from D. saccharalis were allowed free choice for parasitism of either host pupae; F5, F15, and F30 – females emerged from continuous rearing on T. molitor pupae were allowed free choice for parasitism of either host pupae.
Experimental arenas were Petri dishes (12 × 2 cm2, diameter × height) lined with filter paper (n = 20), where the female parasitoid's preference between pupae of both hosts simultaneously was assessed. Host pupae offered were 24-h-old, similar in size and weight, to ensure similarity between treatments available. At 8 a.m., 2 h after the start of the photophase, one female parasitoid, 48-h-old, honey-fed, and mated, was released into the arena. Female parasitoids were continuously monitored after released into the arena and measured: (i) elapsed time to host first choice, (ii) host first choice, and (iii) host handling time. Once the female parasitoid made her choice (indicated by the introduction of her ovipositor), the unselected host pupa was removed from the arena. The selected pupa was reared to measure offspring produced, sex ratio, size of the female offspring, and calculate the parasitism rate.
Data of the choice test were subjected to analysis of deviance performed by generalised linear models (GLMs). The selection of the distribution utilised in the analysis was according to the nature of the data and the dispersion adjustment of the models. Thus, results referring to first choice, percentage of pupa parasitised, and effectively parasitised pupae were subjected to a GLM analysis using a binomial error distribution (link = logit). The time to first choice, the proportion of sex ratio, and morphometric variables were analysed using a GLM with a Gaussian error distribution (link = identity). When necessary to assess differences between generations of T. howardi that emerged in the same natal host, the data were subjected to a contrast analysis, making a comparison of means between the levels of this variable (F0, F5, F15, and F30) (α = 0.05). Finally, the variables of time spent on the pupa and the number of offspring produced were analysed using a GLM with a Poisson distribution (link = log) when it was necessary to adjust the model with a quasi-Poisson distribution of the error. A residual analysis was performed on each model to ensure the error distribution and model building were adequate. The analyses were performed using R software (R Core Team, 2020).
Geometric and traditional morphometrics of wasp wings and legs
Female wing shape and leg morphometry were measured by taking random samples of females that emerged per D. saccharalis pupa. Before taking measurements, females were frozen to death. Photos of the right forewing and right hindleg were taken using a digital camera connected to a stereomicroscope, a DIGILAB DI-150B (5× magnification). Image quality was enhanced using software GIMP V.2.10.30 when necessary. Images of the right forewings and hindlegs of females that emerged at the F0 (n = 42), F5 (n = 24), F15 (n = 36), and F30 (n = 36) generations were captured. To study wing shape variation 18 landmarks (LMs) (fig. 1S) were defined and digitised using TPSUtil v.1.74 and TPSdig2 v.2.30 (Rohlf, Reference Rohlf2017). Before running the morphometric analysis, all traits were digitised twice to test every LM against definition error through the performance of a Procrustes analysis of variance (ANOVA) using MorphoJ v.1.07a (Klingenberg, Reference Klingenberg2011). Using the same software, a generalised Procrustes superposition analysis was performed on wings removing the influence of size, position, and orientation to shape variables (Rohlf and Slice, Reference Rohlf and Slice1990). With these variables, a covariance matrix of the individual shapes was obtained to proceed with the corresponding multivariate analyses.
A principal component analysis (PCA) was carried out individually for each of the digitised trait covariance matrices to replicate and visually represent the shape space (Pearson, Reference Pearson1901). Following the identification of related groupings, the classifier generations (F0, F5, F15 and F30) group was used to conduct a discriminant analysis using the canonical variate analysis. Mahalanobis and Procrustes distances were used to calculate a permutation test (10,000 runs) between groups and display the corresponding P-values. Multivariate regression analysis was used with Procrustes coordinates as the dependent variable and the size centroid as the independent variable to investigate the impact of size on the wing shape (allometry) (Monteiro, Reference Monteiro1999).
For traditional morphometry, the size of the wings and legs was determined from LMs 1–15, whereas wing width considered LMs 13–18 (fig. 1S). Tibia size was the length between the joints of the tibia with the femur and the tarsus.
Egg loading in T. howardi as a function of age, natal host, and ovipositions
Ageing and natal host (assay I)
The pre-oviposition period and reproductive output were determined by the egg load of the female parasitoid. Both D. saccharalis and T. molitor were used as parental hosts. Female parasitoids were dissected under a light stereomicroscope MOTIC SMZ-168 (5× magnification) to account for eggs developing in the ovaries. Parasitised pupae of D. saccharalis or T. molitor were held in glass vials for parasitoid emergence, upon which honey droplets were offered on the inner walls of the vial as food for emerging adults. Mating was allowed until the time of the experiment. Females of six different age intervals (0–24, 24–48, 72–96, 96–120, and 120–144 h old) were collected and dissected for egg loading. Females accounted for a specific interval were those that emerged before 10 a.m. of the upper time limit (i.e. 0–24 h old, honey-fed, and mated before 10 a.m. of the first 24 h of emergence). For data characterisation, females from each age interval were considered as 24, 48, 72, 96, 120, and 144 h old. In each age interval, females were aspirated into 1.5-ml microcentrifuge tubes, frozen to death at −10°C, and later dissected. With the help of a razor blade, we cut open the abdomen of the female, and its contents were extracted using two needles. A cover slip was placed over the female's abdomen, gently pressed down, and the slide containing the female was observed under the microscope to count the number of fully developed eggs in the oviducts.
Superparasitism on a host pupa (assay II)
We measured the number of females' viable ovipositions and the egg-loading recovery following parasitism. Females of T. howardi, 48 h old, mated, and honey-fed, emerged from one of the two parental hosts, D. saccharalis or T. molitor, were evaluated. We recorded the number of ovipositions per 24–48-h-old pupa of D. saccharalis. Host pupae were placed in Petri dishes (3.5 × 1 cm2 in diameter × height) and offered to parasitoids 24 h later. The next day, each Petri dish received a 48-h-old female parasitoid. It was observed continuously during 10 h of photophase (8 a.m. to 6 p.m.) for host oviposition. Females were assigned to oviposit up to one, two, or three times on the same pupa (treatments). An oviposition event consists of the female mounting on the pupa, ovipositor insertion into the host, ovipositor extraction, and finally moving away from the host (Rodrigues et al., Reference Rodrigues, Pereira, Barbosa, Silva-Torres and Torres2021). There were ten replicates (females) per each oviposition (1–3×) treatment. After oviposition on the host pupa, parasitoid females were collected, stored in a 1.5-ml microcentrifuge tube, and stored at −10°C until dissection. In the control group, we used naïve 48-h-old mated females, and honey-fed were killed at −10°C for dissection and egg count.
Parasitised pupae were reared to assess the parasitism rate and number of offspring produced per pupa. Additionally, the parasitism rate also considered dead pupae with mummified parasitoids inside. This information provided insights into unviable parasitism due to host nutrition and defence mechanisms.
As described in the previous test, female parasitoids emerged from both D. saccharalis and T. molitor were offered a D. saccharalis pupae in a Petri dish, and were observed for host parasitism during 10 h of the photophase. Females that successfully completed one, two, or three ovipositions were removed from their hosts and allowed a 24-h rest period without contacting a new host. After this resting period, these females were frozen at −10°C and dissected to count the number of eggs in their ovaries (see fig. 2S). As a control group, 48-h-old honey-fed and mated naïve females were used.
Sequential oviposition on different host pupa (assay III)
The performance of T. howardi was measured after successive ovipositions. Thus, 96-h-old mated, honey-fed females were allowed two sequential ovipositions within a 10-h photophase of continuous observations. Females were single-released into Petri dishes (3.5 × 1 cm2, diameter × height) containing one pupa of D. saccharalis 24–48 h old. Right after parasitism, the pupa was removed from the dish, and another non-parasitised D. saccharalis pupa was offered to allow a second oviposition by the same parasitoid wasp (n = 31). The observations lasted for 10 h of the photophase. After that, each pupa with a parasitism attempt was reared separately to measure either adult parasitoid or sugarcane borer emergence. Pupae with no emergence of parasitoid or host adult were dissected to certify the cause of the mortality.
The number of eggs in the ovaries of naïve females reared from D. saccharalis or T. molitor was subjected to regression analysis using PROC REG of SAS (SAS Institute, 2002) as a function of the age of the parasitoid females (24, 48, 72, 96, 120, and 144 h). Furthermore, to test the hypothesis that females have differences in egg load due to the natal host, the linear portion (slopes) of fitted models was compared between parental hosts using PROC MIXED to test the equality of the linear slopes (SAS Institute, 2002).
The number of eggs remaining in the ovary of T. howardi females after zero, one, two, or three sequential ovipositions on D. saccharalis pupa; the number of offspring produced per parasitised pupa; the number of eggs present in the parasitoid female ovary after 24 h of recovering from one, two, or three ovipositions; and the offspring production per parasitised pupa from these three sequential ovipositions were transformed into square root (x + 0.5) to meet the ANOVA assumptions and subjected to a two-way ANOVA with natal host and number of ovipositions as factors (SAS Institute, 2002). Furthermore, the percentage of parasitism and mortality due to the parasitism, and the number of parasitoids produced per parasitised pupa were tested for the hypothesis of equal performance between the first and second oviposition by a single T. howardi female using a chi-square test at 0.05 significance levels.
Does the T. howardi female stay with the sugarcane borer pupa?
To understand the parasitism of the sugarcane borer pupae inside the sugarcane stalk by T. howardi, pieces of stalks 18–20 cm in length with nodes at both ends were cut from ≈5 cm diameter stalks. A gallery 3 cm deep was made in the stalk section between the nodes using a power drill. After that, the stalk was cut vertically into two halves, and one sugarcane borer larva was placed inside the gallery at the deepest point. Next, the two halves of the stalk were juxtaposed with rubber bands to avoid any opening other than the hole made with the drill, mimicking the natural entrance of sugarcane borers into the stalk. The larvae were 30 days old and, therefore, maintained feeding for about 5 days before moulting to pupa, which allowed the recreation of natural conditions within the gallery, including feeding residues and faeces. The stalk sections were positioned vertically within an 80-ml plastic pot that contained moistened cotton to delay decay. Each stalk section served as one observation unit. The stalk was covered by a 3-litre plastic bottle cage with two lateral openings fixed with organdie fabric for ventilation. Once the larvae reached the pupal stage, six units of stalks were placed vertically on a Styrofoam® board 50 × 32 cm2 (L × W) fixed inside a searching cage made with transparent Plexiglass® 50 × 45 × 32 cm3 (L × H × W). There were ten replicates (ca. 10 cages and 60 units). We measured the number of host findings and parasitism in each unit, which received 84 parasitoid females at a rate of 14 females per available pupa.
The assessments in the cages were conducted in the dark, with the help of red light, from 7 to 8 p.m., to reduce disturbance of the parasitoids. Evaluation days were 3, 6, and 9 days after releasing the female parasitoid into the cages. During each evaluation, we noted the number of parasitoid females on the pupa or inside the gallery, and the stalks were marked to indicate the presence or absence of the parasitoid female. Stalks containing parasitoid females with pupae were promptly sealed and returned individually to the observation units. They were monitored daily until the 9th day to check if the females stayed or abandoned the gallery. On the 9th day of exposure, we opened the stalks and counted the number of parasitoid females inside. The pupae were transferred to glass vials and reared until the adult emergence, either moths or parasitoids. Furthermore, we recorded the number of emerging parasitoids per pupa and the sex ratio of the offspring.
Results
Could the use of alternative host impair the parasitism performance of T. howardi?
T. howardi located and parasitised D. saccharalis pupae even after 30 generations of rearing in the alternate host, T. molitor pupae. Females emerging from T. molitor pupae preferred D. saccharalis pupae over T. molitor pupae (χ 2 = 94.33, P < 0.0001). Furthermore, across all generations evaluated for rearing the parasitoid in the alternate host, the preference for the target host remained consistent (fig. 1). The time required for the initial selection for parasitism remained relatively consistent across the generations reared in the alternative host except generation F0, where D. saccharalis pupae served as both the natal host and target host. Females from the F0 generation made that first choice faster than those from the F5, F15, and F30 generations (table 1). The other annotated variables, such as handling time, parasitism rate, the number of progeny produced, and the sex ratio, did not portray changes across the generations reared using the alternative host (table 1).

Figure 1. Females of T. howardi (n = 20) were allowed to choose between pupae of D. saccharalis or T. molitor after being reared continuously with the alternative host T. molitor pupae for 30 generations. Bars represent the percentages (values between parentheses) of females choosing either pupae over an observation period of 10 h. Chi-squares and respective P-values are given inside bars for pair-wise comparisons.
Table 1. Biological characteristics of T. howardi progeny obtained at different generations parasitising D. saccharalis pupae after continuous rearing in the alternative host T. molitor

a F0 stands for females emerged from D. saccharalis pupae and offered pupae of D. saccharalis (DD) for parasitism.
Geometric and traditional morphometrics of wasp wings and legs
The morphometric analysis showed that rearing T. howardi over several generations on T. molitor had no impact on the size of the female's wing (F 3, 134 = 1.14, P = 0.32, fig. 2A) by comparing the maximum length to the width of the forewing. Females' hindleg tibias, on the other hand, varied through generations (F 3, 134 = 7.10, P < 0.0001, fig. 2B), with larger tibiae for females from the F5 and F30 generations.

Figure 2. Size of the forewing (A) and hindleg tibia (B) of T. howardi females parasitising D. saccharalis (F0) and after different generations of continuous rearing using pupae of T. molitor (F5–F30). Bars with different letters indicate statistical significance means compared by a contrast analysis (P < 0.05).
The Procrustes ANOVA revealed that the group's mean square (MS) and F-values of error (MS = 0.000012, F = 0.69) were lower than those of the individuals (MS = 0.00016, F = 13.09) in the geometric morphometric analysis. It indicates that the LMs' digitisation was done correctly. The first three principal components (PC1 = 36.3%, PC2 = 14.2%, and PC3 = 9.1%) accounted for about 59.6% of the total variance in wing shape, according to the PCA generated for the wing shape. While the PC2 more clearly separates the groups (both for host and generation) (fig. 3A, B), the PC1 shows how the wing shape variation regarding the host switches between Diatraea–Diatraea (DD) and Tenebrio–Diatraea (TD) (fig. 3A) and throughout generations (fig. 3B). The distal portion of the wing (LM pointing arrows) is where the most noticeable alteration in the wing shape was observed in individuals of the F0 generation (DD) (fig. 2S). Although there is less variation in the wing morphometry of generations F5, F15, and F30, it still closely approaches the normal wing shape associated with PC1 (fig. 2S). The first composed of female wings from the F5, F15, and F30 generations, while the other was female wings from the F0 generation (DD) (fig. 4). According to the Mahalanobis and Procrustes distances, there were notable differences in the form of the wings (table 1S). Nevertheless, multivariate regression analysis did not reveal a relationship between wing size and shape (P = 0.63), with wing size accounting for only 0.48% of the variance in wing shape. In summary, there are no differences in wing shape between individuals.

Figure 3. Representation of the PCA for data of wing of T. howardi females reared for 30 generations using the alternative host, T. molitor, as a function of host switch (A) and at each of 30 generations accomplished (B) captured by the PC1 and PC2 axes. Dotted lines with black dots (i.e. LM) stand for change in shape and grey line stands for the average of shape associated with the PCs. Each point in the dispersion graphics represents the shape of the wing. Host switch from D. saccharalis to D. saccharalis (DD) at F00 generation, and from T. molitor to D. saccharalis (TD) at F05, F15, and F30 generations.

Figure 4. Canonical variate analysis. Differences in the average of wing shape generation of T. howardi that were reared during 30 generations on alternative host T. molitor.
Egg loading in T. howardi as a function of age, natal host, and ovipositions
Ageing and natal host (bioassay I)
The egg load increased linearly as a function of female age regardless of the parental host, D. saccharalis or T. molitor pupae (fig. 5). The pattern of egg loading by naïve T. howardi females was consistent with female age (PROC MIXED of SAS for equality of linear coefficient, b1–b2 = −0.027, df = 1, 115, t = −0.43, P = 0.668) (fig. 5). The mean number of eggs (+SE) found in the ovary at a mature age for females emerging from D. saccharalis and T. molitor pupae was 107.0 ± 2.9 and 99.5 ± 3.5 eggs, respectively.

Figure 5. Egg loading by naïve females of T. howardi reared using the target host, D. saccharalis (y = 44.8 + 0.435x, r 2 = 0.72, F 1, 58 = 144.62, P < 0.0001) or the alternative host, T. molitor (y = 39.45 + 0.46x, r 2 = 0.69, F 1, 58 = 73.83, P < 0.0001). Dashed lines stand for 95% confidence interval bands.
Superparasitism on a host pupa (bioassay II)
The number of offspring produced by a single T. howardi female that emerged from either natal host was similar (fig. 6A). These included parasitising the target host pupa (F 1, 54 = 0.53, P = 0.47), and sequential ovipositions (F 2, 54 = 0.26, P = 0.76). There was no interaction between these factors (F 2, 54 = 0.65, P = 0.52). Conversely, the remaining eggs in the female ovaries differed between natal hosts (F 1, 72 = 6.61, P = 0.012). Female parasitoids emerging from D. saccharalis pupae had more eggs in their ovaries than those emerging from T. molitor pupae when T. howardi oviposited twice in the same pupa (fig. 6B). Additionally, the number of eggs in the ovaries was considerably fewer in females that performed one to three ovipositions than in those that did not (F 3, 72 = 143.87, P < 0.0001). Despite that, the results indicate that there is no interaction with the paternal host and that eggs that remain in the female ovary undergoing sequential oviposition (F 3, 72 = 0.62, P = 0.061).

Figure 6. Mean number (+SE) of offspring produced per parasitised pupa of D. saccharalis after one to three observed ovipositions in the same pupa by a single female T. howardi over 12 h-photophase period (A and C), and remaining eggs in the ovary of these females dissected without oviposition (0) or right after completing one to three ovipositions (B) or after 24 h-recover period without oviposition (D). Note: P-values stand for comparisons between parental hosts; see text for details of the statistics.
Females originated from either natal hosts and, after completing two or three sequential ovipositions, had a similar number of eggs remaining in their ovaries. Additionally, these females produced the same number of offspring per parasitised D. saccharalis pupa whether they carried out one, two, or three successful ovipositions on the same pupa (fig. 6C), regardless of the natal host (F 1, 54 = 0.04, P = 0.81), the order of ovipositions (F 2, 54 = 0.31, P = 0.73), or any interaction between these factors (F 2, 54 = 2.23, P = 0.12). Additionally, when females performed one, two, or three sequential ovipositions and, then, had a 24-h interval without parasitism, the number of eggs found in their ovaries changed depending on the natal hosts (F 1, 54 = 4.63, P = 0.03) and the order of the ovipositions (F 2, 54 = 5.44, P = 0.007), but not when these factors were combined (F 2, 54 = 1.82, P = 0.17) (fig. 6D). Females that emerged from the alternative host, T. molitor pupa, had more eggs in the ovary after completing one oviposition and allowed a 24-h recovery period (fig. 6D).
Sequential oviposition on different host pupa (bioassay III)
After the first oviposition, parasitoid females were less likely to parasitise pupae. In fact, only 48.4% of the females were able to do a second oviposition within the 10-h photophase period (table 2). Females that oviposited for the second time successfully killed almost all pupae (93.3%). However, only 35.7% of these killed pupae in the second oviposition produced offspring. Furthermore, there was about a 50% reduction in offspring production from parasitised pupa in the second oviposition.
Table 2. Parasitism performance of T. howardi females accomplishing sequential ovipositions upon D. saccharalis pupae

Values between brackets refer to the raw numbers of pupae.
a Asterisks indicate that better performance was obtained for first host oviposition.
Does the T. howardi female stay with the sugarcane borer pupa?
Two pupae moved out of the stalk during the metamorphosis from 60 larvae that were placed within sugarcane stalks, leaving 58 pupae inside the stalk subject to parasitism. During the nocturnal check of these pupae, five (ca. 8.6%) were found with T. howardi females inside the sugarcane stalk. However, within a day, these parasitoids abandon the pupa. Thirty-three of the 58 pupae (ca. 56.9%) died inside the stalk, with viable parasitism accounting for 45.3% of those pupae and 11.6% of pupae without parasitoid emergence. From parasitised pupae with emergence, an average of 137.1 ± 12.8 parasitoids were produced, with a sex ratio of 0.89 (ca. 89% females), and the overall parasitism rate with offspring emergence was, on average, 45.3 ± 10.4% (mean ± SE).
Discussion
Enhancing the biological control of different target pest species with inundation releases of T. howardi will require large-scale rearing, which can be accomplished with an alternative host. However, the use of an alternative host for extended periods may result in conditioning to that host (Antolin et al., Reference Antolin, Bjorksten and Vaughn2006; Henry et al., Reference Henry, Roitberg and Gillespie2008; Li et al., Reference Li, Wey, Liu and Sun2010), which may impair parasitism performance (Jones et al., Reference Jones, Bilton, Mak and Sait2015; Samková et al., Reference Samková, Raška, Hadrava and Skuhrovec2021). Despite that assumption, T. howardi did not change its choice towards D. saccharalis or was less effective at parasitising it. Furthermore, one oviposition of the parasitoid on the pupa of D. saccharalis was enough to achieve success in parasitism. However, one oviposition by a female considerably reduces the egg load, and the female could not recover the egg load within 24 h of rest. Nonetheless, it doesn't seem that the absence of eggs in the ovary has much of an impact on the parasitoid's female dispersal.
Parasitoids use chemical and physical cues to locate and accept a host. Therefore, we expected that the natal host would have some influence on the parasitoid's performance on the target host (Godfray, Reference Godfray1994; Poelman et al., Reference Poelman, Bruinsma, Zhu, Weldegergis, Boursault, Jongema, van Loon, Vet, Harvey and Dicke2012; Becker et al., Reference Becker, Desneux, Monticelli, Fernandez, Michel and Lavoir2015). But, across all experiments using the alternative host, T. molitor, the parasitoid T. howardi consistently preferred D. saccharalis pupae, attaining comparable or even superior parasitism outcomes on D. saccharalis pupae. Similar results were found by Woltering et al. (Reference Woltering, Romeis and Collatz2019). These authors observed no deterioration in the performance of the parasitoid Trichopria drosophilae Perkins (Hymenoptera: Diapriidae) when reared for several generations on Drosophila melanogaster Meig. (Diptera: Drosophilidae) as an alternative host and subsequently exposed to the target host, Drosophila suzukii Matsumura (Diptera: Drosophilidae), under laboratory conditions. As a result, our findings suggest that rearing T. howardi in a coleopteran alternative host for up to 30 generations, despite being taxonomically distant from the field target hosts (Lepidoptera), will not affect its biological performance, and can be used as an alternative host for large-scale rearing of T. howardi. Under our studied laboratory physical conditions and alternative host, 30 generations of T. howardi were accomplished in ≈600 days, which covers almost two sugarcane cropping cycles, and 12 generations of the sugarcane stalk borer (≈50 days last one generation of Diatraea; Freitas et al., Reference Freitas, Silva, Mendonça, Silva, Fonseca, Mendonça, Santos, Nascimento and Santana2007).
Body size is another common measure of parasitoid quality. In general, larger parasitoid females are expected to have greater longevity and fecundity (West et al., Reference West, Flanagan and Godfray1996), with body size influenced by host size (Silva-Torres et al., Reference Silva-Torres, Ramos Filho, Torres and Barros2009), a common result with eulophid parasitoids (West et al., Reference West, Flanagan and Godfray1996; Silva-Torres and Matthews, Reference Silva-Torres and Matthews2003; Liu et al., Reference Liu, Bauer, Miller, Zhao, Gao, Song, Luan, Jin and Gao2007). Here, we found that tibia length was greater for females of the F5 and F30 generations that emerged from T. molitor compared to those in the F15 generation that emerged from T. molitor and F0 generation that emerged from the D. saccharalis. Similarly, Vargas (Reference Vargas2013) found variation in the cephalic capsule size of T. howardi reared continuously for 25 generations in pupae of D. saccharalis, with this variation observed between generations.
In the same context, although wing size did not differ among the females of the different generations reared in T. molitor, they showed a natural variation in shape, but with similar size and without deformations. In another study, Kölliker-Ott et al. (Reference Kölliker-Ott, Bigler and Hoffmann2004) assessed dispersal and host finding by Trichogramma brassicae Bezdenko (Hymenoptera: Trichogrammatidae) after large-scale rearing over a long period and found that wing size mattered but not wing shape. Furthermore, Ray et al. (Reference Ray, Nakata, Henningsson and Bomphrey2016) showed that modification of Drosophila wing shape by genetic manipulation, which was much greater than natural variation, did not negatively affect the flight ability. Thus, it suggests that the differences in wing shape found in our study are a natural morphological variation not influenced by the rearing host and that, possibly, would not have harmful consequences regarding host location by the parasitoid when released in the field.
Different from pro-ovigenic species, eulophid parasitoids are frequently synovigenic and require feeding to reach egg maturation in the adult stage (Jervis et al., Reference Jervis, Heimpel, Ferns, Harvey and Kidd2001; Zhang et al., Reference Zhang, Liu, Wang, Wan and Li2011). Within 24 h of adulthood, about 50% of the egg load was ready to be laid indicating that egg production increased with parasitoid age. It indicates a gain in ovogenesis through time, a characteristic of synovigenic species. Understanding this parasitoid reproductive strategy will support the recommendation of releasing mated and fed females to improve parasitoid performance in the field.
Females of T. howardi 72 h old reared in D. saccharalis or T. molitor had an average of 72 and 93 developed eggs in the ovary. This knowledge helps to improve the large-scale breeding of this parasitoid using females at an age with maximum reproductive potential. Our findings are in line with those from Zhong et al. (Reference Zhong, Liao, Zhang and Wu2016), who found the first eggs in the ovary of T. howardi 12 h after emergence, while mature eggs were observed only after 24 h, with an increase in egg load up to 72 h (maximum time studied). These authors also found that females produced an average of 71.6 mature eggs. However, females in our study continued producing eggs, reaching maximum egg production at 120 and 144 h when reared with T. molitor and D. saccharalis, respectively. Therefore, females of T. howardi require between 72 and 96 h to complete sexual maturity and egg loading. The findings are consistent with Harvey et al. (Reference Harvey, Poelman and Tanaka2013). According to these authors, the duration to reach sex maturity is an intrinsic trait of species that is based on intraspecific competition and the trade-offs between reproduction and defence, not directly on the natal host.
A similar number of offspring was produced per parasitised pupa of D. saccharalis irrespective of the number of ovipositions on the same pupa. Furthermore, the hypothesis that several ovipositions by T. howardi would be necessary to ensure success in parasitism of the D. saccharalis pupa is ruled out. In fact, one oviposition was enough to kill the sugarcane borer pupa and originate a standard number of descendants. It corroborates with Sequeira and Mackauer (Reference Sequeira and Mackauer1992), who suggested that a female parasitoid must be able to lay the maximum number of eggs in the first oviposition opportunity to ensure the success of offspring production.
Host mortality without offspring production may be related to the significant reduction of eggs in the female ovary after the first oviposition. Females decreased the number of eggs available for additional host parasitism. Most of the eggs were laid at once in the first oviposition, and the recovery time studied seems to not be enough for egg reloading. Therefore, these results support the hypothesis that, after successful parasitism of a sugarcane borer pupa, there will be restrictions on the availability of eggs for a second host attack, at least for a resting period of 24 h. Additionally, we cannot rule out the possibility that a female T. howardi abandoning a parasitised host is not able to kill another host. In fact, pupae were killed with or without the emergence of the parasitoids, a result presumably caused by the female that had already parasitised a host and carried few eggs to address a subsequent oviposition. The death of the host without producing offspring suggests that the number of parasitoid larvae developing in the host was insufficient to overcome the host's immune system. As reported in gregarious species, these species act with cooperative suppression to overcome host resistance (Aya et al., Reference Aya, Montoya-Lerma, Echeverri-Rubiano, Michaud and Vargas2019). Another factor that could cause pupal death has been reported, such as high host sensitivity to microorganism infection (Erb et al., Reference Erb, Bourchier, Van Frankenhuyezen and Smith2001). The parasitism then triggers an immune response, avoiding the emergence of both parasitoid and host.
To overcome the host's defences, eulophid parasitoids that attack large pupae must lay many eggs at once (Andrade et al., Reference Andrade, Serrão, Zanuncio, Zanuncio, Leite and Polanczyk2010; Pereira et al., Reference Pereira, Guedes, Serrão, Zanuncio and Guedes2017). They may also choose to remain with the host (Matthews et al., Reference Matthews, González, Matthews and Deyrup2009). The expectation that the T. howardi female would stay with the sugarcane borer pupa inside the stalk was contradicted, as we found that the female abandoned the parasitised pupa. The data indicate that a single oviposition event of T. howardi on a pupa of D. saccharalis produced several offspring (up to 70 descendants), with only a few eggs remaining in the ovary of the parasitoid. To effectively kill and parasitise the alternative host, T. molitor, more T. howardi females are needed, resulting in more progeny (around 130–140 per pupa on average) than a single female could. This suggests that, in contrast to a D. saccharalis pupa (which has between 40 and 70 offspring), many females lay eggs in a single T. molitor pupa. Therefore, D. saccharalis parasitised pupae are abandoned since they may be killed with only one oviposition and yet generate healthy progeny.
Even though the parasitoid did not remain with the sugarcane borer parasitised pupa inside the stalk, the average number of offspring was 137 parasitoids, with a maximum of 237 parasitoids produced per pupa, a number greater than the average number of descendants produced by a female performing a single oviposition. These findings suggest that under the settings of our bioassay, more than one parasitoid female found the pupa inside the stalk, resulting in superparasitism. This behaviour has been seen in the wild and in the laboratory (Janssen, Reference Janssen1989; Matthews and Deyrup, Reference Matthews and Deyrup2007; Pereira et al., Reference Pereira, Guedes, Serrão, Zanuncio and Guedes2017). Although superparasitism was formerly thought to be an adaptive mistake (Bakker et al., Reference Bakker, Van Alphen, Van Batenburg, Van der Hoeven, Nell, van Strien-van Liempt and Turlings1985), it may enhance natural selection by promoting competition among individuals of the same species. The eulophid P. elaeisis Delvare and LaSalle (Hymenoptera: Eulophidae) had more offspring and greater fitness when it superparasitised on a melonworm pupa (Pereira et al., Reference Pereira, Guedes, Serrão, Zanuncio and Guedes2017).
Currently, Trichogramma galloi Zucchi (Hymenoptera: Trichogrammatidae) and Cotesia flavipes (Cameron) (Hymenoptera: Braconidae) have been released, respectively, against eggs and larval stages of sugarcane stalk borers in Brazil (Parra and Coelho, Reference Parra and Coelho2019). Although there are records of T. howardi parasitising stalk borers' larvae, there is a preference for the pupal stage (Rodrigues et al., Reference Rodrigues, Pereira, Barbosa, Silva-Torres and Torres2021), a clear niche stage preference that will result in increasing pest mortality instead of competition. Therefore, it is worth mentioning that the main goal of T. howardi use is to increase mortality of the sugarcane borer pupae, complementing the control attained during earlier developmental stages by the release of the egg and larval parasitoids.
In conclusion, T. howardi retains its preference and performance over D. saccharalis pupa even after up to 30 generations of rearing using the alternative host, T. molitor. The parasitoid females produced few mature eggs within 24 h of adulthood and only reached maximal egg load between 72 and 96 h, regardless of the natal host, T. molitor or D. saccharalis. Females of T. howardi lay most of their eggs in a single oviposition, and the first oviposition is sufficient for D. saccharalis parasitism. The number of eggs remaining in the ovary after the first oviposition is significantly reduced, and egg load does not recover for 24 h of resting. After the first oviposition, parasitism of T. howardi on a second pupa of D. saccharalis within 24 h will cause the pupa to die; however, this parasitism has a decreased probability of producing progeny. In addition, female T. howardi abandon their host after the parasitism of D. saccharalis pupae within the sugarcane stalk, thus indicating that a single virgin female and one oviposition are sufficient. Alternatively, another female may encounter a pupa that has already been parasitised inside the stalk and proceed to parasitise it as well.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0007485324000129.
Acknowledgements
The authors acknowledge the CAPES Foundation – through Finance Code 001 for providing a doctoral grant to N.C.B. We are also grateful to Fabricio F. Pereira (LECOBIOL/UFGD), Rogério Lira (UFRPE), and Alessandra C. G. Silva (SEE/PE) for suggestions on an early draft of this work and to Gabriella San José (University of California Riverside) for valuable correction of English writing.
Competing interests
None.










