



Este trabajo busca contribuir al entendimiento de la interrelación entre cambio ambiental, modificaciones en las estrategias de subsistencia y movilidad de grupos cazadores-recolectores, y la incorporación de nuevas tecnologías de captura y procesamiento de recursos durante el Holoceno tardío (sensu Walker et al. Reference Walker, Berkelhammer, Björck, Cwynar, Fisher, Long, Lowe, Newnham, Rasmussen and Weiss2012) en el centro-oeste de Santa Cruz (COSC), Argentina. En este contexto se espera una variación en la dieta de estas poblaciones para finales del Holoceno tardío como resultado de modificaciones e innovaciones tecnológicas y, por lo tanto, se plantea que dichas variaciones pueden ser registradas a través del estudio de los isótopos estables en restos humanos. Para ello se utiliza el concepto de nicho isotópico que permite evaluar la amplitud y el solapamiento de los nichos entre grupos humanos previos y posteriores a estos acontecimientos (Jackson et al. Reference Jackson, Inger, Parnell and Bearhop2011; Newsome et al. Reference Newsome, del Rio, Bearhop and Phillips2007).
La investigación se enmarca en un proyecto arqueológico de carácter regional que propone un modelo de poblamiento tardío para el COSC (Goñi Reference Goñi2010; Goñi y Barrientos Reference Goñi, Barrientos, Civalero, Fernández and Guraieb2004). Estudios previos han evidenciado fluctuaciones climáticas durante el Holoceno tardío, que tendió hacia una reducción de la humedad en distintas partes de Patagonia (Moy et al. Reference Moy, Moreno, Dunbar, Kaplan, Francois, Villalba, Haberzettl, Vimeux, Sylvestre and Khodri2009; Stine Reference Stine1994; Stine y Stine Reference Stine and Stine1990; sin embargo, véase Haberzettl et al. Reference Haberzettl, Fey, Lücke, Maidana, Mayr, Ohlendorf, Schäbitz, Schleser, Wille and Zolitschka2005; Ozán et al. Reference Ozán, de Porras, Morales and Barberena2022; Quade y Kaplan Reference Quade and Kaplan2017; Ramos et al. Reference Ramos, Alperin, Schwalb, Markgraf, Ariztegui and Cusminsky2019). Estas nuevas condiciones climáticas se asociaron con una reducción de la movilidad residencial, que derivó en un incremento demográfico como resultado de la concentración de los grupos humanos en las cuencas de altitud baja, tales como los lagos Salitroso-Posadas y Cardiel (Goñi Reference Goñi2010; Goñi et al. Reference Goñi, Re, Guraieb, Cassiodoro, Tessone, Rindel, Dellepiane, Coni, Guichón and Agnolin2019). A su vez, se complementó con un proceso de extensificación a nivel espacial, definido como la introducción de nuevas áreas que previamente no habían sido usadas o que se usaron de manera poco intensiva para la producción y/o extracción de recursos (Binford Reference Binford2001). En este sentido, se postula que se ampliaron los rangos de acción en términos logísticos y estacionales hacia las mesetas y las cuencas altas, como la meseta del Strobel, Pampa del Asador/Guitarra y las cuencas altas del Parque Nacional Perito Moreno (PNPM). Bajo este marco, combinado con la necesidad de mayores requerimientos alimenticios y la predictibilidad conductual de la principal presa de la región, el guanaco (Lama guanicoe), los grupos humanos habrían llegado a una mayor planificación en las actividades relacionadas con la obtención y el procesamiento de recursos. Como resultado, se incorporaron al sistema de la organización social la cerámica, los parapetos, el arco y flecha (Cassiodoro Reference Cassiodoro2011; Flores Coni Reference Flores Coni2019).
Estos cambios en la movilidad y demografía para fines del Holoceno tardío se plasmaron en el registro bioarqueológico. El aspecto más sobresaliente es que el registro mortuorio de la región presenta una alta concentración de entierros en el Lago Salitroso para este período (Figura 1). Hasta la fecha se han analizado 37 estructuras de entierros con presencia de restos humanos, que se diferenciaron en al menos tres modalidades: entierro bajo bloque (EBB) y nichos (ca. 2600-2200 años aP), y chenques. Estos últimos están separados en iniciales (ca. 1600-1200 años aP) y tardíos (ca. 800-350 años aP; García Guraieb et al. Reference García Guraieb, Goñi and Tessone2015; Goñi y Barrientos Reference Goñi, Barrientos, Civalero, Fernández and Guraieb2004; Goñi et al. Reference Goñi, Barrientos and Cassiodoro2000-2002, Reference Goñi, Bosio and Guraieb2003-2005). Los chenques son estructuras de rocas visibles en el espacio y se encuentran en todo Patagonia meridional en los últimos 2.000 años (García Guraieb et al. Reference García Guraieb, Goñi and Tessone2015; Goñi y Barrientos Reference Goñi, Barrientos, Civalero, Fernández and Guraieb2004; Goñi et al. Reference Goñi, Re, Guraieb, Cassiodoro, Tessone, Rindel, Dellepiane, Coni, Guichón and Agnolin2019; Zilio Reference Zilio2013). Los restos humanos del Lago Salitroso han sido analizados desde la paleodemografía y paleopatología (García Guraieb Reference García Guraieb2010; García Guraieb et al. Reference García Guraieb, Goñi and Tessone2015; Guichón Fernández y García Guraieb Reference Guichón Fernández, Guraieb, Fabra, Novellino, Arrieta and Salega2019; Morlesín Reference Morlesín2020); desde los estudios de ADN (Arencibia et al. Reference Arencibia, Crespo, Guraieb, Russo, Dejean and Goñi2019); la tafonomía (Barrientos et al. Reference Barrientos, Goñi, Zangrando, Del Papa, Guraieb, Arregui, Negro, Gutiérrez, Miotti, Barrientos, Goñalons and Salemme2007; Guichón Fernández y García Guraieb Reference Guichón Fernández, Guraieb, Fabra, Novellino, Arrieta and Salega2019), la morfometría y rasgos discretos (Bernal Reference Bernal2008; Pérez et al. Reference Pérez, Beguelin, Del Papa, Civalero, Fernández and Guraieb2004), contextos mortuorios (Cassiodoro Reference Cassiodoro2011; Cassiodoro y García Guraieb Reference Cassiodoro, Guráieb, Salemme, Santiago, Álvarez, Piana, Vázquez and Mansur2009) y la paleodieta (García Guraieb et al. Reference García Guraieb, Goñi and Tessone2015; Tessone et al. Reference Tessone, Guraieb, Goñi and Panarello2015).

Figura 1. Ubicación de la región de estudio en el centro-oeste de Santa Cruz. (Color en la versión electrónica)
Los estudios paleodietarios han señalado que los valores de δ13C y δ15N del colágeno no registran diferencias en los últimos 3000 años aP —es decir, la dieta proteica se mantuvo a través del tiempo. No obstante, la dieta energética o total se modificó, ya que los valores δ13C de la apatita se volvieron más negativos a través del tiempo (Cassiodoro et al. Reference Cassiodoro, Rindel, Goñi, Re, Tessone, Guráieb and Belardi2013). Para entender las continuidades y/o cambios de la dieta cazadora-recolectora, se utiliza el concepto de nicho isotópico (Newsome et al. Reference Newsome, del Rio, Bearhop and Phillips2007). Este término es definido como un espacio multivariado con coordenadas que registran tanto los recursos usados (eje escenopoético), como el escenario sobre el cual se desempeñan los actores (eje bionómico) que se representa mediante los isótopos estables (Newsome et al. Reference Newsome, del Rio, Bearhop and Phillips2007). Como resultado, el nicho isotópico es una poderosa herramienta para investigar y hacer inferencias sobre aspectos claves de la ecología de un individuo o una comunidad (Newsome et al. Reference Newsome, del Rio, Bearhop and Phillips2007).
En este trabajo se presentan valores inéditos de δ13C sobre apatita y valores de δ15N y δ13C sobre colágeno de 42 individuos recuperados en el COSC, los cuales se separaron en grupo temprano (ca. 2600-2200 años aP, EBB y nichos) y tardío (ca. 800-350 años aP, chenques tardíos). El objetivo es corroboran si, efectivamente, estos cambios implicaron modificaciones en el nicho isotópico. Para ello, se utilizaron elipses bayesianas de isótopos estables (SIBER —stable isotope Bayesian ellipse en R, por sus siglas en inglés). Esta herramienta permitirá estimar la amplitud de los nichos y comparar el espacio isotópico utilizado por estos dos grupos, los cuales coinciden con momentos previos y posteriores a la aparición de las nuevas tecnologías y modificaciones socioambientales.
Aunque, anteriormente, se habían abordado las diferencias en la alimentación de acuerdo con las estimaciones etarias de los restos esqueletales del Lago Salitroso (proceso de lactancia y destete; Tessone et al. Reference Tessone, Guraieb, Goñi and Panarello2015), en esta ocasión se abordó la dieta vinculada a la determinación sexual de los individuos. Este análisis se realizó sobre el grupo tardío en la medida que existe una distribución equilibrada de individuos de ambos sexos. Distintas investigaciones etnográficas y arqueológicas han señalado una división sexual del trabajo entre los grupos extractivos, generalmente asociada al hombre cazador y la mujer recolectora, lo cual implicó, en algunas ocasiones, diferencias en la dieta (Aguerre Reference Aguerre2000; Berbesque y Marlowe Reference Berbesque and Marlowe2009; sin embargo, véase Haas et al. Reference Haas, Watson, Buonasera, Southon, Chen, Noe, Smith, Llave, Eerkens and Parker2020). En el caso de la Patagonia esteparia, la focalización en la caza y consumo del guanaco, que es una fuente de proteína magra, sumada a la baja diversidad en fauna y flora disponible en el ambiente, pudieron haber generado requerimientos dietarios especiales —particularmente en las mujeres, quienes habrían necesitado garantizar el consumo de grasa para el mantenimiento de la fertilidad y, además, como fuente de energía para el embarazo y la lactancia (Frisch Reference Frisch1994). De esta manera, a través del SIBER, se buscó evaluar si hubo diferencias temporales y sexuales entre los nichos isotópicos de los individuos enterrados en chenques tardíos, que no sólo presentaron una nueva forma de entierro, sino que también tenían patrones diferentes de organización social y económica en relación a los momentos previos a los 900 años aP en Patagonia meridional.
Ambiente del área de estudio
El COSC, Patagonia Argentina (Figura 1), se caracteriza por ser un ambiente frío con marcada estacionalidad y precipitaciones anuales menores a 200 mm (Oliva et al. Reference Oliva, González, Rial, Livraghi, Borrelli and Oliva2001). Estas condiciones generan desafíos a la supervivencia debido a la baja diversidad de recursos vegetales y animales (Goñi Reference Goñi2010). La comunidad vegetal local consiste en una estepa herbácea y de arbustos bajos. Entre las plantas consumibles se hallan las que tienen órganos de almacenamiento subterráneos comestibles (Azorella, Oxalis, Bolax spp.), que están ampliamente distribuidas por Patagonia y son reservorio de agua, carbohidratos y proteínas (Ochoa y Ladio Reference Ochoa and Ladio2011, Reference Ochoa and Ladio2015). Otros recursos vegetales consumibles son frutos, como los calafates (Berberis sp.), molles (Schinus sp.), murtilla (Empetrum rubrum), entre otros (Ochoa y Ladio Reference Ochoa and Ladio2011; Rapoport y Ladio Reference Rapoport and Ladio1999). En cuanto a la fauna, el choique (Pterocnemia pennata) y el guanaco (Lama guanicoe) son las especies animales más accesibles y ubicuas (Bonino Reference Bonino2005). Este último fue la presa principal de los cazadores-recolectores durante el Holoceno y se caracteriza por ser una excelente fuente de proteínas magras.
De acuerdo con la información paleoambiental de la región, existieron cambios climáticos durante el Holoceno. La dirección e intensidad de los vientos del oeste influyen fuertemente en el clima patagónico, principalmente en las precipitaciones (Gilli et al. Reference Gilli, Anselmetti, Ariztegui, Platt Bradbury, Kelts, Markgraf and McKenzie2001). Hace 2.500 años, los vientos del oeste cambiaron y provocaron fluctuaciones en la distribución de los recursos hídricos producto de una reducción de la humedad a una escala regional (Ariztegui et al. Reference Ariztegui, Gilli, Anselmetti, Markgraf, Goñi, Belardi, Cassiodoro and Re2014; Gilli et al. Reference Gilli, Ariztegui, Anselmetti, McKenzie, Markgraf, Hajdas and McCulloch2005; Horta et al. Reference Horta, Georgieff, Aschero and Goñi2017; Stine y Stine Reference Stine and Stine1990). Investigaciones en el Lago Viedma señalan que para estas fechas el nivel del lago bajó hasta los 266 m snm (Horta et al. Reference Horta, Belardi, Georgieff and Carballo2022). Esto habría habilitado nuevos espacios de circulación para las poblaciones del pasado, aunque hay registros que difieren de esta propuesta (Haberzettl et al. Reference Haberzettl, Fey, Lücke, Maidana, Mayr, Ohlendorf, Schäbitz, Schleser, Wille and Zolitschka2005; Ozán et al. Reference Ozán, de Porras, Morales and Barberena2022; Quade y Kaplan Reference Quade and Kaplan2017; Ramos et al. Reference Ramos, Alperin, Schwalb, Markgraf, Ariztegui and Cusminsky2019). En cuanto a la vegetación, información palinológica del Lago Los Flamencos señala el establecimiento de una estepa arbustiva en el área del Lago Pueyrredón, similar a la actual a partir de los últimos 3500 años aP (Marcos et al. Reference Marcos, Bamonte, Echeverría, Sottile and Mancini2021).
Antecedentes tecnológicos
Durante el Holoceno tardío, las nuevas condiciones ambientales regionales, los cambios en las estrategias de movilidad y una nueva configuración demográfica recrudecieron la necesidad de mejorar el aprovechamiento de la poca grasa del guanaco. Como resultado, estos hechos habrían conllevado cambios en el aspecto tecnológico de las sociedades (Cassiodoro Reference Cassiodoro2011). Por un lado, los cambios en las estrategias tecnológicas vinculadas con la obtención de los recursos estarían relacionados con el desarrollo de técnicas de caza planificadas y la incorporación de nuevas modalidades de propulsión de proyectiles. Así, se habrían implementado técnicas por interceptación y/o acecho en determinados sectores. Las mesetas fueron sistemáticamente equipadas con estructuras de piedra semicirculares, denominadas parapetos, que habrían funcionado como apostaderos de caza en puntos específicos del paisaje (Cassiodoro Reference Cassiodoro2011; Flores Coni Reference Flores Coni2019; Gradin Reference Gradin1959-1960). En el análisis de las cronologías calibradas de las estructuras de parapetos y de las de entierros humanos, resulta relevante destacar cómo los fechados se agrupan de igual manera. En ambos casos, se observa una utilización inicial hacia los 2500-2000 años aP, una escasa representación entre los 1500 y 1000 años aP, y una mayor frecuencia en momentos posteriores a los 900 aP (Flores Coni Reference Flores Coni2019). Asimismo, para los momentos tardíos del Holoceno se registra la incorporación de una tecnología eficiente y confiable para la obtención de presas como es la implementación del arco y flecha (Cassiodoro Reference Cassiodoro2011).
Por otro lado, los cambios en las estrategias tecnológicas vinculadas con el procesamiento de recursos estarían relacionados con la necesidad de contar con un reaseguro contra la escasez en la estación del año más pobre de la región. En determinados momentos del año, los recursos animales tienden a ofrecer menor cantidad de grasas a las poblaciones humanas de Patagonia, de ahí que estas se vean ante la necesidad de optimizar su uso. En condiciones ambientales más secas, estos procesos se podrían haber intensificado. En este sentido, la cerámica pudo haber sido utilizada para facilitar el derretimiento de las grasas animales y, a su vez, para su almacenamiento y posterior consumo (Cassiodoro Reference Cassiodoro, Austral and Tamagnini2008, Reference Cassiodoro2011; Chaile et al. Reference Chaile, Lantos, Maier, Cassiodoro and Tessone2018a; Goñi Reference Goñi2010). Considerando que la incorporación de cerámica mejora el aprovechamiento de proteínas, remueve patógenos y amplía la vida útil de los alimentos (Wandsnider Reference Wandsnider1997), su utilización en Patagonia a partir de determinado momento del Holoceno tardío debe ser entendida en el marco de nuevas estrategias de subsistencia. En la región de estudio la tecnología cerámica comienza a aparecer en el registro arqueológico alrededor de los 1200 años aP (Gradin y Aguerre Reference Gradin and Aguerre1991), aunque se registra con mayor frecuencia en momentos posteriores a los 900 aP (Cassiodoro y Tessone Reference Cassiodoro and Tessone2014; Chaile et al. Reference Chaile, Goñi, Cassiodoro, Schuster and Pérez2020). La cuenca baja del Lago Salitroso es el sector de la región con mayor cantidad de sitios arqueológicos con cerámica (Cassiodoro Reference Cassiodoro2011; Goñi et al. Reference Goñi, Re, Guraieb, Cassiodoro, Tessone, Rindel, Dellepiane, Coni, Guichón and Agnolin2019).
Igualmente, la tecnología de molienda puede constituir un indicador del desarrollo de estrategias que busquen una mayor optimización de los recursos. Fundamentalmente, esta tecnología tiene por objeto resolver problemas vinculados con la alteración de sustancias mediante procesos mecánicos o químicos que tienen lugar cuando entran en contacto dos superficies (Babot Reference Babot2004). La presencia de artefactos de molienda en alta frecuencia en momentos posteriores a los 1600-1500 años aP no sólo constituye, en los contextos analizados, un indicador del equipamiento de los espacios (Cassiodoro Reference Cassiodoro, por Alejandra, Re, Rindel and Valeri2005, Reference Cassiodoro2011), sino que también es vinculable con cambios en el procesamiento de recursos (Cassiodoro Reference Cassiodoro2011). Por ejemplo, en otras áreas de la Patagonia, hay información arqueológica e histórica de su uso para la molienda de algarroba (Prosopis sp.; Capparelli y Prates Reference Capparelli and Prates2015; Ciampagna y Capparelli Reference Ciampagna and Capparelli2012; Prates Reference Prates2009); también se menciona la recolección de rizomas y tubérculos que eran molidos para obtener la harina, y luego guardarla en ollas (Pigafetta Reference Pigafetta2001; Vignati Reference Vignati1941). A su vez, de manera complementaria, podrían estar funcionando para la fractura y machacado de huesos (Bourlot Reference Bourlot2009). Ambas tecnologías podrían constituir un tool kit específico de procesamiento y, probablemente, almacenamiento de recursos necesarios para su mayor aprovechamiento. De esta manera, la tecnología se entiende como parte de una estrategia mayor de resolución de problemas relacionados con el medio físico y social (Binford Reference Binford1979; Torrence Reference Torrence1989), que habría posibilitado una modificación en la dieta de las personas durante el último tramo del Holoceno tardío en el interior de Patagonia continental.
Nicho isotópico y SIBER
Los análisis de isótopos estables han permitido discutir diversas temáticas tales como la paleodieta y la movilidad de las sociedades humanas en el pasado, como también la caracterización del paleoambiente que habitaron (Ambrose Reference Ambrose and Sandford1993; Fernández y Panarello Reference Fernández and Panarello1991; Tieszen Reference Tieszen1991). Para ello, se han estudiado diferentes tejidos y materiales según las preguntas y objetivos de los investigadores. En el caso de la paleodieta en arqueología, el tejido más frecuentemente usado es el hueso por ser el de mayor perdurabilidad en el registro, el cual presenta distintas fracciones (Panarello et al. Reference Panarello, Tessone and Zangrando2010, Reference Panarello, Tessone, Killian Galván, Samec, Kochi, Pirola and Chaile2021). Por un lado, se encuentra la fracción orgánica denominada colágeno. En esta fracción, el 75% del carbono es contribuido por la proteína, mientras que el 25% restante es aportado por los lípidos y carbohidratos (Fernandes et al. Reference Fernandes, Nadeau and Grootes2012). Por otro lado, están los valores de δ13C de la fracción inorgánica o apatita que se compone por el aporte similar de los tres macronutrientes: proteínas, lípidos y carbohidratos, reflejando como resultado el valor de la dieta total. En el caso del nitrógeno, que proviene de la proteína, permite diferenciar el posicionamiento trófico del consumidor y se mide a partir del colágeno óseo (Ambrose Reference Ambrose and Sandford1993).
Los análisis de isótopos estables son una herramienta que permite, a través del nicho isotópico, responder cuáles fueron los recursos que usaron los individuos, así como cuál fue el hábitat en el cual estos vivieron (Newsome et al. Reference Newsome, del Rio, Bearhop and Phillips2007). Pero para ello, es necesario determinar la amplitud del nicho isotópico y el grado de superposición de los nichos entre los grupos humanos. Por este motivo, se utilizó el programa SIBER 2.1.4, que permite analizar datos de isótopos estables y generar métricas de las dimensiones de los nichos isotópicos.
El área total del polígono (TA —total area of the convex hull) representa el área total ocupada por la especie, comunidad y/o grupo. Este se dibuja alrededor de los puntos más externos en la nube de datos, de modo que todos los demás puntos se encuentran dentro del contorno. Por lo tanto, si recogiésemos más muestras, este polígono sólo se mantendría o crecería en tamaño, pero no se reduciría. El resultado es que el tamaño de la muestra influye en los polígonos. A pesar de este problema estadístico, el área total del polígono sigue siendo una forma útil de ayudarnos a visualizar datos bivariados, como los valores de isótopos estables de carbono-nitrógeno (Jackson et al. Reference Jackson, Inger, Parnell and Bearhop2011). Por el contrario, el área de la elipse estándar corregida (SEAC) no está afectada por el tamaño de la muestra o por datos extremos, y se corresponde con el 40% de los datos de la muestra (Jackson et al. Reference Jackson, Inger, Parnell and Bearhop2011). Se utiliza en este trabajo para calcular el área de solapamiento entre elipses. Por último, para comparar la amplitud del nicho entre grupos, se genera una estimación bayesiana de la elipse estándar y su área (SEAB). El enfoque bayesiano devuelve una distribución posterior de SEA que refleja la incertidumbre que surge del proceso de muestreo. Las elipses resultantes se pueden comparar cuantitativamente y las diferencias se establecen con referencia a una probabilidad robusta (Jackson et al. Reference Jackson, Inger, Parnell and Bearhop2011).
Materiales y métodos
En este trabajo se presentan valores inéditos de δ13C sobre biopatita de 42 individuos recuperados en el COSC, acompañados de sus valores de δ13C y δ15N sobre colágeno. Los individuos se distribuyen espacialmente de la siguiente manera: 39 de la cuenca del Lago del Salitroso, uno en el valle del Río Olnie (curso inferior), uno en la cuenca del Lago Pasadas/Pueyrredón y uno en el Parque Nacional Perito Moreno (Figura 1). Asimismo, sólo se consideraron aquellos individuos con estimaciones etarias correspondientes a las categorías de adolescentes y de adultos de ambos sexos. En el caso de la dieta de los subadultos del Lago Salitroso fue analizada en un trabajo previo (Tessone et al. Reference Tessone, Guraieb, Goñi and Panarello2015).
La muestra se separó de la siguiente manera: (1) 11 individuos provenientes de EBB y nichos, y (2) 31 individuos hallados en chenques tardíos. Los restos humanos de los chenques iniciales no fueron analizados debido a la escasez de la muestra para este período. Para responder a la problemática de si hubo diferencias en la dieta en función de las determinaciones sexuales, se analizó un total de 21 individuos enterrados en chenques tardíos, 10 de los cuales fueron identificados como masculinos y 11 como femeninos (García Guraieb et al Reference García Guraieb, Goñi and Tessone2015; Tessone et al. Reference Tessone, Guraieb, Goñi and Panarello2015).
Se priorizaron las costillas para la extracción del colágeno y la apatita. Se realizó una limpieza mecánica de la muestra y enjuague en baño de ultrasonido con agua desionizada. Para la extracción del colágeno se desmineralizó el hueso en ácido clorhídrico (HCl 2%) durante 72 horas, reemplazando el ácido cada 24 horas. Para eliminar los compuestos postdepositacionales, el material se colocó 24 horas en NaOH antes y después de la desmineralización ósea. Por último, la muestra se enjuagó y secó en una estufa a <60°C (Tykot Reference Tykot, Martini, Milazzo and Piacentini2004). Para la extracción de la apatita, se trató la muestra con hipoclorito de sodio al 2% (NaClO) durante 72 horas para disolver la materia orgánica (Tykot Reference Tykot, Martini, Milazzo and Piacentini2004). Los carbonatos postdepositacionales o secundarios se eliminaron con un buffer ácido acético-acetato por cuatro horas, siguiendo las recomendaciones de Garvie-Lok y colaboradores (Reference Garvie-Lok, Varney and Katzenberg2004). Finalmente, se enjuagó de nuevo en agua desionizada y se secó en horno antes de su pesado final.
Estos análisis se realizaron en el Instituto de Geocronología y Geología Isotópica (INGEIS-UBA, CONICET). La medición de 13C/12C y 15N/14N en la fracción de colágeno se realizó con un analizador elemental (CHN) Carlo Erba EA 1108, conectado a un espectómetro de masa Thermo Scientific Delta V Advantage de flujo continuo a través de una interfaz Thermo Scientific ConFlo IV. El análisis de carbono-13 en apatita se realizó en un IRMS Finnigan MAT Delta S de doble entrada. Los resultados de isótopos estables se expresan como la relación entre el isótopo más pesado y el más liviano (13C/12C o 15N/14N) y se informan como valores δ en partes “por mil” (‰) en relación con los estándares de carbono (Vienna Pee Dee Belemnite, VPDB) y nitrógeno (Ambient Inhalable Reservoir, AIR) definidos internacionalmente.
En términos estadísticos para saber si las diferencias entre los grupos estudiados son significativas, se realizó previamente la prueba estadística de normalidad de Shapiro-Wilk. A los grupos que presentaron una distribución normal se les aplicó la prueba estadística paramétrica test-T para muestras independientes, mientras que se aplicó el test no paramétrico Mann-Whitney a los grupos que no presentaron una distribución normal. Además, se realizó la prueba de Levene para evaluar la igualdad de las varianzas.
Resultados
Los valores obtenidos de δ13C en colágeno y apatita y δ15N en colágeno junto a los fechados publicados en trabajos anteriores, se observan en la Tabla suplementaria 1. Respecto a las condiciones de conservación, contaminación y diagénesis, la relación C/N en colágeno se encuentra dentro del rango normal de 2,9-3,6 con una media de 3,2 ± 0,08% (Tabla suplementaria 1; Ambrose Reference Ambrose1990). Respecto a la apatita, si bien no se dispone de parámetros similares a los del colágeno, las características de la muestra —cronología reciente y buen estado general— y del ambiente —condiciones de aridez y frío— (Ambrose Reference Ambrose and Sandford1993), nos hace suponer que los valores obtenidos en apatita son los primarios. De esta manera, los valores de los tres marcadores isotópicos en los 42 individuos son considerados para el ulterior análisis (Tabla suplementaria 1).
La estadística descriptiva de δ13C y δ15N del colágeno (δ13Ccol y δ15N) y de la apatita (δ13Capa) de los dos grupos temporales se presenta en la Tabla 1. Además, se grafica la ecología isotópica del área de estudio con la media de los principales recursos animales disponibles, y la media de la dieta de los dos grupos cronológicos (Figura 2). En las Figuras 3 y 4 se puede apreciar la distribución de los valores en función de los grupos temporales a partir del procesamiento inicial con SIBER. La media del δ13Capa del Grupo 1 (el grupo más temprano), fue de −12,1‰ ± 1,2‰ en tanto que la del Grupo 2 (o el más tardío), fue de −13,5‰ ± 0,7‰. En cuanto a la media del δ15N del primer grupo, fue de 11,6‰ ± 0,7‰, y la del segundo fue de 11,4‰ ± 0,7‰. Por último, los valores medios del δ13Ccol del Grupo 1 fueron de −18,5‰ ± 0,3‰, mientras que los del Grupo 2 fueron de −18,8‰ ± 0,5‰ (Tabla 1).

Figura 2. Ecología isotópica del centro-oeste de Santa Cruz junto a los valores promedios de δ13C y δ15N de las dietas de ambos grupos cronológicos. (Color en la versión electrónica)

Figura 3. Representación del nicho isotópico (valores de δ13C colágeno y δ15N) a través de elipses bayesianas estándar (línea sólida, SEAC) y sus respectivos polígonos (líneas discontinuas, TA) entre ambos períodos cronológicos. (Color en la versión electrónica)

Figura 4. Representación del nicho isotópico (valores de δ13C apatita y δ15N) a través de elipses bayesianas estándar (línea sólida, SEAC) y sus respectivos polígonos (líneas discontinuas, TA) entre ambos períodos cronológicos. (Color en la versión electrónica)
Tabla 1. Estadística comparativa de los distintos marcadores de acuerdo con el grupo temporal.
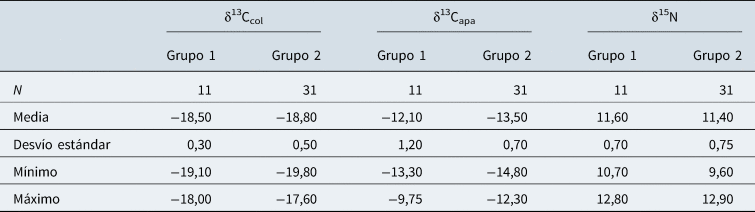
En primer lugar, se evaluó la homocedasticidad de las muestras. Los resultados fueron de p > 0,05; por consiguiente, no hubo diferencias significativas. También se les realizó una prueba de normalidad a los distintos valores de los marcadores isotópicos, los cuales mostraron una distribución normal (Shapiro-Wilk p >0,05), salvo los valores de δ13Capa del conjunto temprano (Shapiro-Wilk p = 0,04). Posteriormente, se realizaron pruebas de significancia para comprobar si existía una diferencia entre los valores isotópicos de los grupos humanos. Los resultados indicaron que el δ15N de la dieta proteica se mantuvo sin cambios significativos (Test-t p = 0,4) a lo largo del tiempo; lo mismo se observó en los valores de δ13Ccol (Test-t p = 0,08). No obstante, los valores de δ13Capa mostraron una diferencia significativa (Mann Whitney p = 0,0002) cuando se compararon los individuos recuperados de EBB y nichos (Grupo 1) con los enterrados en los chenques tardíos (Grupo 2). Estas similitudes y diferencias entre los grupos temporales se observan en las gráficas de dispersión (Figuras 3 y 4). En base a las diferencias halladas en los valores del carbono de la apatita entre ambos segmentos temporales, se compararon las elipses bayesianas y polígonos formados a partir de los valores de la composición isotópica de carbono de la apatita y colágeno (Figura 5). Los resultados de SIBER señalaron que el grupo de EBB y nichos obtuvo un área total del polígono de 1,92%2, un área de elipse estándar de 1,14%2 y su corrección de 1,27%2 mientras que el valor bayesiano fue de 1,26%2. En el caso de los chenques fue un TA de 3,78%2, SEA de 1,27%2, SEAC de 1,31%2 y SEAB de 1,31%2.

Figura 5. Representación del nicho isotópico (valores de δ13C apatita y δ13C colágeno) a través de elipses bayesianas estándar (línea sólida, SEAC) y sus respectivos polígonos (líneas discontinuas, TA) entre ambos períodos cronológicos. (Color en la versión electrónica)
Por otro lado, la Tabla suplementaria 2 presenta la estadística descriptiva de los valores de δ13C de la apatita y del colágeno del hueso según el sexo para los entierros en chenques tardíos. El grupo de los femeninos tiene un valor medio del δ13Ccol de −18,8‰ ± 0,4‰, mientras que su mínimo fue de −19,4‰ y su máximo de −18,2‰. En cuanto a los masculinos, su media fue de −18,9‰ ± 0,6‰ y el rango de valores fue de un mínimo de −19,8‰ a un máximo de −18,0 ‰. En el caso de los resultados del δ13Capa, las mujeres obtuvieron una media de −14,0‰ ± 0,5‰, un mínimo de −14,8‰ y un máximo de −13,0‰. Mientras que los resultados δ13Capa de los hombres presentaron un rango de valores entre −14,4‰ y −12,3‰ y una media de −13,2‰ ± 0,7‰. Además, como en el caso de las comparaciones temporales, se realizaron los test estadísticos para comprobar si existían diferencias significativas entre las dietas según la determinación sexual de los individuos enterrados en chenques tardíos. Las muestras presentaron una homeosticidad p > 0,05 y una distribución normal (p > 0,05). Los resultados indicaron que el valor en la composición isotópica del nitrógeno (Test-t p = 0,90) no presentó diferencias significativas con los del carbono del colágeno (Test-t p = 0,85). Por el contrario, los valores de δ13Capa mostraron diferencias significativas (Test-t p = 0,005) cuando se compararon los individuos masculinos con los femeninos del Grupo 2. En cuanto a los resultados de SIBER, se analizaron las elipses bayesianas y polígonos según el sexo, tomando como datos los valores de carbono de la apatita y el colágeno (Figura 6). Los valores obtenidos fueron, en el caso de las mujeres, de un TA de 1,38%2, un SEA 0,67%2, una SEAC 0,75%2 y un SEAB 0,74%2. Para el caso de los hombres, el área total del polígono fue de 2,18%2, un área de elipsis estándar de 1,27%2 y su corrección de 1,43%2 y, finalmente, un valor bayesiano de 1,42%2.

Figura 6. Representación del nicho isotópico (valores de δ13C apatita y δ13C colágeno) a través de elipses bayesianas estándar (línea sólida, SEAC) y sus respectivos polígonos (líneas discontinuas, TA) entre ambos sexos recuperados de entierros en chenques tardíos (CT). (Color en la versión electrónica)
Discusión y conclusiones
El conjunto de datos presentados en este estudio permite profundizar acerca de las continuidades y cambios en la subsistencia de las poblaciones de Patagonia meridional en los últimos 3.000 años. La información paleodietaria presentada y analizada por SIBER apoyaría la hipótesis de una modificación en el nicho isotópico a través del tiempo. Estos cambios son evidentes en el δ13Capa, que sintetizan y metabolizan la dieta total, conformada por el carbono de los carbohidratos, proteínas y lípidos. En principio, los valores de δ13Capa se empobrecieron en 13C con el tiempo, con una diferencia de 1,4‰ entre las medias de ambos grupos temporales y con una diferencia estadísticamente significativa (Mann-Whitney p <0,05). Trabajos anteriores exploraron las variaciones isotópicas de los últimos 5.000 años en los guanacos de la región de los lagos Tar y San Martin (Tessone y Belardi Reference Tessone, Belardi, Gutiérrez, Nigris, Fernández, Giardina, Gil, Izeta, Neme and Yacobaccio2010). Los autores concluyeron que no hubo un cambio en los valores de δ13C ni de δ15N en el colágeno de los animales a lo largo del tiempo —es decir, no cambió la línea de base trófica (Tessone y Belardi Reference Tessone, Belardi, Gutiérrez, Nigris, Fernández, Giardina, Gil, Izeta, Neme and Yacobaccio2010). Esto llevaría a pensar que la respuesta estaría en un cambio en el tipo de recurso o un mayor énfasis en el consumo de lípidos o carbohidratos durante los últimos 900 años. A partir de estas diferencias registradas se estimó que la amplitud del nicho isotópico del Grupo 1 fue de 1,27‰2, mientras que la de los individuos enterrados en chenques tardíos fue de 1,31‰2. Es decir, no se hallaron grandes diferencias entre el tamaño de las elipses del Grupo 1 y la del Grupo 2 (p = 0,60). No obstante, el solapamiento entre las elipses es pequeño, no supera el 20% si consideramos la SEAC (Tabla 2 y Figura 4). Estos resultados reforzarían la idea de que los restos recuperados en chenques explotaron un nicho isotópico diferente a los individuos EBB y nichos.
Tabla 2. Solapamiento de las áreas de las elipses estándar corregidas por el tamaño de pequeñas muestras (SEAC) para valores isotópicos de distintos grupos de entierros según su cronología.
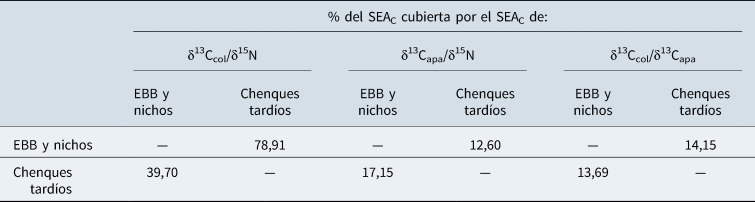
Asimismo, creemos que estas modificaciones se ven reflejadas en los valores de δ13C y δ15N obtenidos en el colágeno. Si bien la dieta proteica se mantuvo constante en los últimos 3.000 años, la amplitud del nicho isotópico de los individuos más tardíos es mayor que la de los más tempranos (Figura 3). Esto se observa al registrar que el 78,91% del área de la elipse estándar (SEAC) del colágeno del Grupo 1 fue solapada por el SEAC de los chenques tardíos, mientras que sólo el 39,70% del SEAC del Grupo 1 solapó con el área de la elipse estándar del Grupo 2. Estas diferencias podrían explicarse porque un 25% del carbono utilizado para la síntesis de colágeno proviene de los carbohidratos/lípidos, que en general son más negativos que la composición isotópica de carbono de las proteínas (Fernandes Reference Fernandes2016). De esta manera, al considerarse que la dieta total fue modificada, son esperables ciertos cambios en los valores δ13C de colágeno. Este es un contrapunto interesante con otras poblaciones de cazadores-recolectores de Patagonia donde se han registrado cambios temporales en el consumo de proteínas, con proporciones diferenciales de proteínas terrestres y marinas (Favier Dubois et al. Reference Dubois, Cristian, Borella, Tykot, Salemme, Santiago, Álvarez, Piana, Vázquez and Mansur2009; Flensborg et al. Reference Flensborg, Martínez and Tessone2020; Zilio et al. Reference Zilio, Tessone and Hammond2018). Por el contrario, en el caso del Lago Salitroso —cazadores-recolectores del interior de Patagonia continental— los cambios en la dieta se observaron en la dieta total, a partir del consumo diferencial de lípidos y/o carbohidratos.
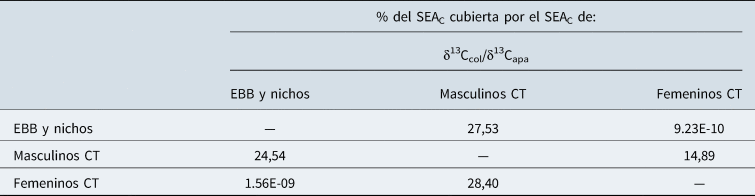
Los análisis de isótopos estables y la aplicación de SIBER en el grupo tardío permitieron discutir la dieta con relación a la determinación sexual de los individuos. Dicho análisis señala que los femeninos de COSC presentaron una dieta significativamente más empobrecida en 13C en la apatita en comparación con los individuos masculinos (Test-t p = 0,005); mientras que, igual que con la cronología, la dieta proteica no mostró diferencias significativas (Figura 6). Los análisis de SIBER, como se mencionó más arriba, señalaron que los valores de SEAC de los masculinos fueron de 1,43‰2, mientras que los de los femeninos de 0,75‰2. Las probabilidades de que la elipse de los masculinos sea más pequeña que la de los femeninos es del 7,75%. Esto significaría que es altamente probable que la amplitud del nicho isotópico y, por ende, la amplitud de la dieta de las mujeres haya sido menor que la de los hombres. En cuanto al porcentaje del área de la elipse estándar de los individuos femeninos cubierta por el SEAC de los masculinos, fue de 28,40% (Tabla 3). En tanto que, a la inversa, el SEAC de los hombres fue cubierta en un 14,89%. De esta manera, los individuos masculinos presentan una mayor diversidad en la dieta en comparación con los femeninos y, asimismo, registran un bajo solapamiento en el nicho isotópico de ambos grupos en los individuos recuperados de los chenques tardíos. Vinculado a este tema, Berbesque y colaboradores (Reference Berbesque, Wood, Crittenden, Mabulla and Marlowe2016) señalan que la mayor parte de la dieta de los hombres Hadza proviene de lo que comen afuera de los campamentos mientras cazan. Por lo tanto, su dieta puede diferir de la de las mujeres ya que incorporan más variedad y cantidad de alimentos que no comparten con sus parejas. Por último, cabe resaltar que el nicho isotópico de las mujeres no se solapó con el de los individuos pertenecientes al grupo de EBB y nichos, a diferencia de los hombres que se superpuso en un 24,54% (Tabla 3). Esto permite señalar que tanto los individuos masculinos como femeninos del grupo tardío presentan una misma tendencia hacia el empobrecimiento del δ13Capa si lo comparamos con el grupo de EBB y nichos. No obstante, los cambios en la dieta fueron mayores en los individuos femeninos que en los masculinos. Una posible explicación es que su dieta contó con un mayor consumo de carbohidrato vegetal, que presenta valores bajos en δ13C. Los estudios etnográficos han vinculado, generalmente, a los artefactos asociados a la explotación de recursos vegetales con la esfera femenina (Berbesque y Marlowe Reference Marlowe and Berbesque2009; Twiss Reference Twiss2012). Otra posibilidad es que las mujeres hayan consumido más cantidad de grasa como requerimiento dietario; dado que su cuerpo necesita mayores proporciones de grasa corporal que los hombres (Berbesque y Marlowe Reference Marlowe and Berbesque2009). Esto se debe a que la grasa tiene la función de proveer energía durante el embarazo y hasta los tres meses de lactancia (Berbesque y Marlowe Reference Marlowe and Berbesque2009; García Guraieb Reference García Guraieb2010; Marlowe y Berbesque Reference Marlowe and Berbesque2009), por lo que sería necesario su almacenamiento para aquellos momentos de mayor requerimiento. Cabe resaltar que ambas propuestas no son excluyentes.
Tabla 3. Solapamiento de las áreas de las elipses estándar corregidas por el tamaño de pequeñas muestras (SEAC) para valores isotópicos de individuos hallados en chenques tardíos según sexo y EBB/nichos.
De esta manera, se observó un empobrecimiento en los valores δ13Capa a través del tiempo al comparar ambos grupos temporales, y esta tendencia es más notoria en los individuos femeninos del grupo tardío. Esto se ajustaría a la idea de un cambio en las estrategias de subsistencia y en la dieta de estos grupos cazadores-recolectores del Holoceno tardío. Estos cambios creemos que se basan, principalmente, en un mayor aprovechamiento y consumo de grasas óseas de guanacos, aunque no se puede descartar un aumento del consumo de vegetales. Respecto a la primera de las posibilidades, creemos que esta actividad ocupó un espacio importante en la dieta de los grupos cazadores-recolectores de Patagonia meridional; en especial para aquellos cuya subsistencia dependió en gran medida de recursos alimenticios altos en proteínas terrestres. Dado que la diversidad ecológica de recursos explotables en el interior de la estepa patagónica es baja, la subsistencia de los grupos humanos se solventó fundamentalmente a través del aprovechamiento integral del guanaco (Cassiodoro Reference Cassiodoro2011; Goñi Reference Goñi2010). Una posible solución ante tales problemas, como ya se mencionó, fue la producción y el almacenamiento planificado y sistemático de grasa animal en los sectores altos (meseta de Pampa del Asador/Guitarra, meseta del Strobel) para su posterior transporte y uso en los sectores bajos durante el invierno (cuenca del Lago Salitroso, Cardiel; Cassiodoro Reference Cassiodoro2011; Chaile et al. Reference Chaile, Lantos, Maier, Cassiodoro and Tessone2018a; Goñi Reference Goñi2010). Para este período se registra un procesamiento estandarizado de las carcasas de guanaco por medio de la fractura transversal/marcado perimetral (Bourlot et al. Reference Bourlot, Rindel, Aragone, Salemme, Santiago, Álvarez, Piana, Vázquez, Mansur and tomo2009). Dicha fractura estaría vinculada con tecnologías como la cerámica y los artefactos de molienda que habrían ayudado a la extracción completa de la médula ósea y grasa trabecular (Bourlot et al. Reference Bourlot, Rindel, Aragone, Salemme, Santiago, Álvarez, Piana, Vázquez, Mansur and tomo2009; Cassiodoro Reference Cassiodoro2011; Cassiodoro y Tessone Reference Cassiodoro and Tessone2014).
Nuestros trabajos previos de identificación de la estructura química de los componentes de los residuos orgánicos presentes en los contenedores cerámicos han sido coherentes con la hipótesis propuesta en esta investigación (Chaile et al. Reference Chaile, Lantos, Maier, Cassiodoro and Tessone2018a, Reference Chaile, Goñi, Cassiodoro, Schuster and Pérez2020). Por un lado, evidencia de lo mencionado, es la detección de esteroles biomarcadores de origen animal (colesterol) y ácidos grasos de carbonos impares lineales y ramificados en residuos absorbidos en los tiestos cerámicos, característicos de animales rumiantes (Chaile et al. Reference Chaile, Lantos, Maier, Cassiodoro and Tessone2018a). Por otro lado, los valores bajos en δ13C de los residuos carbonizados fueron asociados a la abundancia de lípidos y/o carbohidratos (Chaile et al. Reference Chaile, Tessone, Cassiodoro, Bellelli and Belardi2018b).
En suma, en el centro-oeste de Santa Cruz hacia fines del Holoceno tardío hubo una reducción de la humedad que generó cambios en las estrategias de movilidad, nuevas configuraciones demográficas e incorporaciones y modificaciones tecnológicas vinculadas a la captura y procesamiento de recursos. El objetivo de este trabajo fue evaluar si estos cambios se reflejaron en una variación en el nicho isotópico de estos grupos cazadores-recolectores o, por el contrario, si hubo una continuidad en su explotación durante los últimos 3.000 años. Para ello, se presentó información isotópica δ13Capa sobre restos humanos junto a su análisis con SIBER. Los resultados señalaron que hubo un cambio en su dieta en los últimos 1.000 años que se plasmó en valores más bajos en δ13Capa de los individuos enterrados en los chenques tardíos y, en particular, en las mujeres. Especialmente, se trataría de un mayor aprovechamiento de la grasa, probablemente ósea, en los últimos 1.000 años. Sin embargo, no se descarta que parte de los cambios también refieran a una modificación en el consumo de los recursos vegetales. En cuanto a las diferencias dietarias halladas entre los individuos enterrados en chenques tardíos de acuerdo con el sexo; por un lado, se observó que los masculinos presentaron una mayor amplitud en el nicho isotópico, lo que implicaría una mayor diversidad en el consumo de recursos. No obstante, por el otro, si bien la amplitud dietaria de los femeninos fue comparativamente menor, fueron quienes mostraron la mayor diferencia en la dieta en relación con los individuos EBB y nichos. Esto último se vincula con los valores de δ13C más negativos; una posible explicación a este patrón refiere a una necesidad biológica diferencial en el consumo de grasa para mantener la fecundidad en este contexto ambiental particular de latitudes altas.
Agradecimientos
A Gisela Cassiodoro y Rafael Goñi por la lectura del manuscrito y sus valiosos comentarios. Al equipo arqueológico del COSC. Al Instituto de Geocronología y Geología Isotópica (INGEIS-UBA/CONICET) y a Estela Ducos y Nazareno Piperissa por su ayuda en el trabajo de laboratorio y espectrometría de masas. A los evaluadores anónimos por las sugerencias y comentarios sobre el manuscrito.
Declaración de financiamiento
Las investigaciones fueron subsidiadas por los proyectos UBACYT 2018-2020 20020170100150BA, PICT-2018-03813 y PICT 2017-2999.
Declaración de disponibilidad de datos
Los datos estan disponibles en las Tablas Suplementarias 1 y 2.
Conflicto de intereses
Los autores declaran que no hay ningun confilcto de intereses.
Material suplementario
Para acceder al material suplementario que acompaña este artículo, visitar https://doi.org/10.1017/laq.2022.100.
Tabla suplementaria 1. Valores isotópicos de individuos enterrados en COSC.
Tabla suplementaria 2. Estadística descriptiva de los distintos marcadores de acuerdo al sexo.











