Non-technical Summary
The Weng'an Biota, found in the Doushantuo Formation in Guizhou Province, South China, is a remarkable fossil assemblage known for its well-preserved ancient life forms. These include small organisms called acritarchs, algae, and even embryo-like fossils. Among these, acritarchs, shaped like spiny spheres, have been essential for understanding the age and relationships of rocks from the Ediacaran Period. Previous studies mainly focused on larger spiny acritarchs, overlooking the smaller ones. In our study, we carefully examined over 500 thin sections and discovered a wealth of well-preserved small and medium-sized acritarchs. These tiny fossils, with diameters ranging 20–150 μm, help us understand the ancient ecosystems and how life evolved during this critical time in Earth's history. We identified several different species of small spiny acritarchs, e.g., Tanarium conoideum, Tanarium elegans, Mengeosphaera membranifera, Mengeosphaera minima, and Variomargosphaeridium gracile. Additionally, we found medium-sized acritarchs, e.g., Tanarium tuberosum and Weissiella cf. W. grandistella. These new findings provide important clues for correlating the rocks of the Doushantuo Formation in the Weng'an area with those in the Yangtze Gorges region. They also help us understand the evolution of acritarchs in different parts of the world, including Australia, Siberia, and the East European Platform.
Introduction
The Ediacaran Period witnessed the evolution of the Earth-Life system from Snowball Earth to the Cambrian explosion, making it one of the most critical transitional periods in the entire geological history (Narbonne et al., Reference Narbonne, Xiao, Shields, Gradstein, Ogg and Schmitz2012; Xiao and Narbonne, Reference Xiao, Narbonne, Gradstein, Ogg, Schmitz and Ogg2020). Many important events in the evolution of life and the environment, e.g., the origin and early radiation of metazoans and the Neoproterozoic Oxygenation Event (NOE), occurred during the Ediacaran Period and left numerous significant geological records worldwide (e.g., Xiao and Knoll, Reference Xiao and Knoll2000, Reference Xiao and Knoll2007). Consequently, a broadly accepted internal subdivision scheme and chronostratigraphic framework for the Ediacaran System is essential, because it forms the basis for studying the coevolution of biology and the environment during this period. However to date, the internal subdivision scheme and a standard for regional stratigraphic correlation for the Ediacaran System are still under debate (Steiner et al., Reference Steiner, Li, Qian, Zhu and Erdtmann2007; Liu et al., Reference Liu, Chen, Zhu, Li, Yin and Shang2014b; Zhou et al., Reference Zhou, Yuan, Xiao, Chen and Hua2019; Zhu et al., Reference Zhu, Yang, Yuan, Li, Zhang, Zhao, Ahn and Miao2019).
Ediacaran large acanthomorphic acritarchs (LAAs, with diameter normally > 200 μm) are globally distributed microfossils and exhibit high diversity, playing a crucial role in biostratigraphic subdivisions and correlation of the Ediacaran System (Vidal, Reference Vidal1990; Vorob'eva et al., Reference Vorob'eva, Sergeev and Knoll2009; Golubkova et al., Reference Golubkova, Raevskaya and Kuznetsov2010; Sergeev et al., Reference Sergeev, Knoll and Vorob'eva2011; Anderson et al., Reference Anderson, Macdonald, Jones, McMahon and Briggs2017). In 2005, Grey established five acritarch assemblage zones based on LAA fossils for the Ediacaran System of Australia (Grey, Reference Grey2005), and this scheme has gained support from other studies (Willman et al., Reference Willman, Moczydłowska and Grey2006; Willman and Moczydłowska, Reference Willman and Moczydłowska2008, Reference Willman and Moczydłowska2011; Sergeev et al., Reference Sergeev, Knoll and Vorob'eva2011), indicating that LAAs could be a valuable tool for the internal subdivision and global correlation of the Ediacaran System.
Since the 1970s, abundant LAAs have been discovered in the Ediacaran Doushantuo Formation in South China (Yin and Li, Reference Yin and Li1978; McFadden et al., Reference McFadden, Xiao, Zhou and Kowalewski2009; Liu et al., Reference Liu, Xiao, Yin, Chen, Zhou and Li2014a; Liu and Moczydłowska, Reference Liu and Moczydłowska2019). Based on extensive investigation of taxonomic diversity, Liu and Moczydłowska (Reference Liu and Moczydłowska2019) established four acritarch assemblage zones for the Ediacaran Doushantuo Formation in the Yangtze Gorges area. However, biostratigraphic correlation between the Ediacaran Doushantuo Formation of Yangtze Gorges area, South China, and the Ediacaran Pertatataka and Julie formations of Australia has long been difficult due to two reasons. First, silicified acanthomorphic acritarchs from chert nodules (Xiao et al., Reference Xiao, Schiffbauer, McFadden and Hunter2010) revealed by thin sections look different from carbonaceous acritarchs extracted from shales using acid maceration, even for the same species. Second, many acritarchs are mainly local species with limited significance in global biostratigraphic correlations. Therefore, how to correlate Ediacaran acritarch assemblage zones of South China with that of Australia, Siberia, and the Eastern European Platform remains uncertain. To resolve the problem, more investigations on acritarch diversity and taxonomy are needed. In addition, although previous studies have established four acritarch assemblage zones for the Ediacaran Doushantuo Formation in the Yangtze Gorges area (Liu and Moczydłowska, Reference Liu and Moczydłowska2019), it remains uncertain whether the scheme of these four biozones is applicable to the Ediacaran Doushantuo Formation in the Weng'an area.
In this study, we focused on the fossil assemblage of acanthomorphic acritarchs from the Ediacaran Doushantuo Formation in the Weng'an area of Guizhou Province, southwestern China. Unlike many small acanthomorphic acritarchs (SAAs, with diameters < 100 μm) found in other regions, e.g., the Yangtze Gorges area in South China, Australia, Siberia, and the East European Platform, the majority of acanthomorphic acritarchs reported previously from the Weng'an Biota have diameters > 200 μm. For instance, Tianzhushania Yin and Li, Reference Yin and Li1978, emend. Yin, Zhou, and Yuan, Reference Yin, Zhou and Yuan2008 and Yinitianzhushania Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a have diameters that can exceed 600 μm. To date, a total of 25 genera and 47 species plus two undetermined species of acanthomorphic acritarchs (Table 1) have been described from the Weng'an Biota. Among them, 22 genera and 28 species belong to the LAAs. It is noteworthy that only eight genera and 10 species plus one undetermined genus of SAAs have been discovered in the Weng'an Biota (L. Yin et al., Reference Yin, Wang, Yuan and Zhou2011). Additionally, seven genera and eight species of medium-sized acritarchs (MAAs, with diameters ranging 100–200 μm) have been identified (Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a). The Weng'an SAAs include Appendisphaera grandis Moczydłowska, Vidal, and Rudavskaya, Reference Moczydłowska, Vidal and Rudavskaya1993, emend. Moczydłowska, Reference Moczydłowska2005; Bullatosphaera sp. indet.; Dicrospinasphaera virgata Grey, Reference Grey2005; Dicrospinasphaera zhangii Yuan and Hofmann, Reference Yuan and Hofmann1998; Eotylotopalla delicata Yin, Reference Yin1987; Hocosphaeridium anozos (Willman in Willman and Moczydłowska, Reference Willman and Moczydłowska2008) Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a; Mengeosphaera chadianensis (Chen and Liu, Reference Chen and Liu1986) Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a; Tanarium digitiforme (Nagovitsin and Faizullin in Nagovitsin et al., Reference Nagovitsyn, Faizullin and Yakshin2004) Sergeev, Knoll, and Vorob'eva, Reference Sergeev, Knoll and Vorob'eva2011; Tanarium victor Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a; Taedigerasphaera lappacea Grey, Reference Grey2005; and Variomargosphaeridium gracile Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a. The MAAs include Asterocapsoides sinensis Yin and Li, Reference Yin and Li1978, emend. Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a; Asterocapsoides wenganensis (Chen and Liu, Reference Chen and Liu1986) Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a; Cavaspina acuminata (Kolosova, Reference Kolosova1991) Moczydłowska, Vidal, and Rudavskaya, Reference Moczydłowska, Vidal and Rudavskaya1993; Eotylotopalla dactylos Zhang et al., Reference Zhang, Yin, Xiao and Knoll1998; Ericiasphaera rigida Zhang et al., Reference Zhang, Yin, Xiao and Knoll1998; Mengeosphaera reticulata (Xiao and Knoll, Reference Xiao and Knoll1999) Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a; Papillomembrana boletiformis Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a; and Taeniosphaera doushantuoensis Liu and Yin, Reference Liu and Yin2005 (Table 1). Considering that the diversity of small and medium-sized acritarchs in the Weng'an Biota has not been fully explored, this study focuses on them to uncover additional clues for Ediacaran biostratigraphic correlation.
Table 1. Small and medium-sized acanthomorphs from the Doushantuo Formation in the Weng'an area, records from the published literature and this study. EEP = East European Platform; 1 = Chen, Reference Chen2004; 2 = Chen and Liu, Reference Chen and Liu1986; 3 = Chen et al., Reference Chen, Yin, Liu, Gao, Tang and Wang2010; 4 = Golubkova et al., Reference Golubkova, Raevskaya and Kuznetsov2010; 5 = Grey, Reference Grey2005; 6 = Knoll, Reference Knoll1992; 7 = Kolosova, Reference Kolosova1991; 8 = Liu et al., Reference Liu, Yin, Chen, Tang and Gao2012; 9 = Liu et al., Reference Liu, Xiao, Yin, Chen, Zhou and Li2014a; 10 = Liu and Moczydłowska, Reference Liu and Moczydłowska2019; 11 = Moczydłowska, Reference Moczydłowska2005; 12 = Moczydłowska and Nagovitsin, Reference Moczydłowska and Nagovitsin2012; 13 = Moczydłowska et al., Reference Moczydłowska, Vidal and Rudavskaya1993; 14 = Nagovitsin et al., Reference Nagovitsyn, Faizullin and Yakshin2004; 15 = Sergeev et al., Reference Sergeev, Knoll and Vorob'eva2011; 16 = Veis et al., Reference Veis, Vorob'eva and Goubkova2006; 17 = Vorob'eva et al., Reference Vorob'eva, Sergeev and Knoll2009; 18 = Vorob'eva et al., Reference Vorob'eva, Sergeev and Chumakov2008; 19 = Willman and Moczydłowska, Reference Willman and Moczydłowska2008; 20 = Willman and Moczydłowska, Reference Willman and Moczydłowska2011; 21 = Xiao, Reference Xiao2004; 22 = Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a; 23 = Xiao and Knoll, Reference Xiao and Knoll1999; 24 = Xiao et al., Reference Xiao, Knoll, Zhang and Hua1999; 25 = Xie et al., Reference Xie, Zhou, Mcfadden, Xiao and Yuan2008; 26 = Yin, Reference Yin1987; 27 = Yin, Reference Yin1990; 28 = Yin and Li, Reference Yin and Li1978; 29 = C. Yin et al., Reference Yin, Liu, Awramik, Chen, Tang, Gao, Wang and Riedman2011; 30 = Yin et al., Reference Yin, Zhu, Knoll, Yuan, Zhang and Hu2007; 31 = L. Yin et al., Reference Yin, Wang, Yuan and Zhou2011; 32 = Yin et al., Reference Yin, Zhu, Tafforeau, Chen, Liu and Li2013; 33 = Yuan and Hofmann, Reference Yuan and Hofmann1998; 34 = Yuan et al., Reference Yuan, Xiao, Yin, Knoll, Zhou and Mu2002; 35 = Zang and Walter, Reference Zang and Walter1992; 36 = Zhang et al., Reference Zhang, Yin, Xiao and Knoll1998; 37 = Zhou et al., Reference Zhou, Brasier and Xue2001; 38 = Zhou et al., Reference Zhou, Chen and Xue2002a; 39 = Zhou et al., Reference Zhou, Xie, Mcfadden, Xiao and Yuan2007; 40 = Zhou et al., Reference Zhou, Yuan and Xiao2002b; 41 = Zhou et al., Reference Zhou, Yuan, Xiao, Chen and Xue2004; 42 = Shang et al., Reference Shang, Liu and Moczydłowska2019; 43 = Ouyang et al., Reference Ouyang, Zhou, Xiao, Guan, Chen, Yuan and Sun2021; * = diameter < 100 μm; ** = medium-sized acritarchs reported by this study; # = species reported previously with diameter <100μm.

Geological setting
Geological investigation of the Doushantuo Formation in the Weng'an area started in the 1960s (Xiao et al., Reference Xiao, Muscente, Chen, Zhou, Schiffbauer, Wood, Polys and Yuan2014b). The Doushantuo Formation in the Weng'an phosphorite mining area, located in Guizhou Province, southwestern China (Fig. 1.1), outcrops in a pattern controlled by a north/northeast to south/southwest-trending Baiyan-Gaoping anticline (Fig. 1.2). A generalized stratigraphic column of the Doushantuo Formation in this area is displayed in Figure 1.3 (Xiao et al., Reference Xiao, Muscente, Chen, Zhou, Schiffbauer, Wood, Polys and Yuan2014b; Cunningham et al., Reference Cunningham, Vargas, Yin, Bengtson and Donoghue2017). The entire formation is composed of five units, arranged from bottom to top as follows: cap dolomite (Unit 1), the lower phosphorite (Unit 2), the middle dolomite (Unit 3), the upper phosphorite (Unit 4), and the phosphoritic dolomite members (Unit 5). Detailed description of lithostratigraphy for the Doushantuo Formation in the Weng'an area can be found in numerous previous publications (Xiao et al., Reference Xiao, Zhang and Knoll1998, Reference Xiao, Zhou, Liu, Wang and Yuan2014a; Yin et al., Reference Yin, Zhu, Davidson, Bottjer, Zhao and Tafforeau2015; Zhou et al., Reference Zhou, Li, Xiao, Lan, Ouyang, Guan and Chen2017).

Figure 1. The geological setting of Weng'an area. (1) Relationships between the Yangtze Block (YB), North China Block (NCB), and Tarim Block (TB). A red star marks the locality of the Weng'an phoshorite mining area. (2) Geological setting of the Weng'an area; the outcrop is marked by a red star. (3) Ediacaran stratigraphic column of the Beidoushan section in the Weng'an area showing the fossil horizon. Modified from Yin et al., Reference Yin, Zhu, Davidson, Bottjer, Zhao and Tafforeau2015.
The Weng'an Biota was described from the upper phosphorite (Unit 4), which consists of the lower black phosphorite facies (Unit 4A) and upper gray phosphatic dolomite facies (Unit 4B). Although the depositional age range for the Doushantuo Formation is well dated, ranging 635–551 Ma (Condon et al., Reference Condon, Zhu, Bowring, Wang, Yang and Jin2005), the Weng'an Biota has long lacked precise isotopic age constraints. There are two karstic surfaces that have developed in the Doushantuo Formation in the Weng'an area, one at the top of Unit 3 and the other within Unit 5 (Zhu et al., Reference Zhu, Zhang and Yang2007, Reference Zhu, Lu, Zhang, Zhao, Li, Aihua, Zhao and Zhao2013, Reference Zhu, Yang, Yuan, Li, Zhang, Zhao, Ahn and Miao2019). If the lower karstic surface can be correlated to the 582 Ma Gaskiers glaciation (Condon et al., Reference Condon, Zhu, Bowring, Wang, Yang and Jin2005), then the Weng'an Biota would be younger than 582 Ma. However, if the upper karstic surface correlates to the Gaskiers glaciation (Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a), then the Weng'an Biota would be much older than 582 Ma. A U-Pb age of 609 ± 5 Ma, determined from the ash bed immediately above Unit 4 of the Ediacaran Doushantuo Formation at the Zhancunping section in Hubei Province, correlates with Unit 4A of the Ediacaran Doushantuo Formation at the Weng'an area. This age constraint provides important chronological information for the Weng'an Biota (Zhou et al., Reference Zhou, Yuan, Xiao, Chen and Hua2019). However, based on new CA-ID-TIMS U-Pb analyses, Yang et al. (Reference Yang, Rooney, Condon, Li, Grazhdankin, Bowyer, Hu, Macdonald and Zhu2021) claimed that zircons from the same ash bed have a detrital origin with maximum depositional age of 612.5 ± 0.9 Ma. Through chemo- and biostratigraphic correlation, Yang et al. (Reference Yang, Rooney, Condon, Li, Grazhdankin, Bowyer, Hu, Macdonald and Zhu2021) proposed that the age of the Weng'an Biota likely ranges 575–590 Ma.
Materials and methods
The specimens reported in this study were collected from the black phosphorite facies (Unit 4A) of the Doushantuo Formation in the Weng'an area (Fig. 1.3). More than 500 thin sections were examined to search for microfossils under a transmitted light microscope (Nikon Ni-U microscope). All acanthomorphic acritarchs revealed in thin sections were recorded with stage coordinates, and then photographed with a Nikon FIT-1 CCD camera. Measurements for vesicle diameter, length, quantity, and basal width of processes were conducted on high-resolution photographs using Image J (http://imagej.nih.gov/ij); the measured data are provided in Appendix 1.
Repository and institutional abbreviation
Microfossils illustrated here are reposited at the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Science, Nanjing, China (NIGPAS).
Systematic paleontology
In this study, we adhered to the criteria established by Xiao et al. (Reference Xiao, Zhou, Liu, Wang and Yuan2014a) to define vesicle size, which is quantitatively diagnosed as small (<100 μm diameter), medium-sized (100–200 μm diameter), or large (>200 μm diameter). In acritarch taxonomy, the characteristics of processes (e.g., morphology, density, branching, and whether hollow processes communicate with the vesicle interior; Fig. 2) play a significant role. A systematic description is given in alphabetic order for acanthomorphic acritarch taxa below.

Figure 2. Schematic drawings of acritarchs: (1) Estrella recta Liu and Moczydłowska, Reference Liu and Moczydłowska2019; (2) Mengeosphaera membranifera Shang, Liu, and Moczydłowska, Reference Liu and Moczydłowska2019; (3) M. minima Liu et al., Reference Liu, Xiao, Yin, Chen, Zhou and Li2014a; (4) Tanarium conoideum Kolosova, Reference Kolosova1991; (5) Tanarium elegans Liu et al., Reference Liu, Xiao, Yin, Chen, Zhou and Li2014a; (6) Tanarium tuberosum Moczydłowska, Vidal, and Rudavskaya, Reference Moczydłowska, Vidal and Rudavskaya1993, emend. Moczydłowska, Reference Moczydłowska2015; (7) Variomargosphaeridium gracile Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a; (8) Weissiella cf. W. grandistella Vorob'eva, Sergeev, and Knoll, Reference Vorob'eva, Sergeev and Knoll2009, emend. Liu and Moczydłowska, Reference Liu and Moczydłowska2019. The left scale bar is for (6) and (8); the right scale bar is for the others.
Group Acritarcha Evitt, Reference Evitt1963
Genus Estrella Liu and Moczydłowska, Reference Liu and Moczydłowska2019
Type species
Estrella greyae Liu and Moczydłowska, Reference Liu and Moczydłowska2019 from the Yangtze Gorges area, northern Xiaofenghe section, South China.
Estrella recta Liu and Moczydłowska, Reference Liu and Moczydłowska2019
Figures 2.1, 3
- Reference Liu and Moczydłowska2019
Estrella recta Liu and Moczydłowska, p. 109, fig. 57A–F.
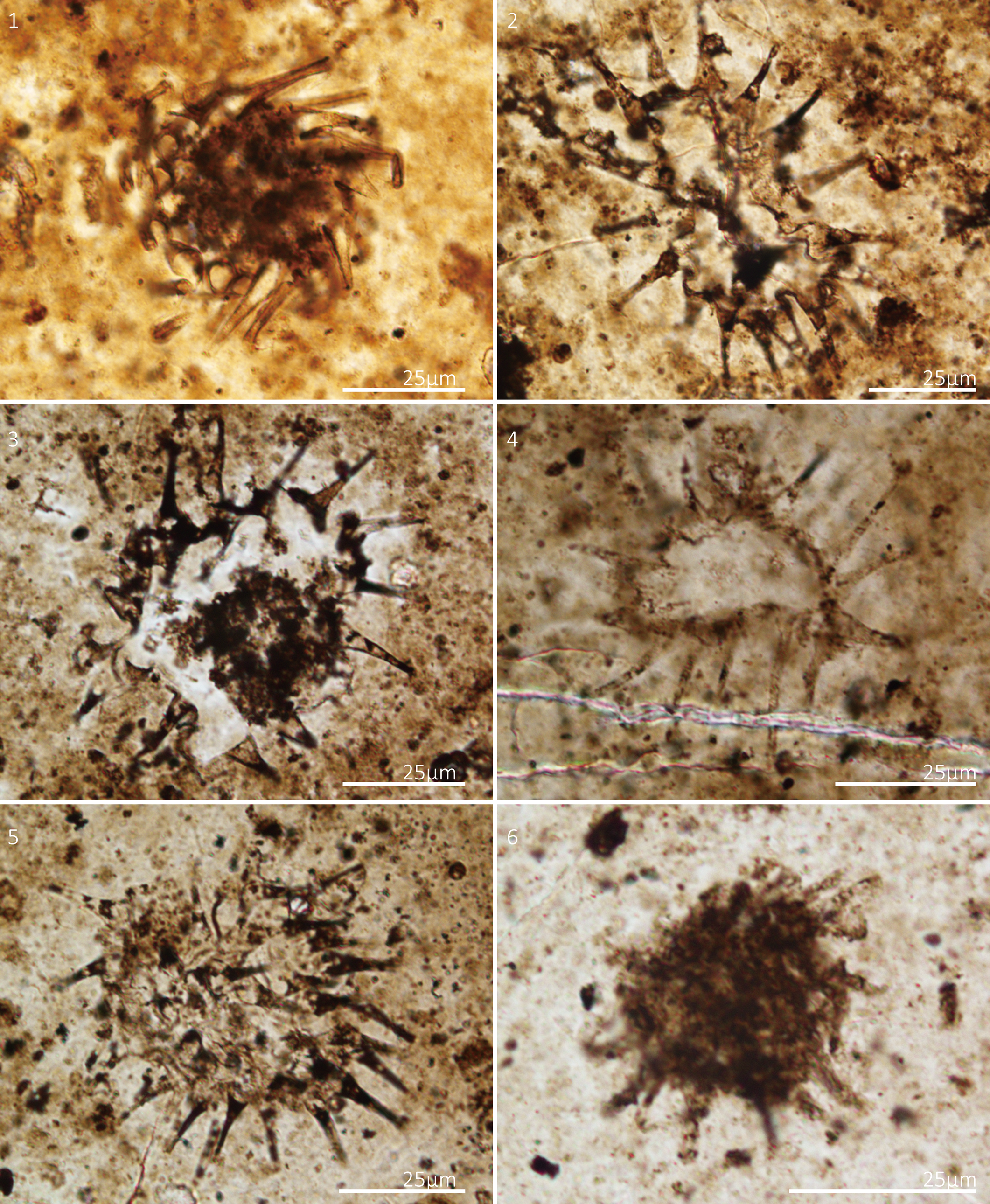
Figure 3. Estrella recta Liu and Moczydłowska, Reference Liu and Moczydłowska2019. Thin section numbers: (1) WS17-225-X42Y105, (2) WS17-225-X46Y106, (3) WS17-195-X54Y99, (4) WS17-157-X64Y101, (5) WS17-259-X19.5Y113, (6) WS17-211-X35Y94.
Holotype
Thin section nos. IGCAGS-D2XFH200 and XFH0946-1-9, F48/4 from the Yangtze Gorges area, Nantuocun section, Member II of the Doushantuo Formation, South China (Liu and Moczydłowska, Reference Liu and Moczydłowska2019, p.108, fig. 57A–D).
Description
Small-sized spherical vesicle with an irregular outline resulting from collapse during taphonomic and diagenetic processes, bearing homomorphic and regularly distributed hollow processes. The processes are elongated tubular structures that taper toward their distal portions, ending in sharp tips. They are hollow and freely communicate with the vesicle cavity. The widened bases of processes create a wavy outline of the vesicle wall. Vesicle diameter 35–48 μm (mean 43 μm, N = 3); process length 8–27 μm (mean 19 μm, N = 47); process basal width 2–11 μm (mean 5 μm, N = 14); distance between processes 6–12 μm (mean 9 μm, N = 13); ~16–25 processes on the vesicle periphery in circumferential view.
Material
Five well-preserved specimens and one poorly preserved specimen from Unit 4A of the Ediacaran Doushantuo Formation in the Weng'an area.
Remarks
The species is distinguished by its robust hollow processes, characterized by an elongate conical shape and bulbous bases. Vesicles exhibit a circular shape with a slightly undulating outline (Fig. 3). Due to taphonomy and diagenesis, many specimens have undergone deformation. Consequently in some specimens, the length of processes exceeds the vesicle diameter, whereas in others, the length of processes is only half of the vesicle diameter. Although sharing similarities in vesicle size and process length with several species of the genus Tanarium Kolosova, Reference Kolosova1991—e.g., Tanarium gracilentum (Yin in Yin and Liu, Reference Yin, Liu, Zhao, Xing, Ding, Liu and Zhao1988) Ouyang et al., Reference Ouyang, Zhou, Xiao, Guan, Chen, Yuan and Sun2021, Tanarium pycnacanthum Grey, Reference Grey2005, and Tanarium paucispinosum Grey, Reference Grey2005—the species differs in process morphology and density. Both Tanarium gracilentum and Tanarium pycnacanthum feature more densely distributed processes (>100 processes in circumferential view). In contrast, Tanarium paucispinosum (with <10 processes in circumferential view) possesses more slender and more flexible processes compared to our specimens. Consequently, our specimens are classified as Estrella recta.
Genus Mengeosphaera Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a
Type species
Mengeosphaera chadianensis (Chen and Liu, Reference Chen and Liu1986) Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a from the Doushantuo Formation at Weng'an area, South China.
Mengeosphaera membranifera Shang, Liu, and Moczydłowska, Reference Liu and Moczydłowska2019
Figures 2.2, 4
- Reference Shang, Liu and Moczydłowska2019
Mengeosphaera membranifera Shang et al., p. 18, fig. 15A–D.
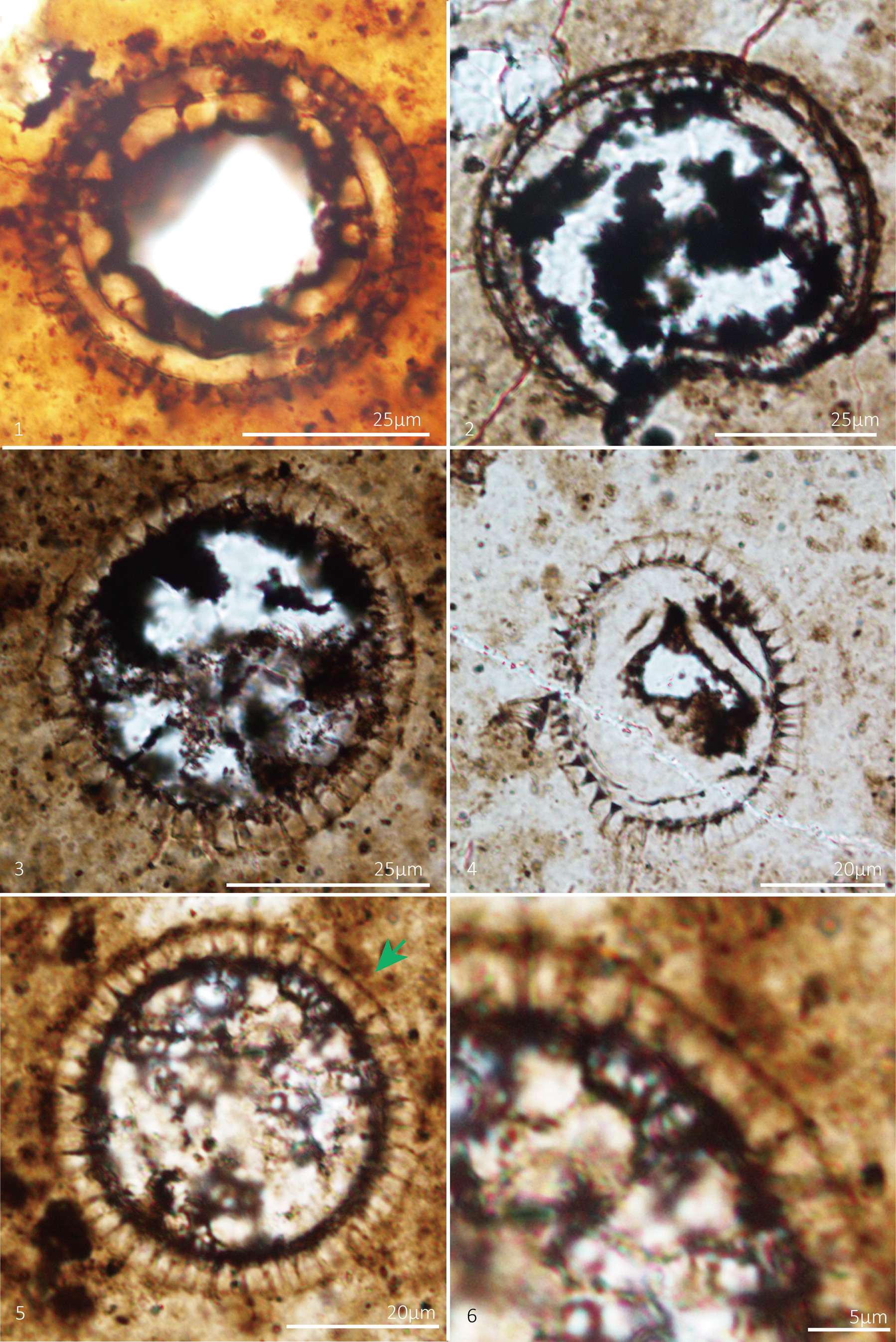
Figure 4. Mengeosphaera membranifera Shang, Liu, and Moczydłowska, Reference Liu and Moczydłowska2019. Thin section numbers: (1) WS17-152-X62Y103, (2) WS17-231-X64Y108, (3) WS17-231-X27Y101, (4) WS17-204-X37Y93, (5) WS17-207-X30Y104, with green arrow indicating site of (6), (6) magnified view of (5).
Holotype
Thin section no. LJ101115-1-7 from the Songlin area of Guizhou Province, Liujing section of chert nodules in medium- to thick-bedded dolostones of the Ediacaran Doushantuo Formation, South China (Shang et al., Reference Shang, Liu and Moczydłowska2019, p. 18, fig. 15A–D).
Description
Small spheroidal vesicles exhibiting abundant, closely, and regularly arranged biform processes of uniform length, each surrounded by an outer membrane. The outer membrane is supported by processes, and the distance from the outer membrane to the bases of the processes remains constant within a single specimen. Processes consist of regular conical bases and cylindrical, thin apical spines with sharp tips. They are hollow and freely communicate with the vesicle interior. Vesicle diameter 40–48 μm (mean 43 μm, N = 4); process length 4–7 μm (mean 6 μm, N = 185); process basal width 1–2 μm (mean 2 μm, N = 17); distance between processes 1–4 μm (mean 3 μm, N = 19); ~40 processes in circumferential view.
Material
Five specimens from Unit 4A of the Ediacaran Doushantuo Formation in the Weng'an area.
Remarks
Acritarchs featuring an outer membrane supported by biform processes are rare in the Ediacaran Period, with known occurrences limited to chert nodules in the Liujing section, Songlin area of Guizhou Province, southwestern China (Shang et al., Reference Shang, Liu and Moczydłowska2019). Our specimens exhibit similarities to some Mengeosphaera gracilis Liu et al., Reference Liu, Xiao, Yin, Chen, Zhou and Li2014a in terms of process morphology and share resemblance with species of Cymatiosphaeroides (Knoll, Reference Knoll1984) Knoll, Swett, and Mark, Reference Knoll, Swett and Mark1991 in the outer membrane structure. However, they differ from Mengeosphaera gracilis due to the presence of the outer membrane and are distinguished from Cymatiosphaeroides by their distinct biform processes. Therefore, despite its smaller size compared to the specimens described by Shang et al. (Reference Shang, Liu and Moczydłowska2019), our specimens align more closely with the diagnostic features of Mengeosphaera membranifera.
Mengeosphaera minima Liu et al., Reference Liu, Xiao, Yin, Chen, Zhou and Li2014a
Figures 2.3, 5, 6
- Reference Liu, Xiao, Yin, Chen, Zhou and Li2014a
Mengeosphaera minima Liu et al., p. 69, 101, figs. 51.8, 63.
- Reference Ouyang, Zhou, Xiao, Guan, Chen, Yuan and Sun2021
Mengeosphaera minima; Ouyang et al., p. 24, fig. 17M, N.
- Reference Ye, Li, Tong, An, Hu and Xiao2022
Mengeosphaera minima; Ye et al., p. 52, fig. 32G, H.
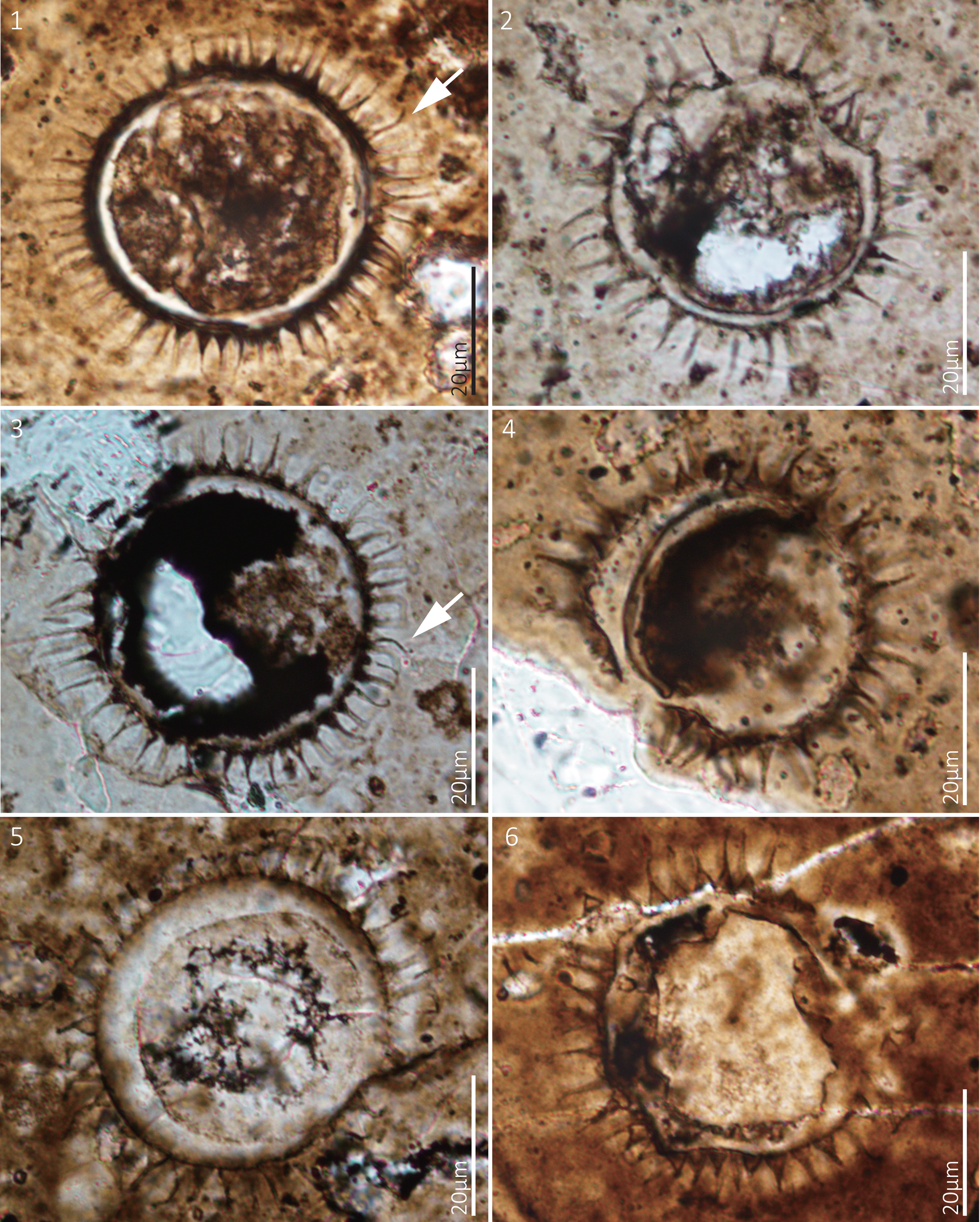
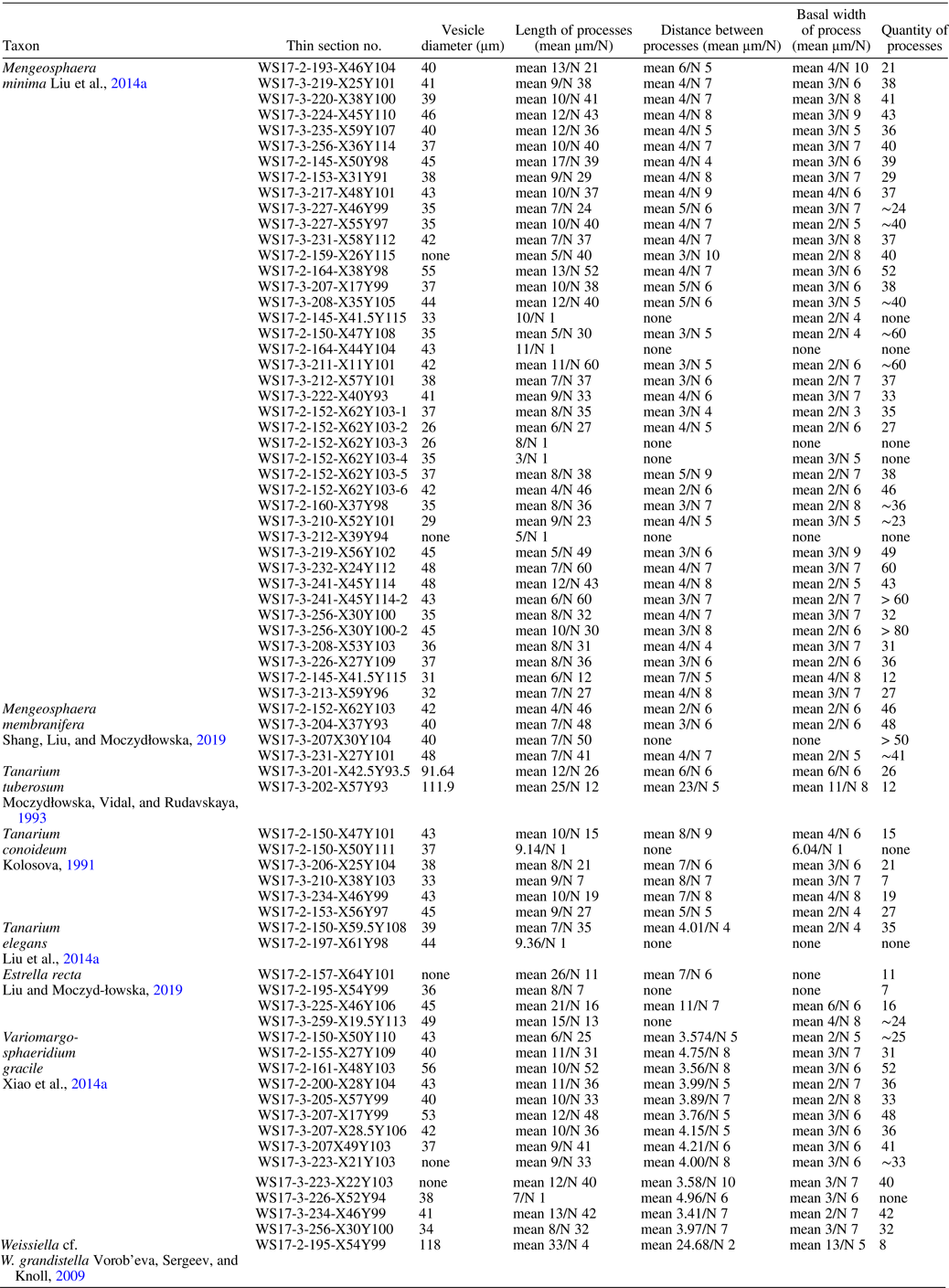
Figure 5. Mengeosphaera minima Liu et al., Reference Liu, Xiao, Yin, Chen, Zhou and Li2014a. Thin section numbers: (1) WS17-217-X48Y101, (2) WS17-153-X31Y91, (3) WS17-227-x57y97, (4) WS17-227-X46Y99, (5) WS17-231-X58Y112, (6) WS17-145-X50Y98.

Figure 6. Mengeosphaera minima Liu et al., Reference Liu, Xiao, Yin, Chen, Zhou and Li2014a. Thin section numbers: (1) WS17-224-X45Y110, (2) WS17-235-X59Y107, (3) WS17-207-x17y99, (4) WS17-256-X36Y114, (5) WS17-220-X38Y100, (6) WS17-219-X25Y101. (1, 2) showing the multicellular structures within their vesicle (blue arrows).
Holotype
Thin section no. I IGCAGS-NPIII-090A from the Yangtze Gorges area, Niuping section of the upper Member III, Doushantuo Formation, South China (Liu et al., Reference Liu, Xiao, Yin, Chen, Zhou and Li2014a, p. 69, fig. 63.1).
Description
Small spheroidal vesicle bearing abundant, closely, and evenly distributed biform processes. Each process consists of a conical base and a thin apical filament. Occasionally, the apical spine is curved at the distal end (Figs. 5.1, 5.3, 6.1, 6.4, white arrows). The processes are hollow and freely communicate with the vesicle interior. Some specimens show multicellular structure (Fig. 6.1, 6.2, blue arrows). Vesicle diameter 33–46 μm (mean 39 μm, N = 40); process length 5–13 μm (mean 8 μm, N = 1,432); process basal width 2–4 μm (mean 3 μm, N = 246); process spacing 3–6 μm (mean 4 μm, N = 233); ~21–60 processes on vesicle periphery in circumferential view.
Material
Forty-two specimens from Unit 4A of the Ediacaran Doushantuo Formation in the Weng'an area.
Remarks
The specimens illustrated in Figures 5 and 6 align with the diagnostic criteria of Mengeosphaera minima (Liu et al., Reference Liu, Xiao, Yin, Chen, Zhou and Li2014a; Ouyang et al., Reference Ouyang, Zhou, Xiao, Guan, Chen, Yuan and Sun2021; Ye et al., Reference Ye, Li, Tong, An, Hu and Xiao2022). The presence of the curved spine might be a taphonomic artifact, because this feature was not consistently observed. Although approximately a dozen species of Mengeosphaera exhibit conical expansion of the process bases (please refer the fig. 51 and table 2 in Liu et al., Reference Liu, Xiao, Yin, Chen, Zhou and Li2014a), only M. bellula Liu et al., Reference Liu, Xiao, Yin, Chen, Zhou and Li2014a, M. minima, M. spicata Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a, and M. stegosauriformis Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a share small vesicle sizes comparable to our specimens. Notably, the basal parts of the processes in M. spicata and M. stegosauriformis are larger (basal width 7–20 μm for M. spicata and 20–22 μm for M. stegosauriformis, according to Liu et al., Reference Liu, Xiao, Yin, Chen, Zhou and Li2014a). Additionally, M. bellula displays a different conical shape than M. minima.
Genus Tanarium Kolosova, Reference Kolosova1991, emend. Moczydłowska et al., Reference Moczydłowska, Vidal and Rudavskaya1993
Type species
Tanarium conoideum Kolosova, Reference Kolosova1991, emend. Moczydłowska, et al., Reference Moczydłowska, Vidal and Rudavskaya1993, from the Ediacaran (Vendian) Kursov Formation in the Yakutia area of the Siberian Platform (Kolosova, Reference Kolosova1991; Moczydłowska et al., Reference Moczydłowska, Vidal and Rudavskaya1993; Moczydłowska, Reference Moczydłowska2005).
Tanarium conoideum Kolosova, Reference Kolosova1991, emend. Moczydłowska et al., Reference Moczydłowska, Vidal and Rudavskaya1993
Figures 2.4, 7
- Reference Kolosova1991
Tanarium conoideum Kolosova, p. 56, 57, fig. 5.1–5.3.
- Reference Moczydłowska, Vidal and Rudavskaya1993
Tanarium conoideum; Moczydłowska et al., p. 514–516, fig. 10C, D.
- Reference Liu, Xiao, Yin, Chen, Zhou and Li2014a
Tanarium conoideum; Liu et al., p. 109, figs. 76.2, 77.1–77.6.
- non Reference Prasad and Ramson2016
Tanarium conoideum; Prasad and Ramson, Reference Prasad and Ramson2016, p. 56, pl. 7, fig. 8.
- Reference Shang, Liu and Moczydłowska2019
Tanarium conoideum; Shang et al., p. 26, fig. 16A–E.
- non Reference Yang, Pang, Chen, Zhong and Yang2020
Tanarium conoideum; Yang et al., p. 6, 7, fig. 2K.
- Reference Vorob'eva and Petrov2020
Tanarium conoideum; Vorob'eva and Petrov, p. 374, 375, pl. 1, fig. 15.
- Reference Yang, Pang, Chen, Zhong and Yang2020
Tanarium conoideum; Yang et al., p. 8, fig. 2K.
- Reference Ye, Li, Tong, An, Hu and Xiao2022
Tanarium conoideum; Ye et al., p. 61, fig. 42.
Holotype
Thin section no. YIGS Nr 87-115 from the Kursov Formation, Upper Proterozoic, Vendian of Yakulia, Siberian Piatform and Borehole Byk-Tanar area (Kolosova, Reference Kolosova1991, p. 56, fig. 5.1, 5.2).
Description
Small spherical vesicles, bearing a small number of conical processes. The bases of the processes are slightly expanded and distally tapered to a pointed tip, or sometimes blunt due to breakage. The processes are hollow, allowing for free communication with the vesicle interior. Vesicle diameter 33–45 μm (mean 40 μm, N = 6); process length 8–10 μm (mean 7 μm, N = 89); process basal width 3–4 μm (mean 4 μm, N = 27); distance between processes 5–8 μm (mean 7 μm, N = 35); ~7–27 processes on the vesicle periphery in circumferential view.
Material
Six specimens from Unit 4A of the Ediacaran Doushantuo Formation in the Weng'an area.
Remarks
Most processes are straight, with a few showing distal hooks (Fig. 7.2, 7.3, white arrows). According to Xiao et al. (Reference Xiao, Zhou, Liu, Wang and Yuan2014a), distally hooked processes can result from taphonomic alteration. Although the specimens in our collection are smaller than those previously reported for Tanarium conoideum, the process features align more closely with the diagnosis of Tanarium conoideum than with other species. The specimens diagnosed as Tanarium conoideum by Moczydłowska and Nagovitsin (Reference Moczydłowska and Nagovitsin2012) and Prasad and Ramson (Reference Prasad and Ramson2016), exhibit thin, short processes that are inconsistent with the diagnosis of Tanarium conoideum. Similarly, the specimen described as Tanarium conoideum by Yang et al. (Reference Yang, Pang, Chen, Zhong and Yang2020) has abundant, relatively short, densely distributed processes with basal expansions, leading to its exclusion from Tanarium conoideum.
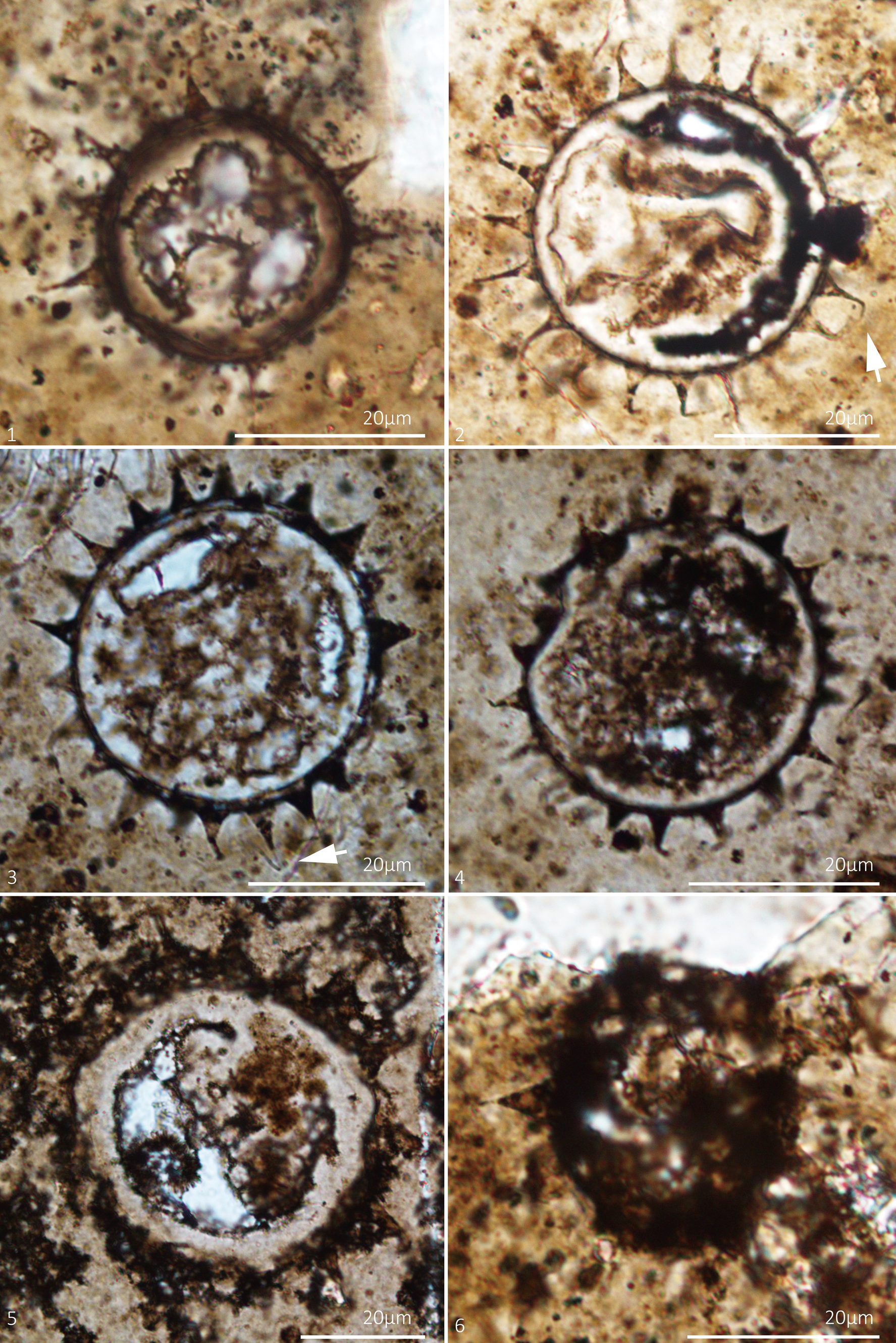
Figure 7. Tanarium conoideum Kolosova, Reference Kolosova1991, emend. Moczydłowska et al., Reference Moczydłowska, Vidal and Rudavskaya1993. Thin section numbers: (1) WS17-210-X38Y103, (2) WS17-150-X47Y101, (3) WS17-234-X46Y99, (4) WS17-206-X25Y104, (5) WS17-208-X48Y98, (6) WS17-150-X50Y111.
Tanarium elegans Liu et al., Reference Liu, Xiao, Yin, Chen, Zhou and Li2014a
Figures 2.5, 8.1, 8.2
- Reference Liu, Xiao, Yin, Chen, Zhou and Li2014a
Tanarium elegans Liu et al., p. 81, fig. 75.8–75.16.

Figure 8. (1, 2) Tanarium elegans Liu et al., Reference Liu, Xiao, Yin, Chen, Zhou and Li2014a and (3, 4) Tanarium tuberosum Moczydłowska, Vidal, and Rudavskaya, Reference Moczydłowska, Vidal and Rudavskaya1993, emend. Moczydłowska, Reference Moczydłowska2015. Thin section numbers: (1) WS17-196-X40Y107, (2) WS17-150-X59.5Y108, (3) WS17-201-x42.5y93.5, (4) WS17-202-X57Y93.
Holotype
Thin section no. IGCAGS–XFH–270 from the Yangtze Gorges area, Xiaofenghe section of the lower Member II, Doushantuo Formation, South China (Liu et al., Reference Liu, Xiao, Yin, Chen, Zhou and Li2014a, fig. 75.10).
Description
Small spheroidal vesicles exhibiting numerous evenly and regularly arranged conical processes. These processes, hollow in nature, freely communicate with the vesicle interior, and taper gradually to a blunt termination. Vesicle diameter 39 and 44 μm (only two specimens discovered during this study); process length 6–9 μm (mean 7 μm, N = 36); process basal width 1–3 μm (mean 2 μm, N = 4); > 30 processes in circumferential view.
Material
Two specimens from Unit 4A of the Ediacaran Doushantuo Formation in the Weng'an area.
Remarks
The specimens illustrated in Figures 8.1 and 8.2 align with the diagnostic criteria of Tanarium elegans. Tanarium elegans exhibits closest similarities to Tanarium acus Liu et al., Reference Liu, Xiao, Yin, Chen, Zhou and Li2014a and Xenosphaera liantuoensis Yin, Reference Yin1987, emend. Liu et al., Reference Liu, Xiao, Yin, Chen, Zhou and Li2014a in terms of small vesicle size and very thin processes. However, when compared to Tanarium elegans, Tanarium acus features relatively fewer but longer processes whereas Xenosphaera liantuoensis has a larger vesicle and longer processes. In addition, it is noteworthy that Tanarium elegans typically lacks a thin filamentous tip.
Tanarium tuberosum Moczydłowska, Vidal, and Rudavskaya, Reference Moczydłowska, Vidal and Rudavskaya1993,
emend. Moczydłowska, Reference Moczydłowska2015
Figures 2.6, 8.3, 8.4
- Reference Moczydłowska, Vidal and Rudavskaya1993
Tanarium tuberosum Moczydłowska et al., p. 516, fig. 15B–D.
- Reference Moczydłowska2015
Tanarium tuberosum; Moczydłowska, pl. 3, figs. 1–6.
- Reference Prasad and Ramson2016
Tanarium tuberosum; Prasad and Ramson, p. 56, 58, pl. 8, figs. 1, 2.
- Reference Liu and Moczydłowska2019
Tanarium tuberosum; Liu and Moczydłowska, p. 153–156, fig. 86, pl. 21, fig. 18A.
- Reference Shang, Liu and Moczydłowska2019
Tanarium tuberosum; Shang et al., p. 27–29, fig. 18.
- Reference Ouyang, Zhou, Xiao, Guan, Chen, Yuan and Sun2021
Tanarium tuberosum; Ouyang et al., p. 27, fig. 19R
Holotype
Thin section no. PMU-Sib.4-J/30/3 from the Nepa-Botuoba area, lowermost Kbamaka Formation, Neoproterozoic, Upper Vendian (Moczydłowska et al., Reference Moczydłowska, Vidal and Rudavskaya1993, p. 516, fig. 15B–D).
Description
Medium-sized spheroidal vesicle showcasing irregularly distributed wide conical processes. These processes, hollow and broad, feature blunt tips and freely communicate with the vesicle interior. Vesicle diameter 91 and 111 μm (only two specimens discovered during this study); process length 12–25 μm (mean 16 μm, N = 38); process basal width 6–11 μm (mean 9 μm, N = 14); distance between processes 6–23 μm (mean 14 μm, N = 11); ~12–26 processes on vesicle periphery in circumferential view.
Material
Two specimens from Unit 4A of the Ediacaran Doushantuo Formation in the Weng'an area.
Remarks
The current specimens exhibit irregularly conical processes and a medium-sized vesicle diameter (91–111 μm). The vesicle size is comparable to the specimens featured by Liu and Moczydłowska (Reference Liu and Moczydłowska2019). There is a slight difference in the process number of the specimens in Figures 8.3 and 8.4 that could be interpreted as infraspecific variability (Liu and Moczydłowska, Reference Liu and Moczydłowska2019).
Genus Variomargosphaeridium Zang and Walter, Reference Zang and Walter1992, emend. Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a
Type species
Variomargosphaeridium litoschum Zang and Walter, Reference Zang and Walter1992, Doushantuo Formation at Weng'an area, South China; Amadeus Basin, Rodinga 4 borehole at a depth of 48.38–48.67 m, Pertatataka Formation, Ediacaran successions in Australia.
Variomargosphaeridium gracile Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a
Figures 2.7, 9
- Reference Xiao, Zhou, Liu, Wang and Yuan2014a
Variomargosphaeridium gracile Xiao et al., p. 58, fig. 36.
- Reference Ouyang, Zhou, Xiao, Guan, Chen, Yuan and Sun2021
Variomargosphaeridium gracile; Ouyang et al., p. 31, fig. 22A.

Figure 9. Variomargosphaeridium gracile Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a. Thin section numbers: (1) WS17-150-X50Y110, (2) WS17-161-X48Y103, (3) WS17-234-X46Y99, (4) WS17-207-X17Y99. These four specimens show bifurcated structures at the tops of the processes (blue arrows).
Holotype
Thin section no. Slide 87ZW15-4, CPC27760, from the Pertatataka Formation, Ediacaran successions in Australia (Zhang and Walter, Reference Zang and Walter1992, p. 114, 117, figs. 63D–G, 88).
Description
Small-sized spheroidal vesicle bearing evenly and densely distributed branching processes. These processes consist of conical bases and bifurcate tips, being thin, hollow, and freely communicating with the vesicle interior. Vesicle diameter 34–55 μm (mean 42 μm, N = 11); process length 6–13 μm (mean 10 μm, N = 449); process basal width 2–3 μm (mean 3μm, N = 84); distance between processes 3–5 μm (mean 4 μm, N = 87); ~25–52 processes in circumferential view.
Material
Thirteen specimens from Unit 4A of the Ediacaran Doushantuo Formation in the Weng'an area.
Remarks
The specimens are identified as Variomargosphaeridium gracile based on vesicle size and process morphology. Although V. litoschum and V. floridum Nagovitsin and Moczydłowska in Moczydłowska and Nagovitsin, Reference Moczydłowska and Nagovitsin2012 also exhibit branching processes, they are considerably larger in both vesicle and process sizes compared to our specimens (Grey, Reference Grey2005; Moczydłowska and Nagovitsin, Reference Moczydłowska and Nagovitsin2012; Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a). In addition, the processes of V. floridum branch distally to form an apical crown of branchlets, and the branching pattern of V. litoschum is more complex than that of V. gracile (Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a).
Genus Weissiella Vorob'eva, Sergeev, and Knoll, Reference Vorob'eva, Sergeev and Knoll2009, emend. Liu and Moczydłowska, Reference Liu and Moczydłowska2019
Type species
Weissiella grandistella Vorob'eva, Sergeev, and Knoll, Reference Vorob'eva, Sergeev and Knoll2009, emend. Liu and Moczydłowska, Reference Liu and Moczydłowska2019 from the East European Platform and 2605.5 m depth of keltminsk 1 borehole, Ediacaran and Yangtze Gorges area, Doushantuo Formation.
Weissiella cf. W. grandistella Vorob'eva, Sergeev, and Knoll, Reference Vorob'eva, Sergeev and Knoll2009,
emend. Liu and Moczydłowska, Reference Liu and Moczydłowska2019
Figures 2.8, 10
- Reference Vorob'eva, Sergeev and Knoll2009
Weissiella grandistella Vorob'eva, Sergeev, and Knoll, p. 183–185, figs. 10.1a–f.
- Reference Liu and Moczydłowska2019
Weissiella grandistella; Liu and Moczydłowska, p. 163, figs. 91F, G, 92A–G.
- Reference Ouyang, Zhou, Xiao, Guan, Chen, Yuan and Sun2021
Weissiella cf. W. grandistella; Ouyang et al., p. 40, 41, fig. 24A–H.
- Reference Ye, Li, Tong, An, Hu and Xiao2022
Weissiella cf. W. grandistella; Ye et al., p. 70, fig. 49D–F.
Holotype
Thin section no. 14700-13, from the Ediacara, upper part of Vychegda Formation (Vorob'eva et al., Reference Vorob'eva, Sergeev and Knoll2009, p. 183–185, figs. 10.1a–f).
Description
Medium-sized spheroidal vesicle featuring a few large processes. These processes, hollow and conical, exhibit a rounded or blunt end and have transverse crosswalls that are flat or distally convex. Vesicle diameter 118 μm (from specimen in Fig. 10.1); process length 33 μm (mean 33 μm, N = 4); process basal width 13 μm (mean 13 μm, N = 5); distance between processes 3–5 μm (mean 3 μm, N = 2); ~25–52 processes on vesicle periphery in circumferential view.

Figure 10. Weissiella cf. W. grandistella Vorob'eva, Sergeev, and Knoll, Reference Vorob'eva, Sergeev and Knoll2009, emend. Liu and Moczydłowska, Reference Liu and Moczydłowska2019. Thin section numbers: (1) WS17-2-195-X54Y101. (2–4) Magnified views of (1) showing crosswall structures: (2) red arrow; (3) green arrow; (4) blue arrow.
Material
One specimen from Unit 4A of the Ediacaran Doushantuo Formation in the Weng'an area.
Remarks
The specimen exhibits similarities to Weissiella grandistella in terms of process morphology, length, and crosswalls. However, its vesicle diameter is smaller compared to that reported for W. grandistella specimens from the East European Platform (vesicle diameter 450–500 μm). Following the description of W. cf. W. grandistella by Ouyang et al. (Reference Ouyang, Zhou, Xiao, Guan, Chen, Yuan and Sun2021) and Ye at al. (Reference Ye, Li, Tong, An, Hu and Xiao2022), we categorize our specimen as W. cf. W. grandistella.
Summary of taxonomy
After examining >500 thin sections of black phosphorite, we discovered numerous acanthomorphic acritarchs, the majority of which belong to LAAs with a diameters > 200 μm. Although we identified several new species of Yinitianzhushania that might have not been reported previously (which will be discussed in a separate paper), most of the specimens can be attributed to previously reported genera. This report specifically delves into SAAs, particularly those with diameters <60 μm (Fig. 11). These diminutive specimens have been infrequently reported by previous researchers in the Weng'an Biota. The abundance of these SAAs is significantly lower than that of LAAs. Due to their smaller size, they are more difficult to detect under the microscope. We discovered >70 specimens of SAAs (< 60 μm in diameter; Fig. 11) and three specimens of MAAs (no more than 150 μm in diameter; Fig. 11).

Figure 11. Measurements for process length (left) and process quantity (right) versus vesicle diameter of acanthomorphic acritarchs in the Weng'an Biota.
Based on detailed observations and microscopic imaging (Fig. 12), we characterized the vesicle morphology and structure of these small and medium-sized acritarchs, including the shape, length, and density of their processes (specific descriptions are in the Systematic paleontology section). Through meticulous morphological comparison, we preliminarily identified seven species from four genera, namely Tanarium conoideum, Tanarium elegans, Tanarium tuberosum, Mengeosphaera membranifera, M. minima, Estrella recta, and Variomargosphaeridium gracile, and one possibly new form tentatively placed in open nomenclature, Weissiella cf. W. grandistella. The abundance of Mengeosphaera and Tanarium specimens was relatively higher, accounting for > 60% of the discovered specimens, whereas the abundance of the other taxa was relatively low. Mengeosphaera was identified by its biform process, whereas Tanarium was identified by its conical process; Estrella by the length of its processes exceeding the vesicle diameter; Variomargosphaeridium by branching hollow processes; and Weissiella by short conical to cylindrical processes with internal crosswalls.

Figure 12. The distribution of the Ediacaran small and medium-sized acanthomorphic acritarchs in the present study (modified from Li et al., Reference Li, Bogdanova, Collins, Davidson, Waele, Ernst, Fitzsimons, Fuck, Gladkochub, Jacobs, Karlstrom, Lu, Natapov, Pease, Pisarevsky, Thrane and Vernikovsky2008).
Tanarium conoideum was previously reported in the Weng'an Biota by an earlier study (Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a). However, the specimens that they reported had a diameter >100 μm, whereas the specimens that we reported for the same species had a diameter of no more than 60 μm. The other SAAs and MAAs from three genera and five species (Estrella recta, Mengeosphaera membranifera, Mengeosphaera minima, Tanarium elegans, and Tanarium tuberosum) mentioned above were discovered for the first time in the Weng'an Biota. These fossils not only provide new materials for reconstructing the composition of the fossil assemblage of the Weng'an Biota, but also offer new evidence for gaining a more comprehensive understanding of the marine ecosystem at that time.
Moreover, these SAAs are not exclusive to the Weng'an Biota. Some common acritarch elements have also been found in the Yangtze Gorges region of South China, Australia, Siberia, and the East European Platform (Fig. 12). For example, Estrella recta and Mengeosphaera bellula were found in both the Weng'an and the Yangtze Gorges region of South China, whereas Tanarium tuberosum was found in the Weng'an and the Yangtze Gorges region of South China, the Easten Officer Basin of South Australia, the Siberian Platform of Russia, and the Timan Ridge of the East European Platform. Tanarium conoideum was found in the Weng'an, the Yangtze Gorges of South China, and the Siberia Platform of Russia. Although their phylogenetic affinities are still disputed, these globally distributed acanthomorphic acritarchs have the potential to be further explored in the subdivision and global/regional correlation of the Ediacaran system.
Implications for biostratigraphic correlation
Given the widespread distribution of these newly discovered acanthomorphic acritarchs in the Weng'an Biota, they have the potential to contribute to stratigraphic division and correlation within the Ediacaran System. In 2014, Liu et al. established two biozones based on acritarch fossil assemblages from the Ediacaran Doushantuo Formation in the Yangtze Gorges area: the lower acritarch biozone, dominated by Tianzhushania spinosa Yin and Li, Reference Yin and Li1978, emend. C. Yin in Yin and Liu, Reference Yin, Liu, Zhao, Xing, Ding, Liu and Zhao1988, and the upper acritarch biozone, dominated by Tanarium conoideum, Hocosphaeridium scaberfacium Zhang in Zhang and Walter, Reference Zang and Walter1992, and Hocosphaeridium anozos (see Liu et al., Reference Liu, Xiao, Yin, Chen, Zhou and Li2014a). Liu and Moczydłowska (Reference Liu and Moczydłowska2019) proposed four new biozones based on the latest fossil data, in ascending order: the Appendisphaera grandis-Weissiella grandistella-Tianzhushania spinosa Assemblage Zone, Tanarium tuberosum-Schizofusa zangwenlongii Assemblage Zone, Tanarium conoideum-Cavaspina basiconica Assemblage Zone, and Tanarium pycnacanthum-Ceratosphaeridium glaberosum Assemblage Zone.
Among the newly discovered SAAs and MAAs in the Weng'an Biota, Tanarium tuberosum from the Yangtze Gorges area belongs to the Tanarium tuberosum-Schizofusa zangwenlongii Assemblage Zone, whereas Tanarium conoideum comes from the Tanarium conoideum-Cavaspina basiconica Assemblage Zone. Considering the presence of Tianzhushania spinosa in the Weng'an Biota, it belongs to the Appendisphaera grandis-Weissiella grandistella-Tianzhushania spinosa Assemblage Zone in the Yangtze Gorges area. In the Doushantuo Formation of the Weng'an area, these taxa mainly come from a < 2 m thick layer of black phosphorite (Unit 4A). This contrasts with the Yangtze Gorges area, where the distribution ranges of acritarchs in the four biozones do not overlap. This situation poses challenges for stratigraphic correlation.
The acritarch fossil assemblage from the Weng'an Biota, as presented in this report, originates from the same stratigraphic horizon, namely Unit 4A of the Doushantuo Formation in the Weng'an area. However, the acritarch fossils of the same genus and species in the Yangtze Gorges area are distributed across three different fossil zones within Member II of the Doushantuo Formation. Several factors could contribute to this discrepancy. We believe that the 2 m thick black phosphorite (Unit 4A) is highly condensed, and the fossils in the Weng'an Biota are not in situ but rather redeposited after undergoing repeated transportation and reworking within the basin (Yin et al., Reference Yin, Liu, Li, Tafforeau and Zhu2014; Bottjer et al., Reference Bottjer, Yin, Zhao and Zhu2020). This might explain why acritarchs that were originally expected to come from three different fossil zones (i.e., different stratigraphic layers) coexist in one 2 m thick stratigraphic layer in the Weng'an area's Doushantuo Formation. Despite this, previous studies have reported 24 genera and 69 species of acritarchs from Member Ⅱ of the Doushantuo Formation at three sections (Jiulongwan, Jinguadun, and Wuzhishan) in the Yangtze Gorges area (Ouyang et al., Reference Ouyang, Zhou, Xiao, Guan, Chen, Yuan and Sun2021). Two taxa (Mengeosphaera minima and Weissiella cf. W. grandistella) in this study are consistent with those reported by Ouyang et al. (Reference Ouyang, Zhou, Xiao, Guan, Chen, Yuan and Sun2021). It is recommended to correlate Unit 4A of the Doushantuo Formation containing the Weng'an Biota to the Ediacaran Doushantuo Formation Member Ⅱ in the Yangtze Gorges area using acritarch fossils (Xiao et al., Reference Xiao, Zhou, Liu, Wang and Yuan2014a; Liu and Moczydłowska, Reference Liu and Moczydłowska2019; Ouyang et al., Reference Ouyang, Zhou, Xiao, Chen and Shao2019).
In 2005, Grey identified the Tanarium conoideum-Schizofusa risoria-Variomargosphaeridium litoschum Assemblage Zone in the Australian Ediacaran strata. Some acritarch species from this biozone have also been found in the Weng'an area, including Tanarium conoideum and Variomargosphaeridium litoschum. Consequently, Xiao et al. (Reference Xiao, Zhou, Liu, Wang and Yuan2014a) suggested correlating the stratigraphic level of the Weng'an Biota to the Tanarium conoideum-Schizofusa risoria-Variomargosphaeridium litoschum Assemblage Zone in the Australian Ediacaran System. The new findings in this study are consistent with this perspective. However, considering the co-occurrence of acritarch fossils from different biozones in the same black phosphorite horizon (i.e., Unit 4A of the Doushantuo Formation) in the Weng'an area, it is challenging to separate their distribution ranges in the stratigraphic sequence, and such correlations should be made cautiously. In the future, conducting higher-resolution stratigraphic sampling of acanthomorphic acritarch fossils in the black phosphorite and gray phosphatic dolomite of the Weng'an Doushantuo Formation could contribute to a more comprehensive resolution of this issue.
Conclusion
This study discovered seven species and one possible new form tentatively placed in open nomenclature (Weissiella cf. W. grandistella) of SAAs and MAAs in the Weng'an Biota, including five species reported for the first time in the Weng'an area. Together with the previously reported fossil materials, there are a total of 50 species and one undetermined species of acanthomorphs in the Weng'an Biota (Table 1). Although some of these acritarchs are local species, most of these species have also been found in other regions, e.g., the Yangtze Gorges area (South China), Australia, and Siberia. The presence of so many shared acritarch species among different regions plays a crucial role in the subdivision and correlation of the Ediacaran System. Based on the current data, the acritarch assemblages of the Doushantuo Formation in the Weng'an area share common species with three acritarch assemblage zones (Appendisphaera grandis–Weissiella grandistella–Tianzhushania spinosa Assemblage Zone, Tanarium tuberosum–Schizofusa zangwenlongii Assemblage Zone, and Tanarium conoideum–Cavaspina basiconica Assemblage Zone) from the lower part of Member II of the Ediacaran Doushantuo Formation in the Yangtze Gorges area, as well as the Tanarium conoideum–Schizofusa risoria–Variomargosphaeridium litoschum Assemblage Zone from the Ediacaran Pertatataka Formationin in Australia. This taxonomic similarity indicates a correlation of Unit 4A of the Ediacaran Doushantuo Formation in the Weng'an area, Member II of the Ediacaran Doushantuo Formation in the Yangtze Gorges area, and the upper part of the Ediacaran Pertatataka Formation in Australia.
The fossils reported in this report were collected from the black phosphorites of the Doushantuo Formation in Weng'an. Similar to the chert nodules in the Yangtze Gorges area, the study of acritarchs in black phosphorites can only be conducted through thin sections. Because the information obtained from acritarchs observed in thin sections differs to some extent from the compressed organic acritarchs obtained from shale through acid maceration, achieving consistent taxonomic criteria for acritarchs from different taphonomic windows becomes challenging. This inconsistency somewhat undermines the potential of acritarchs in biostratigraphic subdivision and correlation. Therefore, achieving higher-resolution biostratigraphic correlation of the Ediacaran System in the future not only requires detailed layer-by-layer sampling in different localities, but also needs to overcome the preservation bias and inconsistency in taxonomic criteria for acritarchs from different taphonomic windows by introducing new techniques and methods.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (42022010, 41921002, U2244202). We would like to thank Professor C. Zhou for providing suggestions about schematic drawings, and we thank the guest editor T. Selly. We are also most grateful for the detailed and constructive comments provided by two anonymous referees.
Declaration of competing interests
The authors declare none.
Appendix 1. Measured data of acritarchs in this study.