



Introduction
Graptolites are known to contribute to the establishment of lower Paleozoic terrane in Malaysia, the study of which was pioneered by Clive Roderick Jones together with various other field geologists in the late 1950s and early 1960s. The earliest record of graptolite work was made by Jones (Reference Jones1959) while investigating the geology of northwest Peninsular Malaysia from a band of shales and limestone formations on the Langkawi islands. More graptolite localities were later found in Langkawi as well as in the surrounding area, including the mainland Kedah and Perlis states, which led to the publication of concise graptolite reports by Jones (Reference Jones1973, Reference Jones1978). Subsequent studies have been done on the stratigraphy of the area by other authors. For example, Cocks et al. (Reference Cocks, Fortey and Lee2005) erected a new stratigraphical nomenclature for the successions of the area while Hassan et al. (Reference Hassan, Erdtmann, Wang-Xiaofeng and Peng2013) reviewed a monograptid from the exposures of Devonian black shale in Perlis state. Nevertheless, little study has been done on the systematic paleontology of the Ordovician and Silurian graptolites after Jones's last publication, and the biostratigraphy is lacking in revision for the past 40 years. The latest Ordovician graptolites in particular are of great interest because they mark the earliest graptolite fauna so far recorded from the Malaysian Peninsula. Furthermore, this interval, which is regarded as the Hirnantian, is relatively brief but significant in the geological record for documenting the second largest global extinction bioevent in Earth history. Two pulses of extinction event have been identified, which seem to correspond with the start and end of the Late Ordovician glaciation (Harper et al., Reference Harper, Hammarlund and Rasmussen2014). The purpose of this paper is to provide a systematic description of the graptolites from the Ordovician–Silurian boundary of northwest Peninsular Malaysia. Graptolites collected previously by Jones are reexamined and updated herein while new specimens that have been recently recovered are described in detail. A revised graptolite biozonation is also suggested for this interval, which would allow for regional and global correlation.
Geological setting
The Langkawi archipelago is located at the most northwestern edge of Peninsular Malaysia and comprises a collection of 99 islands and islets. The region is part of the northwestern domain of the Peninsular Malaysia Western Belt and is predominantly of Paleozoic rocks (Fig. 1). In Langkawi, the lower Paleozoic graptolites can be found in the Tanjung Dendang Formation (formerly the Lower Detrital Member), which is part of the Setul Group. The name of the formation is taken after Pulau Tanjung Tembus Dendang, an island northeast of Langkawi, but the type section is located on an island west of it, Pulau Langgun, on the south shore of Teluk Mempelam between grid references 6°26′40″N, 99°53′00″E and 6°26′42″N, 99°53′02″E (Jones, Reference Jones1966).

Figure 1. The location of the Langkawi Islands in the northwestern edge of Peninsular Malaysia. The Setul Group in Langkawi is concentrated mostly on the eastern fringe of the island archipelago. The Setul Group is constrained inside the northwestern domain of the peninsula Western Belt, consisting of predominantly Paleozoic rocks.
The Tanjung Dendang Formation in Pulau Langgun is ~27 m thick overall and consists of well-bedded quartzites through siltstone to dark shales and sometimes includes bedded cherts, which have been recorded to span from the Hirnantian (Late Ordovician) to the top of the Aeronian Stage of the Llandovery Series (lower Silurian). Graptolites are present mainly in the well-bedded, carbonaceous siltstones and shales where the individual bed is generally less than 10 cm in thickness. The Tanjung Dendang Formation overlies the Ordovician Kaki Bukit Limestone Formation (formerly the Lower Setul Limestone), which is a very hard, thickly bedded, gray stylolithic limestone with an abrupt contact (Cocks et al., Reference Cocks, Fortey and Lee2005; Lee, Reference Lee, Hutchison and Tan2009) (Fig. 2.1). The uppermost unit of the Kaki Bukit Limestone has been described to be of late Sandbian–early Katian age using conodont fossils, suggesting the existence of an unconformity with the base of the Tanjung Dendang Formation (Niko et al., Reference Niko, Sone and Leman2019). An exposed bed that represents the Hirnantian stage can be found on a vertical rock face near a small cave, away from the coast, and positioned slightly higher than the high tide line (Fig. 2.2). Walking along the shore, the interbedded dark shale and cherty strata are exposed in an almost horizontal succession, dipping to the northeast. A moderate number of graptolite specimens can be recovered on the raised mudstone exposure just after a nearby sandy ditch and can be observed only when the tide is low (Fig. 2.3).

Figure 2. (1) The stratigraphy of the northwest coast of Pulau Langgun, Langkawi (modified from Jones, Reference Jones1978). The graptolites were recovered from the Ordovician–Silurian Tanjung Dendang Formation strata that can be found in Teluk Mempelam area. (2) The basal section exposure of the Tanjung Dendang Formation that overlies the Kaki Bukit Limestone where graptolites from the uppermost Ordovician age have been recovered. (3) The raised exposure where graptolites of the earliest Silurian age have been recovered. Note that the exposure can be observed only when the tide is at its lowest.
Materials and methods
A total of 26 slabs of graptolite-bearing mudstone collected previously by Jones were obtained from the repository of the Department of Mineral and Geoscience Malaysia in Ipoh, Perak (172F –176F, 178F–183F, 185F–190F, 191-1F, 196F–198F, 200F-1, 200F-2, 202F, 204F, 205F). The materials were recovered from the basal 1.1 m of Tanjung Dendang Formation in Pulau Langgun, Langkawi, which Jones (Reference Jones1973) noted as the lower graptolite beds. At least nine samples recovered more recently from the same interval have been included to supplement the graptolite data. Several shelly specimens from the overlying sandy Hirnantia fauna bed were also retrieved from this succession. Above the Hirnantia fauna bed, we have taken fresh samples at ~1 m intervals each to recover the smaller number of graptolites that would be present in the mudstone layers. Seventeen specimens have been taken from LG4, nine from LG7, seven from LG8, and four from LG9 and are housed in the Geosciences Department, Universiti Teknologi PETRONAS. Specimens that have been prepared were observed and photographed using an Olympus SZX16 stereomicroscope mounted with a digital camera and Andonstar ADSM302 digital microscope (a smaller, affordable microscope that can be used for working at home). Illustrations of the graptolites were made to highlight the outline and important details such as the proximal astogeny and thecal morphology and for morphometric measurements involving the dorsoventral width (DVW), two-theca repeat distance (2TRD, as defined by Howe, Reference Howe1983), tubarium length, virgella length, and other quantifiable features. The collected data were later compared with literature, and revisions were made if necessary.
Repositories and institutional abbreviations
Specimens examined in this study are deposited in the following institutions: Department of Mineral and Geoscience Malaysia (JMG), Perak, Malaysia, or Geosciences Department, Universiti Teknologi PETRONAS (UTP), Perak, Malaysia.
Biostratigraphy
Metabolograptus extraordinarius Biozone.—Overlying the thickly bedded stylolithic limestone with an abrupt and possibly unconformable contact, but underlying the Hirnantian shelly sandstone layer, is a 60 cm thick unit of dark, hard, and thinly bedded graptolitic mudstone with discontinuous lamination. Most-abundant graptolites can be recovered in the middle of the section and in some instances cover almost the entire bedding surface. Previously, Jones had identified an indifferently preserved graptolite fauna that included rare Neodiplograptus modestus (Lapworth, Reference Lapworth1876), abundant Metabolograptus persculptus (Elles and Wood, Reference Elles and Wood1907), and occasionally Normalograptus temalaensis (Jones, Reference Jones1973) (Jones, Reference Jones1968, Reference Jones1973, Reference Jones1978). He then assigned this section and the basal part of the overlying sandstone bed to the persculptus Biozone and, thus, assigned the graptolite-bearing succession to the basal Silurian, with the Ordovician–Silurian boundary located at the contact with the limestone unit below, following practice current at the time (Jones, Reference Jones1973, Reference Jones1978). It was more than 20 years before Cocks et al. (Reference Cocks, Fortey and Lee2005) reviewed the stratigraphy and established the section to be instead of the latest Ordovician (Hirnantian) age. Subsequently, we have taken a chance to reexamine Jones's specimens and, together with our own recently collected specimens, use them to restudy this noteworthy section (Fig. 3). From the result of our research, a more diverse graptolite fauna could be identified, which includes Anticostia aff. A. lata (Elles and Wood, Reference Elles and Wood1906), Korenograptus elegantulus (Mu and Ni, Reference Mu and Ni1983), Metabolograptus extraordinarius (Sobolevskaya, Reference Sobolevskaya and Obut1974), Metabolograptus ojsuensis (Koren’ and Mikhaylova in Koren et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980), Neodiplograptus shanchogensis (Li, Reference Li1984) (Li, Reference Li1984b), Normalograptus ajjeri (Legrand, Reference Legrand1977), Normalograptus aff. N. mirnyensis (Obut and Sobolevskaya in Obut et al., Reference Obut, Sobolevskaya and Nikolaev1967), and Normalograptus temalaensis (Jones, Reference Jones1973) (Figs. 4, 5). The faunal assemblages suggested an earlier extraordinarius Biozone for the section underlying the shelly Hirnantia fauna bed compared with the previously thought persculptus Biozone. This finding seems to match well with biostratigraphy schemes in the nearby regions such as in the Mandalay Region in Myanmar (Chen et al., Reference Chen, Chen, Aung and Muir2020) and South China (Chen et al., Reference Chen, Rong, Li and Boucot2004), suggesting a close paleogeographical tie, but it seems to differ from that reported in southern Thailand (Wongwanich et al., Reference Wongwanich, Burrett, Tansathein and Chaodumrong1990). In the Mandalay region, graptolites that can be found from the extraordinarius Biozone include Korenograptus acanthus (Lin in Mu et al., Reference Mu, Li, Ge, Chen, Lin and Ni1993), Korenograptus selectus Chen et al., Reference Chen, Chen, Aung and Muir2020, Neodiplograptus incommodus Legrand, Reference Legrand2009, Normalograptus ajjeri (Legrand, Reference Legrand1977), Normalograptus angustus (Perner, Reference Perner1895), Normalograptus pseudovenustus (Legrand, Reference Legrand, Hughes, Rickards and Chapman1986), Metabolograptus ojsuensis (Koren’ and Mikhaylova in Koren et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980), and Metabolograptus extraordinarius (Sobolevskaya, Reference Sobolevskaya and Obut1974) (Chen et al., Reference Chen, Chen, Aung and Muir2020). The Global Boundary Stratotype Section and Point (GSSP) for the base of the Hirnantian Stage itself was proposed at a point below the base of the Kuanyinchiao Bed in the Wangjiawan North section (Chen et al., Reference Chen, Rong, Fan, Zhan, Mitchell, Harper, Melchin, Peng, Finney and Wang2006) and coincided with the first appearance of Metabolograptus extraordinarius (Sobolevskaya, Reference Sobolevskaya and Obut1974) and a slightly earlier appearance of Metabolograptus ojsuensis (Koren’ and Mikhaylova in Koren et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980). In the Satun province of southern Thailand, however, the index fossil Metabolograptus persculptus (Elles and Wood, Reference Elles and Wood1907) instead was reported from the lower member of Wang Tong Formation, below the Hirnantia fauna bed (Rickards in Wongwanich et al., Reference Wongwanich, Burrett, Tansathein and Chaodumrong1990). The Wang Tong Formation is the stratigraphic equivalent of the Tanjung Dendang Formation (Wongwanich et al., Reference Wongwanich, Burrett, Tansathein and Chaodumrong1990; Cocks et al., Reference Cocks, Fortey and Lee2005), and in theory, they should share the same fauna at this level. However, without any graptolite illustrations to confirm this, it is perhaps more appropriate to restudy the Satun material. At the Vinini Creek section in north central Nevada, the interval defined by the extraordinarius Biozone possesses a moderate species diversity that includes the index species Metabolograptus extraordinarius (Sobolevskaya, Reference Sobolevskaya and Obut1974) and Metabolograptus ojsuensis (Koren’ and Mikhaylova in Koren et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980), which are abundant throughout, while normalograptids such as Normalograptus ajjeri (Legrand, Reference Legrand1977), Normalograptus mirnyensis (Obut and Sobolevskaya in Obut et al., Reference Obut, Sobolevskaya and Nikolaev1967), and rare Normalograptus angustus (Perner, Reference Perner1895) also make their first appearance in the interval (Štorch et al., Reference Štorch, Mitchell, Finney and Melchin2011). Diplograptina such as A. lata (Elles and Wood, Reference Elles and Wood1906) do occur but made their last appearance within the extraordinarius Biozone, which coincides with the first appearance of the normalograptids in the Vinini Creek section (Mitchell et al., Reference Mitchell, Sheets, Belscher, Finney, Holmden, Laporte, Melchin and Patterson2007; Štorch et al., Reference Štorch, Mitchell, Finney and Melchin2011); the same pattern can also be seen in South China (Chen et al., Reference Chen, Fan, Melchin and Mitchell2005a, Reference Chen, Melchin, Sheets, Mitchell and Fanb). The dramatic faunal change in this level is attributable to the planktonic graptolite communities under environmental stress due to the climate change leading up to the Late Ordovician mass extinction event (Sheets et al., Reference Sheets, Mitchell, Melchin, Loxton, Štorch, Carlucci and Hawkins2016).

Figure 3. The stratigraphy and occurrence of the graptolites of the uppermost Ordovician to the lowermost Silurian of Tanjung Dendang Formation in Pulau Langgun, Langkawi.

Figure 4. Graptolites of the Metabolograptus extraordinarius Biozone in Pulau Langgun, Langkawi. (1–7) Metabolograptus ojsuensis (Koren’ and Mikhaylova in Koren’ et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980): (1) 191F-1_1; (2) 179F_1; (3) 175F_3; (4) 188F_1; (5) 176F_2; (6) 173F_1; (7) 176F_4. (8, 9) Anticostia aff. A. lata (Elles and Wood, Reference Elles and Wood1906): (8) 179F_3; (9) 179F_4. (10–20) Metabolograptus extraordinarius (Sobolevskaya, Reference Sobolevskaya and Obut1974): (10) 190F_6; (11) 174F_3; (12) 173F_3; (13) LG1_01; (14) LG1_04-1; (15) 174F_2; (16) LG1_04-2; (17) LG1_02; (18) 187F_6; (19) 174F_1; (20) LG1_03.

Figure 5. Graptolites of the Metabolograptus extraordinarius Biozone in Pulau Langgun, Langkawi. (1, 2) Neodiplograptus shanchongensis (Li, 1984): (1) 180F_3; (2) 173F_5. (3–5) Korenograptus elegantulus (Mu and Ni, Reference Mu and Ni1983): (3) 182F_4; (4) 183F_3; (5) 172F_3. (6–9) Normalograptus aff. N. mirnyensis (Obut and Sobolevskaya in Obut et al., Reference Obut, Sobolevskaya and Nikolaev1967): (6) LG1_06-2; (7) 185F_5; (8) 190F_3; (9) LG1_05. (10–12) Normalograptus ajjeri (Legrand, Reference Legrand1977): (10) 180F_1; (11) 187F_5; (12) 175F_1. (13–17) Normalograptus temalaensis (Jones, Reference Jones1973): (13) 189F_3; (14) 187F_3; (15) 172F_2; (16) 205F_2; (17) 187F_7.
Shelly Hirnantia fauna bed
The Hirnantia fauna bed is represented by 50 cm of interbedded dark-grey mudstone and fine sandstone at the bottom part and ~30 cm gray, thick-bedded, very fine-grained sandstone at the upper part, in which dark-brown shelly materials are pervasive. Only incomplete specimens of trilobites (most likely Mucronaspis sp.) could be recovered from the thickly bedded upper part together with evidence of shelly fauna fragments (Fig. 6). Previously, Jones (Reference Jones1973, Reference Jones1978) referred to the beds as the Dalmanitina siltstone bed (bed 2) and described it as a thickly bedded siltstone and gritty subgraywacke with shelly fossils. Several shelly fossils were recognized, including gastropods (Megolomphala? sp., Lophospira sp.) and the trilobite Dalmanitina malayensis Kobayashi and Hamada, Reference Kobayashi and Hamada1964. Subsequently, Cocks et al. (Reference Cocks, Fortey and Lee2005) regarded Dalmanitina malayensis as a junior synonym of Mucronaspis mucronata (Brongniart in Brongniart and Desmarest, Reference Brongniart and Desmarest1822). The Hirnantia fauna bed can be recognized by fauna of distinctive brachiopods and trilobites associated with the Late Ordovician Hirnantian glaciation and subsequent drop in sea level occurring in wide ranges of settings (Brenchley, Reference Brenchley, Cocks and Rickards1988; Harper et al., Reference Harper, Hammarlund and Rasmussen2014; Goldman et al., Reference Goldman, Sadler, Leslie, Melchin, Agterberg, Gradstein, Gradstein, Ogg, Schmitz and Ogg2020). It is noted to represent the survival fauna of the early part of the Late Ordovician mass extinction and is used as an important tool of correlation worldwide (Harper et al., Reference Harper, Hammarlund and Rasmussen2014). Cocks et al. (Reference Cocks, Fortey and Lee2005) noted, however, that it is unlikely for the Sibumasu terrane to suffer more than a relatively cool climate during the glaciation period. A similar occurrence with the section in Langkawi can be observed in the Wangjiawan North section of China, where an interval of argillaceous limestone yielding Hirnantia fauna occurred between the extraordinarius and persculptus biozones (Chen et al., Reference Chen, Rong, Fan, Zhan, Mitchell, Harper, Melchin, Peng, Finney and Wang2006). While global correlation data suggest the first and last appearance of the shelly Hirnantia fauna to be diachronous (Rong et al., Reference Rong, Chen and Harper2007, Reference Rong, Harper, Huang, Li, Zhang and Chen2020), the majority occurs within the extraordinarius Biozone and disappears in the persculptus Biozone (= within the lower-middle Hirnantian) (Chen et al., Reference Chen, Rong, Fan, Zhan, Mitchell, Harper, Melchin, Peng, Finney and Wang2006; Rong et al., Reference Rong, Harper, Huang, Li, Zhang and Chen2020). Close to the Malaysian border in the Satun Province of Southern Thailand, a correlative Hirnantia fauna bed can be observed from the middle member of the Wang Tong Formation at locality 8 (Cocks and Fortey, Reference Cocks and Fortey1997). The similar facies change in Thailand is noted to represent the local expression of a global eustatic regression associated with the latest Ordovician glaciation, and the fauna occurring in the section include the trilobite Mucronaspis mucronata (Brongniart in Brongniart and Desmarest, Reference Brongniart and Desmarest1822) (Cocks and Fortey, Reference Cocks and Fortey1997; Cocks et al., Reference Cocks, Fortey and Lee2005). The Thailand records, however, differ somewhat from our observation in Langkawi by having the appearance of the shelly Hirnantia fauna within the reported persculptus Biozone (Cocks and Fortey, Reference Cocks and Fortey1997; Wongwanich et al., Reference Wongwanich, Tansathien, Leevongcharoen, Paengkaew, Mantajit, Potisat and Wongwanich2002).

Figure 6. Shelly fossils from the Hirnantia bed in Pulau Langgun, Langkawi. (1–5) Mucronaspis sp.: (1) pygidium; (2) incomplete mold with part of the thorax and pygidium; (3) counterpart to (1); (4) thoracic axial lobe; (5) counterpart to (4). Scale bars = 1 mm.
Metabolograptus persculptus Biozone.—Directly above the Hirnantia fauna bed lies an interbedded dark-gray mudstone unit. The rocks are much more fissile with continuous lamination and break easily into even layers. It is, overall, less carbonaceous than the mudstone from the extraordinarius Biozone. The beddings are distinct at the bottom of this section but become much more deformed going up the succession. The first 1 m of the section has been designated as LG4. Several graptolite fragments have been recovered near the base, which includes Anticostia aff. A. lata (Elles and Wood, Reference Elles and Wood1906), Avitograptus? sp., Metabolograptus persculptus (Elles and Wood, Reference Elles and Wood1907), Normalograptus angustus (Perner, Reference Perner1895), and Normalograptus rhizinus (Li and Yang in Nanjing Institute of Geology and Mineral Resources, 1983) (Fig. 7). The presence of the index species Metabolograptus persculptus (Elles and Wood, Reference Elles and Wood1907) in particular suggests a persculptus Biozone assignment for the section above the Hirnantia fauna bed. The graptolites found are relatively scarce and fragmented, likely due to taphonomic processes. Jones previously regarded this section as the lower part of the cherty beds (unit 3), which he described as a closely bedded black quartzite and siltstone without any fossils in it (Jones, Reference Jones1973, Reference Jones1978). Although it is true that the majority of this section is indeed unfossiliferous, the presence of fragmentary yet distinct graptolites that are notably different from the previous biozone warrants a proper biozonal assignment for this section. Our observation suggests that the Ordovician–Silurian boundary could occur somewhat higher in the succession above the Hirnantia fauna bed, perhaps within the unfossiliferous interval. In South China, the base of the Lungmachi black shales of the Yangtze Platform also contains a fauna consisting of normalograptids and neodiplograptids, including Normalograptus angustus (Perner, Reference Perner1895), Normalograptus mirnyensis (Obut and Sobolevskaya in Obut et al., Reference Obut, Sobolevskaya and Nikolaev1967), Neodiplograptus shanchongensis (Li, Reference Li1984) (Li, Reference Li1984b), and Neodiplograptus modestus (Lapworth), which completely replaces the previous diplograptine fauna by the end of the persculptus Biozone interval (Chen et al., Reference Chen, Fan, Melchin and Mitchell2005a; Fan et al., Reference Fan, Melchin, Chen, Wang, Zhang, Chen, Chi and Chen2011). In the Zhejiang Province, the graptolites recovered from the persculptus Biozone of the Anji biota mudstone interval include Avitograptus acanthocystus (Fang et al., Reference Fang, Liang, Zhang and Yu1990) and Avitograptus akidomorphus Muir et al., Reference Muir, Zhang, Botting and Ma2020, which may represent the morphological intermediates between Avitograptus avitus (Davies, Reference Davies1929) and similar akidograptines (Muir et al., Reference Muir, Zhang, Botting and Ma2020). Elsewhere in China, in the Tielugou section, Shennongjia Anticline, Hubei Province, possible specimens of Metabolograptus persculptus (Elles and Wood, Reference Elles and Wood1907) have been recognized from the lowest 1.25 m interval; however, overall faunas are poorly preserved (Maletz et al., Reference Maletz, Wang, Kai and Wang2021b). Common graptolites include Normalograptus mirnyensis (Obut and Sobolevskaya in Obut et al., Reference Obut, Sobolevskaya and Nikolaev1967) and Korenograptus elegantulus (Mu and Ni, Reference Mu and Ni1983). In the Mandalay region of Myanmar, overlying the Hwe Mawng Purple Shale Member, in which the Hirnantia fauna can be found, is the shale of the Panghsa-pye Formation, where graptolite assemblages include Normalograptus angustus (Perner, Reference Perner1895), Normalograptus mirnyensis (Obut and Sobolevskaya in Obut et al., Reference Obut, Sobolevskaya and Nikolaev1967), Neodiplograptus incommodus Legrand, Reference Legrand2009, Neodiplograptus mandalayensis Chen et al., Reference Chen, Chen, Aung and Muir2020, and Avitograptus cf. A. avitus (Davies, Reference Davies1929), which possibly indicate a persculptus Biozone considering that the eponymous index species is absent in the section (Chen et al., Reference Chen, Chen, Aung and Muir2020). Close by in north of Satun, southern Thailand, abundant graptolites had been reported from the black shales that correlate with the upper member of the Wang Tong Formation (Agematsu et al., Reference Agematsu, Sashida, Salyapongse and Sardsud2006). These include Normalograptus pseudovenustus pseudovenustus (Legrand, Reference Legrand, Hughes, Rickards and Chapman1986) and Normalograptus sp. Normalograptus pseudovenustus is noted to be a reliable index species of the Ordovician–Silurian boundary interval. However, the proximal end of the graptolites illustrated does not seem to include the typical pattern H astogeny that can be found in normalograptids; for example, the base of th11 is at or slightly above the level of the sicular aperture, but further revision could help to corroborate this matter.
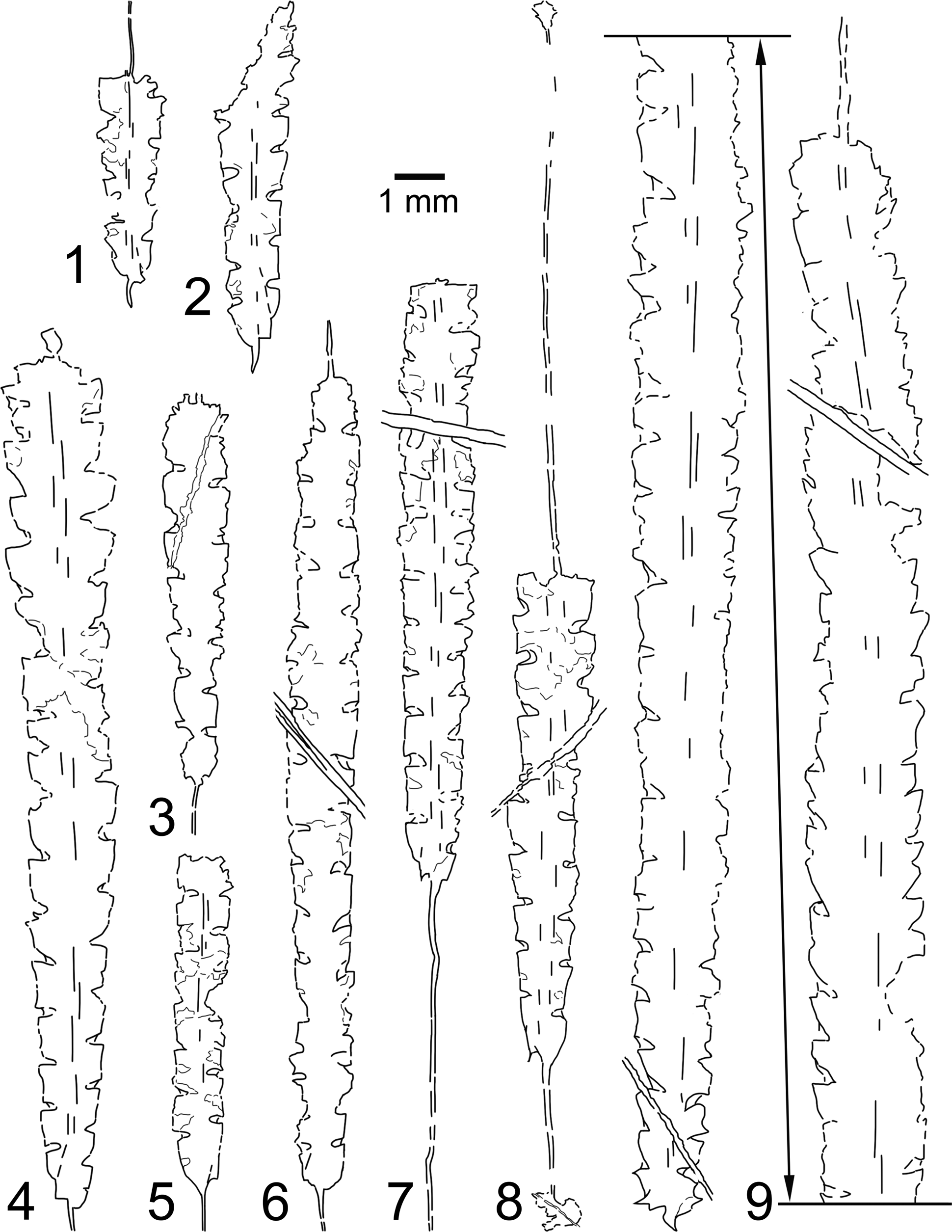
Figure 7. Graptolites of the Metabolograptus persculptus Biozone in Pulau Langgun, Langkawi. (1) Avitograptus? sp., LG4L_05. (2, 4) Metabolograptus persculptus (Elles and Wood, Reference Elles and Wood1907): (2) LG4L_04; (4) LG4L_01. (3, 5, 6) Normalograptus angustus (Perner, Reference Perner1895): (3) LG4M_06; (5) LG4L_06; (6) LG4L_02. (7, 8) Normalograptus rhizinus (Li and Yang in Nanjing Institute of Geology and Mineral Resources, 1983): (7) LG4M_07-2; (8) LG4M_02. (9) Anticostia aff. A. lata (Elles and Wood, Reference Elles and Wood1906), LG4M_07-1.
Akidograptus ascensus–Parakidograptus acuminatus Biozone.—Around 3 m above the base of the persculptus Biozone interval is a dark mudstone exposure, just after a nearby sandy ditch. Graptolites have been collected from intervals designated as LG7 to LG9 in this succession. A moderate number of graptolites can be recovered from the thin, fissile mudstone intervals, which are usually sandwiched between thicker, more silicified cherty mudstone layers. Note that the apparent bottom part of the exposure (lower LG7) is usually infested with barnacles and sessile sea life due to the rising sea tide. The graptolites recovered become more fragmented above the LG9 interval as the rocks become much harder and silicified. Jones (Reference Jones1973, Reference Jones1978) most likely intended to have this whole section as the middle part of the cherty bed (unit 4), which he described as 5.4 m of black, carbonaceous, and silicious mudstone layers with occasional indeterminate graptolite fragments. The result based on our collection identifies a prominent number of graptolites with distinctive elongated proximal ends such as Avitograptus cf. A. avitus (Davies, Reference Davies1929). Other graptolites occurring, but usually in lower numbers, include Hirsutograptus sp., Korenograptus bifurcus (Mu et al. in Nanjing Institute of Geology and Palaeontology, 1974), Neodiplograptus parajanus? (Štorch, Reference Štorch1983), Normalograptus aff. N. radicatus (Chen and Lin, Reference Chen and Lin1978), and Normalograptus normalis (Lapworth, Reference Lapworth1877) (Fig. 8). The occurrence of Neodiplograptus parajanus? (Štorch, Reference Štorch1983) and Hirsutograptus sp. are used to indicate the Akidograptus ascensus–Parakidograptus acuminatus Biozone, despite the lack of the eponymous index taxa. It should be noted, however, that our specimens of Neodiplograptus parajanus? (Štorch, Reference Štorch1983) are admittedly too poorly preserved for confident assignment of the LG7 or LG8 intervals. However, the spine-bearing species of the genus Hirsutograptus are only known from the Akidograptus ascensus–Parakidograptus acuminatus Biozone; hence, LG9 is definitely within the Silurian. The base of the biozone and hence the base of the Silurian System itself could be lower, but more complete specimens need to be recovered to ascertain this. The GSSP of the Silurian was defined at a point 1.6 m above the base of the Birkhill Shale Formation at Dob's Linn in the southern uplands of Scotland, which is marked by the first appearance of Akidograptus ascensus Davies, Reference Davies1929 and Parakidograptus praematurus (Davies) (Zalasiewicz et al., Reference Zalasiewicz, Taylor, Rushton, Loydell, Rickards and Williams2009; Melchin et al., Reference Melchin, Sadler, Cramer, Gradstein, Ogg, Schmitz and Ogg2020). The section can also be further subdivided into a lower Akidograptus ascensus Biozone and an upper Parak acuminatus Biozone (Melchin and Williams, Reference Melchin and Williams2000). Particular taxa that seem to be prevalent in this interval are biserial graptolites with distally forking or branching virgella, including Korenograptus bifurcus (Mu et al. in Nanjing Institute of Geology and Palaeontology, 1974), and Normalograptus aff. N. radicatus (Chen and Lin, Reference Chen and Lin1978) (which Maletz et al., Reference Maletz, Wang, Kai and Wang2021b regarded as the Normalograptus bifurcus/coremus/radicatus group). Korenograptus bifurcus (Mu et al. in Nanjing Institute of Geology and Palaeontology, 1974), however, occurs from the Akidograptus ascensus–Parakidograptus acuminatus Biozone in China (Chen et al., Reference Chen, Fan, Melchin and Mitchell2005a), Spain (Štorch et al., Reference Štorch, Bernal and Gutiérrez-Marco2019), and central Laos (Loydell et al., Reference Loydell, Udchachon and Burrett2019). Chen et al. (Reference Chen, Fan, Melchin and Mitchell2005a) noted Normalograptus radicatus (Chen and Lin, Reference Chen and Lin1978) as a junior synonym of Normalograptus coremus (Chen and Lin, Reference Chen and Lin1978); however, we consider them as separate species for now (see the remarks that follow for Normalograptus aff. N. radicatus). Neodiplograptus parajanus (Štorch, Reference Štorch1983) is also one of the key species commonly used to correlate within and between the Gondwanan and peri-Gondwanan regions (Štorch, Reference Štorch1996). Štorch (Reference Štorch1996) indicated the range of Neodiplograptus parajanus (Štorch, Reference Štorch1983) to be from the middle to upper Akidograptus ascensus–Parakidograptus acuminatus Biozone and coincidentally similar findings had been recorded in Jordan (Loydell, Reference Loydell2007) and Saudi Arabia (Williams et al., Reference Williams, Zalasiewicz, Boukhamsin and Cesari2016). As a result, it is possible to further constrain the current Langkawi interval to the middle–upper part of the Akidograptus ascensus–Parakidograptus acuminatus Biozone, at least until more specimens are recovered.

Figure 8. Graptolites of the Akidograptus ascensus–Parakidograptus acuminatus Biozone in Pulau Langgun, Langkawi. (1–4) Avitograptus cf. A. avitus (Davies, Reference Davies1929): (1) LG8_06; (2) LG8_01-1; (3) LG7_06-2; (4) LG8_03. (5, 6) Neodiplograptus parajanus? (Štorch, Reference Štorch1983): (5) LG7_08; (6) LG8_05. (7) Hirsutograptus sp., LG9_04-2. (8?, 9) Korenograptus bifurcus (Mu et al. in Nanjing Institute of Geology and Palaeontology, 1974): (8) LG8_01-2; (9) LG7_09-1. (10, 11) Normalograptus aff. N. radicatus (Chen and Lin, Reference Chen and Lin1978): (10) LG8_07, no virgella preserved; (11) LG7_06-1. (12) Normalograptus normalis (Lapworth, Reference Lapworth1877), LG9_02.
Graptolite significance
Chen et al. (Reference Chen, Jiayu, Mitchell, Harper, Junxuan, Renbin, Yuandong, Rongyu and Yi2000) noted that Metabolograptus persculptus specimens previously reported in the Langkawi Islands had not yet been described in detail and had to assume that the identification of said species is correct when making a correlation between the Ordovician–Silurian succession from the Yangtze region, South China, and the Malay Peninsula. Their analysis suggested that the section bearing the shelly Hirnantia fauna in Langkawi could be correlated with the upper Metabolograptus persculptus Biozone or above, which is slightly higher compared with a similar section from the Yangtze region. On restudying the Langkawi specimens, however, we conclude that the hitherto Metabolograptus persculptus species instead belong to several different graptolite species from the Metabolograptus extraordinarius Biozone, and thus the shelly Hirnantia fauna from the Langkawi Islands demonstrate a much closer association with the South Chinese section in terms of biostratigraphy. A revised graptolite biozonation is thus proposed for the latest Ordovician to the earliest Silurian succession of northwest Peninsular Malaysia (Fig. 9). Other studies had also recognized the similarity between the Sibumasu terrane and South China in the Upper Ordovician based on the fossils and sedimentary facies (Fortey and Cocks, Reference Fortey, Cocks, Hall and Holloway1998; Rong et al., Reference Rong, Harper, Huang, Li, Zhang and Chen2020). In that regard, we believe that new findings or at the very least revision of available graptolite specimens from the Sibumasu terrane (which include those from Peninsular Malaysia, Sumatra, West Thailand, and Myanmar) can be further used for biostratigraphical correlation with other parts of the world. Furthermore, the early Paleozoic is perhaps one of the most critical intervals where evidence of climate shift that lead to a dramatic impact on the biosphere can be observed. Sheets et al. (Reference Sheets, Mitchell, Melchin, Loxton, Štorch, Carlucci and Hawkins2016), for example, showed the dramatic changes in the distribution of planktonic graptolites going into the Late Ordovician mass extinction. A more detailed record of graptolite fossils together with high-resolution photographs (Figs. 10–13) could contribute to the paleontological database, which admittedly is rather lacking in the Southeast Asian region. Consequently, the data can be used to promote higher-resolution analysis of the biodiversity patterns and, together with the sedimentological record, give an insight into the marine conditions during the aforementioned period.

Figure 9. Graptolite biostratigraphy of the Ordovician–Silurian boundary in the northwestern domain of Peninsular Malaysia.

Figure 10. Photographs of the graptolites from the Metabolograptus extraordinarius Biozone in Pulau Langgun, Langkawi. (1, 2, 5, 6, 8) Metabolograptus ojsuensis (Koren’ and Mikhaylova in Koren et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980): (1) 191F-1_1; (2) 179F_1; (5) 188F_1; (6) 173F_1; (8) 176F_4. (3, 4, 7, 9, 12, 13) Metabolograptus extraordinarius (Sobolevskaya, Reference Sobolevskaya and Obut1974): (3) 174F_2; (4) 174F_3; (7) 174F_1; (9) 190F_6; (12) LG1_01; (13) LG1_04-1. (10, 11) Anticostia aff. A. lata (Elles and Wood, Reference Elles and Wood1906): (10) 179F_4; (11) 179F_3. Scale bars = 1 mm.

Figure 11. Photographs of the graptolites from the Metabolograptus extraordinarius Biozone in Pulau Langgun, Langkawi. (1, 2, 5) Korenograptus elegantulus (Mu and Ni, Reference Mu and Ni1983): (1) 182F_4; (2) 183F_3; (5) 172F_3. (3, 4) Neodiplograptus shanchongensis (Li, 1984): (3) 173F_5; (4) 180F_3. (6–8, 14) Normalograptus temalaensis (Jones, Reference Jones1973): (6) 189F_3; (7) 205F_2; (8) 187F_7; (14) 187F_3. (9–11) Normalograptus ajjeri (Legrand, Reference Legrand1977): (9) 187F_5; (10) 175F_1; (11) 180F_1. (12, 13) Normalograptus aff. N. mirnyensis (Obut and Sobolevskaya in Obut et al., Reference Obut, Sobolevskaya and Nikolaev1967): (12) 190F_3; (13) 185F_5. Scale bars = 1 mm.
Systematic paleontology
Suborder Axonophora Frech, Reference Frech1897
Infraorder Diplograptina Lapworth, Reference Lapworth1880
Family Diplograptidae Lapworth, Reference Lapworth1873
Genus Anticostia Stewart and Mitchell, Reference Stewart and Mitchell1997, emend. Štorch et al., Reference Štorch, Mitchell, Finney and Melchin2011
Type species
Anticostia macgregorae Stewart and Mitchell (Reference Stewart and Mitchell1997, p. 221, pl. 1, figs. 5A–M). From the limestone of the Vauréal Formation, Anticosti Island, Quebec, Canada, by original designation.
Anticostia aff. A. lata (Elles and Wood, Reference Elles and Wood1906)
Figures 4.8, 4.9, 7.9, 10.10, 10.11, 12.6, 12.7
- aff. *Reference Elles and Wood1906
Climacograptus latus Elles and Wood, p. 204, pl. 27, fig. 3a–h; text-fig. 135.
- aff. Reference Riva1988
Amplexograptus latus; Riva, p. 226, figs. 2a–h, 4.
- aff. Reference Štorch, Mitchell, Finney and Melchin2011
Anticostia lata; Štorch et al., p. 336, figs. 14G, O, Y, 19H–J, M, O, T, table 9 (see for further synonymy).
- aff. Reference Maletz, Wang and Wang2021a
Anticostia lata; Maletz et al., p. 170, fig. 5A, J.

Figure 12. Photographs of the graptolites from the Metabolograptus persculptus Biozone in Pulau Langgun, Langkawi. (1, 4) Metabolograptus persculptus (Elles and Wood, Reference Elles and Wood1907): (1) LG4L_01; (4) LG4L_04. (2, 8) Normalograptus angustus (Perner, Reference Perner1895): (2) LG4L_02; (8) LG4L_06. (3, 5) Normalograptus rhizinus (Li and Yang in Nanjing Institute of Geology and Mineral Resources, 1983): (3) LG4M_07-2; (5) LG4M_02. (6, 7) Anticostia aff. A. lata (Elles and Wood, Reference Elles and Wood1906), LG4M_07-1: (6) proximal half; (7) distal half. (9) Avitograptus? sp., LG4L_05. Scale bars = 1 mm.
Holotype
Figured by Elles and Wood (Reference Elles and Wood1906, pl. 27, fig. 3a) and selected by Riva (Reference Riva1988, fig. 2b), specimen SM A19680. From the upper Hartfell Shale, Dicellograptus anceps Biozone, Main Cliff, Dob's Linn, Scotland.
Material
Two specimens (179F_3 and 179F_4) previously collected by Jones below the shelly Hirnantia fauna bed and one poorly preserved specimen collected by the authors (LG4M_07-1) above the shelly Hirnantia fauna bed of the Tanjung Dendang Formation in Pulau Langgun.
Remarks
The present specimens have a similar thecal characteristic to Anticostia lata (Elles and Wood, Reference Elles and Wood1906) described by Riva (Reference Riva1988) and Štorch et al. (Reference Štorch, Mitchell, Finney and Melchin2011); however, it has a bigger overall measurement of DVW and could be instead a closely related species or subspecies of Anticostia lata (Elles and Wood, Reference Elles and Wood1906). The DVW is ~1.2–2.3 mm for the first three thecal pairs and reaches an observed maximum of 2.2–2.6 mm, which it seems to maintain throughout the preserved part of the tubarium. The 2TRD at the proximal section measures 1.4–1.85 mm while for the distal section it is 1.6–1.9 mm. The present specimens displayed a diplograptid-like proximal end furnished with a stout virgella, subapertural spine and anti-virgellar spine, which are the key features for this genus. Previously the species referred to as Amplexograptus latus (Riva, Reference Riva1988) was later redesignated as Anticostia lata (Štorch et al., Reference Štorch, Mitchell, Finney and Melchin2011). Riva (Reference Riva1988) also provided a detailed remark on the synonymy of this species when it was described under a different name in other parts of the world. Anticostia lata (Elles and Wood, Reference Elles and Wood1906) can be differentiated from the similar-looking Anticostia tenuissima (Ross and Berry, Reference Ross and Berry1963) by the former's strong thecal geniculation with thickened rim (Štorch et al., Reference Štorch, Mitchell, Finney and Melchin2011).
Infraorder Neograptina Štorch et al., Reference Štorch, Mitchell, Finney and Melchin2011
Paraphyletic Family Normalograptidae Štorch and Serpagli, Reference Štorch and Serpagli1993, emend. Melchin et al., Reference Melchin, Mitchell, Naczk-Cameron, Fan and Loxton2011
Genus Normalograptus Legrand, Reference Legrand1987, emend. Melchin et al., Reference Melchin, Mitchell, Naczk-Cameron, Fan and Loxton2011
Type species
Climacograptus scalaris var. normalis Lapworth (Reference Lapworth1877, p. 138, pl. 6, fig. 31). From the Llandovery of County Down, Ireland, by original designation.
Normalograptus ajjeri (Legrand, Reference Legrand1977)
Figures 5.10–5.12, 11.9–11.11
- *Reference Legrand1977
Climacograptus (Climacograptus) normalis ajjeri n. ssp., Legrand, p. 171, text-figs. 9A–D, 10A, B.
- Reference Loydell2007
Normalograptus ajjeri; Loydell, p. 29, text-figs. 12G, 15A–F, H, I, O (see for full list of synonymies and discussions).
- Reference Štorch, Mitchell, Finney and Melchin2011
Normalograptus ajjeri; Štorch et al., p. 368, figs. 25B–D, P, V, W, 26L, M, table 21.
- Reference Štorch and Schönlaub2012
Normalograptus ajjeri; Štorch and Schönlaub, p. 759, 3A, B, ?D, ?O, 4E, F.
- Reference Wang, Muir, Chen and Tang2015
Normalograptus ajjeri; Wang et al., p. 532, figs. 4B–D, F, O, 8H.
- Reference Chen, Chen, Aung and Muir2020
Normalograptus ajjeri; Chen et al., p. 50, figs. 4E–G, J, K, N, 8C, D, J, L, M, S.
Holotype
Designated by Legrand (Reference Legrand1977, p. 171), UL2223 d9. From Oued In Djerane, Algeria.
Material
Three moderately well-preserved specimens (175F_1, 180F_1, and 187F_5) were chosen for illustration and identification. Recovered below the shelly Hirnantia fauna bed of the Tanjung Dendang Formation in Pulau Langgun.
Remarks
The dimensions of the present materials match well with the description of Normalograptus ajjeri (Legrand, Reference Legrand1977) from Loydell (Reference Loydell2007). The DVW at the first thecal pair is 0.9–1.1 mm, at the second thecal pair is 1.1–1.2 mm, at the third thecal pair is 1.4 mm, at the fifth thecal pair is 1.45–1.5 mm, and after the fifth thecal pair reaches a maximum of 1.5–1.7 mm, which it maintains throughout the rest of the tubarium. The 2TRD measures 1.4–1.7 mm at the proximal part and 1.9–2.2 mm at the distal part. Elles and Wood (Reference Elles and Wood1906) distinguished Normalograptus normalis (Lapworth, Reference Lapworth1877) as specimens that have a DVW of no more than 1.5 mm, which has consequently been used repeatedly to diagnose Normalograptus normalis (Lapworth, Reference Lapworth1877) in much of the literature. Loydell (Reference Loydell2007), however, restudied the collection of Normalograptus normalis (Lapworth, Reference Lapworth1877), which had been previously figured by Elles and Wood, and noted that the specimens attained a DVW that is significantly greater than 1.5 mm; the holotype, for example, was recorded to instead be 1.85 mm in DVW. It is then agreed upon that the narrower form of Normalograptus normalis (Lapworth, Reference Lapworth1877) would henceforth be assigned to Normalograptus ajjeri (Legrand, Reference Legrand1977). Normalograptus ajjeri (Legrand, Reference Legrand1977) can be further distinguished from other similar normalograptids by having a tubarium that is narrower than that of Normalograptus medius (Törnquist, Reference Törnquist1897) and less tapering than that of Normalograptus transgradiens (Waern in Waern et al., Reference Waern, Thorslund, Henningsmoen and Säve-Söderbergh1948), a more densely spaced thecae compared with Normalograptus premedius (Waern in Waern et al., Reference Waern, Thorslund, Henningsmoen and Säve-Söderbergh1948), and a much bigger DVW compared with Normalograptus angustus (Perner, Reference Perner1895) and Normalograptus mirnyensis (Obut and Sobolevskaya in Obut et al., Reference Obut, Sobolevskaya and Nikolaev1967) (Štorch and Schönlaub, Reference Štorch and Schönlaub2012).
Normalograptus angustus (Perner, Reference Perner1895)
Figures 7.3, 7.5, 7.6, 12.2, 12.8
- *Reference Perner1895
Diplograptus (Glyptograptus) euglyphus Lapworth, var. angustus mihi Perner, p. 27, pl. 8, figs. 14a, b.
- Reference Přibyl1949
Climacograptus angustus; Přibyl, p. 7, pl. 2, figs. 2–9.
- Reference Štorch and Serpagli1993
Normalograptus angustus; Storch and Serpagli, p. 22, pl. 1, figs. 3, 4, ?6, pl. 2, figs. 2, 6, text-figs. 7A?, B, C, E, F.
- Reference Koren’ and Melchin2000
Normalograptus angustus; Koren’ and Melchin, p. 1097, figs. 4.8, 4.11–4.13, 5.4.
- Reference Chen, Fan, Melchin and Mitchell2005a
Normalograptus angustus; Chen et. al, p. 252, text-fig. 5D, I, K, Q, DD.
- Reference Loydell2007
Normalograptus angustus; Loydell, p. 30, pl. 1, fig. 3, text-fig. 15L–M (see for further synonymy).
- Reference Goldman, Mitchell, Melchin, Fan, Wu and Sheets2011
Normalograptus angustus; Goldman et al., p. 228, fig. 1K.
- Reference Štorch, Mitchell, Finney and Melchin2011
Normalograptus angustus; Štorch et al., p. 369, figs. 25G, Q, 26H.
- Reference Wang, Muir, Chen and Tang2015
Normalograptus angustus; Wang et al., p. 534, figs. 3S, 4I, 8G, J.
- Reference Chen, Chen, Aung and Muir2020
Normalograptus angustus; Chen et al., p. 52, figs. 4A–C, H, I, O, P, 7E, G, 8B, E–H, K, N–Q, 9Q, R.
Holotype
Figured by Perner (Reference Perner1895, pl. 8, fig. 14a, b), refigured and designated by Přibyl (Reference Přibyl1949, pl. 2, fig. 8), specimen NM L27552. From the Králův Dvůr Formation, Králův Dvůr Bohemia.
Material
Three specimens collected by the authors (LG4L_02, LG4L_06 and LG4M_06). Recovered above the shelly Hirnantia fauna bed of the Tanjung Dendang Formation in Pulau Langgun.
Remarks
The material described agrees well in overall dimension with Ordovician specimens of Perner (Reference Perner1895). DVW at the first thecal pair is 0.8–0.9 mm, at the second thecal pair is 1.0 mm, at the fifth thecal pair is ~1.1–1.2 mm, and subsequently is maintained at less than 1.3 mm throughout the tubarium (note that specimens LG4L_02 and LG4M_06 have a mineral vein running across the tubarium that would make the DVW slightly higher). The overall 2TRD is large, measuring 1.6–2.2 mm. Normalograptus angustus (Perner, Reference Perner1895) can be distinguished by its relatively narrow DVW, parallel-sided climacograptid thecae and a wide 2TRD. Štorch and Serpagli (Reference Štorch and Serpagli1993) noted that the Sardinian and Bohemian materials of early Silurian age are narrower compared with the latest Ordovician material of Perner (Reference Perner1895). Riva (Reference Riva1988) and many other authors considered Normalograptus miserabilis (Elles and Wood, Reference Elles and Wood1906) to be a junior synonym of Normalograptus angustus (Perner, Reference Perner1895); however, Normalograptus miserabilis (Elles and Wood, Reference Elles and Wood1906) was referred to the genus Styracograptus after Goldman et al. (Reference Goldman, Mitchell, Melchin, Fan, Wu and Sheets2011) restudied specimens of Normalograptus angustus (Perner, Reference Perner1895), including materials from southeast China (Chen et al., Reference Chen, Jiayu, Mitchell, Harper, Junxuan, Renbin, Yuandong, Rongyu and Yi2000, Reference Chen, Fan, Melchin and Mitchell2005a) along with the type material of Styracograptus miserabilis (Elles and Wood) and concluded that they have different internal structures. Normalograptus angustus (Perner, Reference Perner1895) can be differentiated from other similarly parallel-sided normalograptids such as Normalograptus mirnyensis (Obut and Sobolevskaya in Obut et al., Reference Obut, Sobolevskaya and Nikolaev1967) by having more widely spaced thecae and from Normalograptus ajjeri (Legrand, Reference Legrand1977) by having a much narrower overall DVW (Chen et al., Reference Chen, Jiayu, Mitchell, Harper, Junxuan, Renbin, Yuandong, Rongyu and Yi2000, Reference Chen, Fan, Melchin and Mitchell2005a; Koren’ and Melchin, Reference Koren’ and Melchin2000; Loydell, Reference Loydell2007; Štorch et al., Reference Štorch, Mitchell, Finney and Melchin2011; Wang et al., Reference Wang, Muir, Chen and Tang2015).
Normalograptus aff. N. mirnyensis (Obut and Sobolevskaya in Obut et al., Reference Obut, Sobolevskaya and Nikolaev1967)
Figures 5.6–5.9, 11.12, 11.13
- aff. *Reference Obut, Sobolevskaya and Nikolaev1967
Hedrograptus mirnyensis Obut and Sobolevskaya in Obut et al., p. 47, pl. 1, figs. 4–9.
- aff. Reference Koren’ and Melchin2000
Normalograptus mirnyensis new combination; Koren’ and Melchin, p. 1099, figs. 5.10, 5.17, 7.12, 7.13, 8.1–8.5.
- aff. Reference Chen, Fan, Melchin and Mitchell2005a
Normalograptus mirnyensis; Chen et al., p. 262, text-fig. 8B, F, G, I, L–N.
- aff. Reference Štorch, Mitchell, Finney and Melchin2011
Normalograptus mirnyensis; Štorch et al., p. 374, figs. 25E, F, I, J, 26C, F, K, table 24.
- aff. Reference Wang, Muir, Chen and Tang2015
Normalograptus mirnyensis; Wang et al., p. 536, figs. 4A, E, G, H, J, L, M, R, X, 7E, G, 8A, C.
- aff. Reference Chen, Chen, Aung and Muir2020
Normalograptus mirnyensis; Chen et al., p. 52, figs. 4L, 10M.
Holotype
Original designation previously figured by Obut et al. (Reference Obut, Sobolevskaya and Nikolaev1967, pl. 1, fig.4). From the Akidograptus ascensus–Parakidograptus acuminatus Biozone of Mirny Creek, northeast Russia.
Material
Two specimens previously collected by Jones (185F_5 and 190F_3) and two specimens collected by the authors (LG1_05 and LG1_06-2). Recovered below the shelly Hirnantia fauna bed of the Tanjung Dendang Formation in Pulau Langgun.
Remarks
The present material resembles Normalograptus mirnyensis (Obut and Sobolevskaya in Obut et al., Reference Obut, Sobolevskaya and Nikolaev1967) specimens described by Chen et al. (Reference Chen, Fan, Melchin and Mitchell2005a), Štorch et al. (Reference Štorch, Mitchell, Finney and Melchin2011), and Wang et al. (Reference Wang, Muir, Chen and Tang2015); however, it possesses a slightly wider DVW and 2TRD overall and might be related to said species. The DVW at the first thecal pair is 0.8–1.0 mm, at the third thecal pair is 1.1 mm, at the fifth thecal pair ~1.15–1.25 mm, and distally reaches an average of less than 1.3 mm, which it maintains throughout the whole tubarium. The 2TRD is relatively dense; at the proximal part it measures 1.1–1.5 mm, and at the distal part it is 1.3–1.7 mm. Normalograptus mirnyensis (Obut and Sobolevskaya in Obut et al., Reference Obut, Sobolevskaya and Nikolaev1967) is typically characterized by its relatively narrow DVW (normally ~1.1 mm or less in maximum width), parallel-sided supragenicular walls, and climacograptid thecae with strong geniculation and dense 2TRD. Depending on the preservation, however, the thecae might look slightly glyptograptid. It has a more densely spaced thecae than Normalograptus angustus (Perner, Reference Perner1895) and a markedly narrower tubarium than Normalograptus ajjeri (Legrand, Reference Legrand1977) (Chen et al., Reference Chen, Fan, Melchin and Mitchell2005a, Reference Chen, Chen, Aung and Muir2020; Štorch et al., Reference Štorch, Mitchell, Finney and Melchin2011; Štorch and Schönlaub, Reference Štorch and Schönlaub2012; Wang et al., Reference Wang, Muir, Chen and Tang2015). It also differs by having no genicular flanges compared with Normalograptus jideliensis (Koren’ and Mikhaylova in Koren’ et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980) and Normalograptus acceptus (Koren’ and Mikhaylova in Koren’ et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980) (Koren’ and Melchin, Reference Koren’ and Melchin2000). One specimen (190F_3), which had been described as the juvenile paratype of Glyptograptus temalaensis Jones, Reference Jones1973, was previously recorded with a maximum DVW of 1.6 mm (Jones, Reference Jones1973, specimen 190F); however, restudy reveals a maximum DVW of 1.3 mm, and it is hereby reassigned to Normalograptus aff. N. mirnyensis (Obut and Sobolevskaya in Obut et al., Reference Obut, Sobolevskaya and Nikolaev1967).
Normalograptus normalis (Lapworth, Reference Lapworth1877)
Figures 8.12, 13.9
- *Reference Lapworth1877
Climacograptus scalaris Hisinger (Non. Linnaeus.) Var. b. normalis Lapworth, Lapworth, p. 138, pl. 6, fig. 31.
- Reference Blackett, Zalasiewicz, Zalasiewicz and Rushton2007
Normalograptus normalis; Blackett and Zalasiewicz.
- Reference Loydell2007
Normalograptus normalis; Loydell, p. 38, pl. 1, figs. 4, 7, text-figs. 12E, F, 16C, H, N, 19 (see for full list of synonymies, including specimens formerly assigned to N. normalis and further discussions).

Figure 13. Photographs of the graptolites from the Akidograptus ascensus–Parakidograptus acuminatus Biozone in Pulau Langgun, Langkawi. (1–3) Avitograptus cf. A. avitus (Davies, Reference Davies1929): (1) LG8_01-1; (2) LG8_03; (3) LG7_06-2. (4, 5) Neodiplograptus parajanus? (Štorch, Reference Štorch1983): (4) LG7_08; (5) LG8_05. (6) Hirsutograptus sp., LG9_04-2. (7) Korenograptus bifurcus (Mu et al. in Nanjing Institute of Geology and Palaeontology, 1974), LG7_09-1. (8) Normalograptus aff. N. radicatus (Chen and Lin, Reference Chen and Lin1978), LG7_06-1. (9) Normalograptus normalis (Lapworth, Reference Lapworth1877), LG9_02. Scale bars = 1 mm.
Holotype
Figured by Elles and Wood (Reference Elles and Wood1906, pl. 26, fig. 2a), Williams (Reference Williams1983, text-fig. 4a), specimen BU 1136. From the Parakidograptus acuminatus s.l. Biozone of Dob's Linn, southern Scotland.
Material
One specimen collected by the authors (LG9_02). From the LG9 bed (equivalent to Jones's cherty bed) of the Tanjung Dendang Formation in Pulau Langgun.
Remarks
The present specimen fully agrees with the measurements of the holotype of Normalograptus normalis (Lapworth, Reference Lapworth1877) refigured in the Atlas of Graptolite Type Specimens Folio 2 (no. 2.61) and restudied by Loydell (Reference Loydell2007) together with his Jordanian material. The DVW at the first thecal pair is 1.0 mm, at the second thecal pair is 1.1 mm, at the third thecal pair is 1.3 mm, at the fifth thecal pair is 1.5 mm, and at the tenth thecal pair is 1.9 mm, which is the maximum DVW recorded. The 2TRD measures 1.45–2.00 mm at the proximal part and 1.8–2.3 mm distally. Note that the holotype of Normalograptus normalis (Lapworth, Reference Lapworth1877) reaches a maximum DVW of 1.85 mm; hence, all instances of Normalograptus normalis (Lapworth, Reference Lapworth1877) in literature measuring less than 1.5–1.6 mm can instead be assigned to Normalograptus ajjeri (Legrand, Reference Legrand1977). Previous non-synonymies for Normalograptus normalis (Lapworth, Reference Lapworth1877) are noted by Loydell (Reference Loydell2007) for reference. Jones (Reference Jones1968) also described a somewhat narrow form of Normalograptus normalis (Lapworth, Reference Lapworth1877) from the Cystograptus vesiculosus to Coronograptus cyphus biozones in Langkawi and from the Coronograptus cyphus to Coronograptus gregarius biozones in central Kedah, with a DVW measuring 1.0–1.5 mm, which could be instead assigned to Normalograptus ajjeri (Legrand, Reference Legrand1977). Do note, however, that one illustrated specimen from Langkawi (Jones, Reference Jones1968, pl. 7b, specimen 97F) shows a slightly protracted proximal end and is preserved in a somewhat subscalariform view. Normalograptus normalis (Lapworth, Reference Lapworth1877) can be distinguished from Normalograptus medius (Törnquist, Reference Törnquist1897) from the latter having a characteristically rounded proximal end while specimens with much narrower thecal apertures can instead be assigned to Normalograptus scalaris (Hisinger, Reference Hisinger1837) (Loydell, Reference Loydell2007).
Normalograptus aff. N. radicatus (Chen and Lin, Reference Chen and Lin1978)
Figures 8.10, 8.11, 13.8
- aff. *Reference Chen and Lin1978
Climacograptus radicatus (Chen and Lin, Reference Chen and Lin1978), p. 31, pl. 5, 12–19.
- aff. ?Reference Chen1984
Climacograptus radicatus; Chen, p. 253, table A1.
- aff. Reference Loxton2017
Normalograptus? radicatus; Loxton, p. 259, pl. 47.13?, 50.6–50.8, 55.11?; figs. 41.1–41.4.
Holotype
Chen and Lin (Reference Chen and Lin1978, pl. 5.14), specimen number 35960a. From the Akidograptus ascensus–Climacograptus bicaudatus Biozone of Tongzi, Northern Guizhou.
Material
Two specimens were collected by the authors (LG7_06-1 and LG8_07). Specimen LG8_07 does not have its virgella preserved. From the LG7 and LG8 bed (equivalent to Jones's cherty bed) of the Tanjung Dendang Formation in Pulau Langgun.
Remarks
The specimens are most likely related to the Normalograptus bifurcus/coremus/radicatus group, which can be characterized by the distal branching of the virgella (Maletz et al., Reference Maletz, Wang, Kai and Wang2021b). The DVW at the first thecal pair is 1.00–1.05 mm, at the second thecal pair is 1.10–1.15 mm, at the third thecal pair is 1.25–1.30 mm, at the fifth thecal pair is 1.4–1.5 mm, and afterward maintains a DVW of 1.60–1.65 mm throughout the whole tubarium. The 2TRD is large, 1.7–1.9 mm at the proximal part and 1.95–2.40 mm at the distal part. Chen et al. (Reference Chen, Fan, Melchin and Mitchell2005a) regarded Normalograptus radicatus (Chen and Lin, Reference Chen and Lin1978) as a junior synonym of Normalograptus coremus (Chen and Lin, Reference Chen and Lin1978), suggesting that the differences between the two would be a result of preservation (the former are said to be preserved in a subscalariform to scalariform view). Further inspection of the holotype of Normalograptus radicatus (Chen and Lin, Reference Chen and Lin1978) and the best syntype of Normalograptus coremus (Chen and Lin, Reference Chen and Lin1978), however, reveals significant differences that present throughout the tubarium: (1) the proximal section of Normalograptus coremus (Chen and Lin, Reference Chen and Lin1978) starts relatively narrow (DVW = 0.6 mm) but increases rapidly compared with the blunt end of Normalograptus radicatus (Chen and Lin, Reference Chen and Lin1978), which starts with a DVW of 0.95 mm; (2) the maximum distal DVW for Normalograptus coremus (Chen and Lin, Reference Chen and Lin1978) is fairly large, reaching ~2.1 mm, while Normalograptus radicatus (Chen and Lin, Reference Chen and Lin1978) reaches a maximum DVW of less than 1.6 mm throughout the tubarium; (3) Normalograptus radicatus (Chen and Lin, Reference Chen and Lin1978) maintains climacograptid-like thecae throughout the tubarium while in Normalograptus coremus (Chen and Lin, Reference Chen and Lin1978), the thecae become progressively less geniculate distally; (4) 2TRD for Normalograptus radicatus (Chen and Lin, Reference Chen and Lin1978) ranged between 1.6 and 1.9 mm while Normalograptus coremus (Chen and Lin, Reference Chen and Lin1978) have a consistently higher 2TRD of more than 2.1 mm throughout; (5) the thecae of Normalograptus radicatus (Chen and Lin, Reference Chen and Lin1978) alternate for the whole of the tubarium while in Normalograptus coremus (Chen and Lin, Reference Chen and Lin1978), the thecae initially alternate in the proximal section but become more opposite to each other in the distal section. In any case, both share a similar long virgella that forked into several branches at ~2.7 mm down the turning point of the first theca. By comparison, our specimen has a significantly shorter virgella that forked into two after 1 mm down. We would consider these graptolites as a separate species for now; however, more collection and a detailed study would help to further enhance our understanding of this group. Another very similar graptolite is Normalograptus minor (Huang, Reference Huang1982), which also features the same branching virgella. Chen et al. (Reference Chen, Fan, Melchin and Mitchell2005a) described Normalograptus minor (Huang, Reference Huang1982) to have a significantly narrower DVW compared with Normalograptus radicatus (Chen and Lin, Reference Chen and Lin1978) and Normalograptus coremus (Chen and Lin, Reference Chen and Lin1978), which can also be seen in Loxton (Reference Loxton2017), where the maximum DVW measures ~1.2–1.4 mm. Other authors, however, described Normalograptus minor (Huang, Reference Huang1982) with a maximum DVW of 1.5–1.6 mm such as in Štorch et al. (Reference Štorch, Bernal and Gutiérrez-Marco2019). Loxton (Reference Loxton2017) further differentiated Normalograptus minor (Huang, Reference Huang1982) from Normalograptus radicatus (Chen and Lin, Reference Chen and Lin1978) on the basis of its smaller proximal 2TRD, which is less than 1.75 mm. Normalograptus radicatus (Chen and Lin, Reference Chen and Lin1978) can be distinguished from Normalograptus rhizinus (Li and Yang in Nanjing Institute of Geology and Mineral Resources, 1983) from the latter having wider thecal spacing and virgella that tend to expand distally into a vane-like structure (Chen et al., Reference Chen, Fan, Melchin and Mitchell2005a). The present specimen also resembles very closely Normalograptus ajjeri (Legrand, Reference Legrand1977) if the former virgella is not preserved, with both having the same DVW and thecal characteristic; however, our specimen of Normalograptus aff. N. radicatus (Chen and Lin, Reference Chen and Lin1978) has a slightly bigger 2TRD compared with the Hirnantian Normalograptus ajjeri (Legrand, Reference Legrand1977) specimens.
Normalograptus rhizinus (Li and Yang in Nanjing Institute of Geology and Mineral Resources, 1983)
Figures 7.7, 7.8, 12.3, 12.5
- *1983
Climacograptus rhizinus Li and Yang in Nanjing Institute of Geology and Mineral Resources, p. 454, pl. 162, figs. 13, 14.
- Reference Li1999
Climacograptus rhizinus; Li, pl. 4, fig. 8.
- Reference Chen, Fan, Melchin and Mitchell2005a
Normalograptus rhizinus; Chen et al., p. 268, text-fig. 9I, M, P, V–X.
- Reference Chen, Rong, Fan, Zhan, Mitchell, Harper, Melchin, Peng, Finney and Wang2006
Normalograptus rhizinus; Chen et al., p. 191, fig. 9.2.
- Reference Loxton2017
Normalograptus? rhizinus; Loxton, p. 264, pl. 32.1–32.3, 32.6, 32.7, figs. 28.1, 28.2, 28.10, 29.14.
- Reference Loydell, Udchachon and Burrett2019
Normalograptus rhizinus; Loydell et al., p. 370, fig. 6A.
- Reference Štorch, Bernal and Gutiérrez-Marco2019
Normalograptus rhizinus; Štorch et al., p. 7, figs. 3n, q, 7b.
- Reference Maletz, Wang, Kai and Wang2021b
Normalograptus rhizinus; Maletz et al., fig. 4k.
Holotype
Designated by Li and Yang in Nanjing Institute of Geology and Mineral Resources (1983, pl. 162. fig. 13), refigured by Li (Reference Li1999, pl. 4, fig. 8), specimen number 54127. From the Parakidograptus acuminatus Biozone of Southern Anhui, China.
Material
Two specimens collected by the authors (LGM4M_02 and LG4M_07-2). Recovered above the shelly Hirnantia fauna bed of the Tanjung Dendang Formation in Pulau Langgun.
Remarks
The present specimens are similar to material recorded elsewhere such as in China (Chen et al., Reference Chen, Fan, Melchin and Mitchell2005a, Reference Chen, Rong, Fan, Zhan, Mitchell, Harper, Melchin, Peng, Finney and Wang2006; Maletz et al., Reference Maletz, Wang, Kai and Wang2021b), Spain (Štorch et al., Reference Štorch, Bernal and Gutiérrez-Marco2019), and central Laos (Loydell et al., Reference Loydell, Udchachon and Burrett2019). The DVW at the first thecal pair is 0.95–1.00 mm, at the second thecal pair is 1.15–1.20 mm, at the third thecal pair is 1.20–1.25 mm, and at the fifth thecal pair is ~1.40–1.45 mm; distally it reaches a maximum of 1.50–1.65 mm. The 2TRD is relatively large, measuring 1.85–2.20 mm. Chen et al. (Reference Chen, Fan, Melchin and Mitchell2005a) noted the distal width variation and suggested that Normalograptus rhizinus (Li and Yang in Nanjing Institute of Geology and Mineral Resources, 1983) includes both Climacograptus rhizinus and Climacograptus acus (Jin et al., Reference Jin, Ye, He, Wan, Wang, Zhao, Li, Xu and Zhang1982). The type specimen of Climacograptus acus, which has unfortunately gotten lost, is reported to reach a maximum DVW of 1.7 mm. Specimens illustrated by Chen et al. (Reference Chen, Fan, Melchin and Mitchell2005a) and Loydell et al., (Reference Loydell, Udchachon and Burrett2019, fig. 6A) possess a virgella that either expands abruptly into a broad vane or expands more gradually. The holotype of Normalograptus rhizinus (Li and Yang in Nanjing Institute of Geology and Mineral Resources, 1983), for example, has a much more prominent bulb-like vane. The presence of an anti-virgellar spine can be seen in specimens illustrated by Chen et al. (Reference Chen, Rong, Fan, Zhan, Mitchell, Harper, Melchin, Peng, Finney and Wang2006, p. 191, fig. 9.2) and Loxton (Reference Loxton2017); however, from the latter, several specimens of Normalograptus? rhizinus recorded feature multiple spines. Normalograptus rhizinus (Li and Yang in Nanjing Institute of Geology and Mineral Resources, 1983) can be differentiated from a very similar Avitograptus avitus (Davies, Reference Davies1929) in the width of the tubarium and in having a long, distally widening virgella. Avitograptus avitus (Davies, Reference Davies1929) is also noted to commonly possess a relatively elongate first thecal pair (Chen et al., Reference Chen, Fan, Melchin and Mitchell2005a). Normalograptus rhizinus (Li and Yang in Nanjing Institute of Geology and Mineral Resources, 1983) are also similar to Akidograptus ascensus Davies, Reference Davies1929 in DVW and general thecal characteristic; however, the genus Akidograptus is generally characterized by th11 that turns up significantly high up the sicular aperture, giving a more protracted proximal end.
Normalograptus temalaensis (Jones, Reference Jones1973)
Figures 5.13–5.17, 11.6–11.8, 11.14
- *Reference Jones1973
Glyptograptus temalaensis Jones, p. 18, pl. 1, fig. 3a, c.
- ?Reference Li1984a
Glyptograptus temalaensis; Li, p. 322, pl. 2, figs. 8, ?9–11.
- Reference Zhen, Zhang, Harper, Zhan, Fang, Wang, Yu and Li2020
Diplogarptus temalaensis (Jones); Zhen et al., p. 384, fig. 12C.
Holotype
Designated by Jones (Reference Jones1973, pl.1, fig. 3a). The specimen on slab numbered 180F, recovered from Bed 1, equivalent to the Metabolograptus persculptus Biozone interval in the Tanjung Dendang Formation in Pulau Langgun, Langkawi. However, the original holotype cannot be identified from the present slab. A new lectotype, specimen 205F_2, as well as two new paratypes, specimens 187F_7 and 189F_3, have been chosen.
Material
Five specimens previously collected by Jones (172F_2, 187F_3, 187F_7, 189F_3, and 205F_2) were picked for illustration and identification. Originally recorded from the Metabolograptus persculptus and Coronograptus gregarius biozones of the Tanjung Dendang Formation in Pulau Langgun. However, the record from the latter biozone seems to be ambiguous upon reexamination of Jones's graptolite collection.
Remarks
The DVW at the first thecal pair is 0.9–1.2 mm, at the second thecal pair is 1.10–1.25 mm, at the third thecal pair is 1.3–1.5 mm, at the fifth thecal pair is 1.4–1.7 mm, at the tenth thecal pair is 1.7–1.8 mm, and after the tenth thecal pair reaches a maximum of 1.80–2.05 mm, which it maintains throughout the rest of the tubarium. The 2TRD increases from 1.5–1.8 mm at the proximal part to 1.7–2.1 mm at the distal part of the tubarium. Jones (Reference Jones1973) noted that the diagnostic features of Normalograptus temalaensis (Jones, Reference Jones1973) are its medium-sized tubarium (maximum DVW ~1.8–2.0 mm), rather blunt proximal end, and thecae that are widely spaced. In addition, there is a description of how the thecae show a considerable axial torsion, but these features could instead be a preservational artifact, also acknowledged by Štorch et al. (Reference Štorch, Mitchell, Finney and Melchin2011). The original holotype (Jones, Reference Jones1973, pl.1, fig. 3a), which lacked the proximal termination, cannot be identified from Jones's original collection (specimen 180F_1 seems to resemble the holotype but has a different proximal section, smaller length, and slightly smaller DVW). We decided to select a lectotype of Normalograptus temalaensis (Jones, Reference Jones1973), which is specimen 205F_2, for future reference. The paratypes designated by Jones are either missing (for example, specimens that were recorded to be higher in the succession) or reassigned to another species (the smaller paratype regarded as a juvenile specimen, upon reexamination, was assigned to Normalograptus aff. N. mirnyensis [Obut and Sobolevskaya in Obut et al., Reference Obut, Sobolevskaya and Nikolaev1967]), and consequently, two new paratypes, specimens 187F_7 and 189F_3, are selected. The thecal form of Normalograptus temalaensis (Jones, Reference Jones1973) is somewhere between climacograptid and glyptograptid types and can be further diagnosed from the supragenicular walls that are slightly inclined to no more than 10°, the infragenicular wall that is inclined at a low angle, and the moderately deep excavation overall. Admittedly, Normalograptus temalaensis (Jones, Reference Jones1973) is difficult to distinguish from the similar-looking Normalograptus normalis (Lapworth, Reference Lapworth1877), Normalograptus ajjeri (Legrand, Reference Legrand1977), and Normalograptus elegantulus (Mu and Ni, Reference Mu and Ni1983). Normalograptus temalaensis (Jones, Reference Jones1973) possesses similar parallel-sided tubarium as Normalograptus normalis (Lapworth, Reference Lapworth1877) and Normalograptus ajjeri (Legrand, Reference Legrand1977); however, Normalograptus temalaensis (Jones, Reference Jones1973) seems to have slightly more inclined supragenicular walls and, overall, a bigger DVW compared with Normalograptus ajjeri (Legrand, Reference Legrand1977) (Normalograptus ajjeri [Legrand, Reference Legrand1977] usually attain a maximum DVW of significantly less than 1.8 mm). Normalograptus temalaensis is somewhat similar to Normalograptus ex. gr. N. normalis illustrated by Underwood et al. (Reference Underwood, Deynoux and Ghienne1998, fig. 5C) and Normalograptus normalis (Lapworth, Reference Lapworth1877) in Chen et al. (Reference Chen, Fan, Melchin and Mitchell2005a, p. 264) in having a distinct blunt proximal end. Specimens identified as Normalograptus temalaensis (Jones, Reference Jones1973) from South Anhui, China (illustrated by Li, Reference Li1984a), had been regarded as a junior synonym of Normalograptus elegantulus (Mu and Ni, Reference Mu and Ni1983); however, the latter specimen bears a distinctive glyptograptid thecae with much more inclined supragenicular walls throughout the whole tubarium (Štorch et al., Reference Štorch, Mitchell, Finney and Melchin2011). Normalograptus temalaensis (Jones, Reference Jones1973) also have a smaller maximum DVW compared with Metabolograptus extraordinarius (Sobolevskaya, Reference Sobolevskaya and Obut1974) (which could reach more than 2.5 mm), Metabolograptus ojsuensis (Koren’ and Mikhaylova in Koren et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980), and Metabolograptus persculptus (Elles and Wood, Reference Elles and Wood1907) (where the latter two also have more inclined glyptograptid thecae distally).
Family incertae sedis
Genus Hirsutograptus Koren’ and Rickards, Reference Koren’ and Rickards1996, emend. Melchin et al., Reference Melchin, Mitchell, Naczk-Cameron, Fan and Loxton2011
Type species
Hirsutograptus longispinosus Koren’ and Rickards (Reference Koren’ and Rickards1996, p. 40, pl. 7, fig.4, text-fig. 8A, F–G) from the Kos-Istek region, western Kazakhstan, by original designation.
Hirsutograptus sp.
Figures 8.7, 13.6
Material
One poorly preserved specimen collected by the authors (LG9_04-2). From the LG9 bed (equivalent to Jones's cherty bed) of the Tanjung Dendang Formation in Pulau Langgun.
Remarks
Melchin et al. (Reference Melchin, Mitchell, Naczk-Cameron, Fan and Loxton2011) in their emended diagnosis described the genus Hirsutograptus as having a proximal pattern H astogeny with climacograptid thecae that bear laterally and ventrally directed genicular hoods, genicular spines, or both. The present specimen is too poorly preserved to completely discern its actual species and can be identified only up to the genus level by the presence of the climacograptid thecae with apparent spinosity. The proximal DVW is ~1.25 mm. Mesially it maintains a DVW of 1.7 mm; however, distally it may reach to a maximum of 2.0 mm depending on preservation. The 2TRD measures ~1.85–2.40 mm. Similar species include Hirsutograptus longispinosus (Koren’ and Rickards, Reference Koren’ and Rickards1996, pl. 7, fig. 4; text-fig. 8A, F–G), with the genicula having one mesial spine each and a maximum DVW of 1.7 mm.
Superfamily Monograptoidea Lapworth, Reference Lapworth1873, emend. Melchin et al., Reference Melchin, Mitchell, Naczk-Cameron, Fan and Loxton2011
Paraphyletic Family Dimorphograptidae Elles and Wood, Reference Elles and Wood1908, emend. Melchin et al., Reference Melchin, Mitchell, Naczk-Cameron, Fan and Loxton2011
Genus Avitograptus Melchin et al., Reference Melchin, Mitchell, Naczk-Cameron, Fan and Loxton2011
Type species
Glyptograptus(?) avitus Davies (Reference Davies1929, p. 8, fig. 21). From Dob's Linn, southern Scotland, UK, by original designation.
Avitograptus cf. A. avitus (Davies, Reference Davies1929)
Figures 8.1–8.4, 13.1–13.3
- cf. *Reference Davies1929
Glyptograptus? avitus Davies, p. 8, fig. 21.
- Reference Packham1962
Glyptograptus? avitus; Packham, text-fig. 7a.
- cf. Reference Williams1983
Glyptograptus? avitus; Williams, p. 625, pl. 66, figs. 8–10; text-figs. 7h–l, 9 a–c, ?d, 10a–c.
- non Reference Chen, Fan, Melchin and Mitchell2005a
Normalograptus avitus (Davies, Reference Davies1929); Chen et al., p. 255, pl. 1, fig. 11; pl. 2, fig. 9; text-fig. 7D, G, J–L.
- cf. Reference Melchin, Mitchell, Naczk-Cameron, Fan and Loxton2011
Avitograptus avitus (Davies, Reference Davies1929); Melchin et al., p. 292, fig. 6B.
- cf. Reference Loxton2017
Avitograptus avitus (Davies, Reference Davies1929); Loxton, pl. 31.25, 32.4, 32.5, 32.8, 32.9, 37.24, 38.1; figs. 27.13, 27.17, 28.7, 29.9, 29.10.
- cf. Reference Muir, Zhang, Botting and Ma2020
Avitograptus avitus (Davies, Reference Davies1929); Muir et al., p. 6, figs. 3.2, 4.4.
Holotype
Figured by Davies (Reference Davies1929) and Packham (Reference Packham1962); later refigured by Williams (Reference Williams1983, text-fig. 7l), specimen SM A10019 stored in the Sedgwick Museum, Cambridge. From the Parakidograptus acuminatus? Biozone of the Birkhill Shale, Dob's Linn, Scotland.
Material
Four specimens were collected by the authors (LG7_06-2, LG8_01-1, LG8_03, and LG8_06). From the LG7 and LG8 beds (equivalent to Jones's cherty bed) of the Tanjung Dendang Formation in Pulau Langgun.
Remarks
Avitograptus avitus (Davies, Reference Davies1929) is the type species used by Melchin et al. (Reference Melchin, Mitchell, Naczk-Cameron, Fan and Loxton2011) to describe the genus Avitograptus. The diagnosis for the genus includes specimens with pattern J astogeny, with th11 turning at the level of the sicular aperture, slightly elongated first thecal pair (usually length of th11 at least 1.4 times the length of the free ventral wall for th21), geniculate thecae with slight to moderately inclined supragenicular walls and a full median septum (Melchin et al., Reference Melchin, Mitchell, Naczk-Cameron, Fan and Loxton2011, p. 295). Note that the point of upturn of th11 itself seems to vary within the population. Williams (Reference Williams1983) made a detailed redescription of the type species of Avitograptus avitus (Davies, Reference Davies1929) and diagnosed the species as having a maximum DVW of 1.5 mm, straight to sloping supragenicular walls, and virgella commonly bifurcating close to the tubarium. The Langkawi specimens, however, differ in having a bigger DVW compared with the type specimen of Avitograptus avitus (Davies, Reference Davies1929) and as such are referred to Avitograptus cf. A. avitus (Davies, Reference Davies1929). The DVW measured at the first thecal pair is 1.0–1.1 mm, at the second thecal pair is 1.05–1.20 mm, at the third thecal pair is 1.10–1.45 mm, at the fifth thecal pair is 1.50–1.55 mm, and distally maintains a maximum of ~1.7–1.8 mm. The 2TRD is conspicuously large, measuring 1.9–2.5 mm. Some authors recorded a maximum DVW higher than the type specimen; for example, Chen et al. (Reference Chen, Fan, Melchin and Mitchell2005a) described Avitograptus avitus (Davies, Reference Davies1929) with a maximum DVW of 1.5–1.8 mm, which Loxton (Reference Loxton2017) suggested to be incorrectly assigned due to their great distal width and the lack of protracted proximal thecae. Meanwhile, Wang et al. (Reference Wang, Muir, Chen and Tang2015) described specimens of Avitograptus sp. aff. A. avitus with a distal maximum width of 1.6–1.7 mm while Chen et al. (Reference Chen, Chen, Aung and Muir2020) described one specimen of Avitograptus cf. A. avitus (Davies, Reference Davies1929) that measures 1.8 mm at the ninth thecal pairs (figs. 5G, 9O); however, the proximal section seems to be slightly curved and would be better assigned to Avitograptus acanthocystus (Fang et al., Reference Fang, Liang, Zhang and Yu1990). Avitograptus avitus (Davies, Reference Davies1929) can be differentiated from the similar looking Avitograptus akidomorphus (Muir et al., Reference Muir, Zhang, Botting and Ma2020) by the latter having more pronounced thecal geniculation and smaller DVW (max DVW = 1.2 mm) overall (Muir et al., Reference Muir, Zhang, Botting and Ma2020). Normalograptus rhizinus (Li and Yang in Nanjing Institute of Geology and Mineral Resources, 1983) and Metabolograptus wangjiawanensis (Mu and Lin, Reference Mu and Lin1984), however, can be distinguished from Avitograptus avitus (Davies, Reference Davies1929) by its width and its long, spatulate virgella (Chen et al., Reference Chen, Fan, Melchin and Mitchell2005a).
Avitograptus? sp.
Figures 7.1, 12.9
Material
One specimen collected by the authors (LG4L_05). Recovered above the shelly Hirnantia fauna bed of the Tanjung Dendang Formation in Pulau Langgun.
Remarks
This specimen is questionably assigned to the genus Avitograptus, which can be identified by the possible pattern J astogeny with the first theca that normally grows down to a point at or above the sicular aperture and then turns upward (Melchin et al., Reference Melchin, Mitchell, Naczk-Cameron, Fan and Loxton2011; Muir et al., Reference Muir, Zhang, Botting and Ma2020). The DVW is 0.98 mm at the first pair, 1.07 mm at the second pair, 1.25 mm at the third pair, and 1.35 mm at the fourth pair. The 2TRD measures 1.85–2.00 mm. The current specimen, however, seems to have an abnormal proximal growth, with the sicula on the side of where th11 grows, resulting in a shorter upward growing portion of th11. Another possibility is that it could be a transitional species between Normalograptus and Akidograptus (Melchin et al., Reference Melchin, Mitchell, Naczk-Cameron, Fan and Loxton2011), but more specimens need to be collected to ascertain this idea.
Superfamily Retiolitoidea Lapworth, Reference Lapworth1873, emend. Melchin et al., Reference Melchin, Mitchell, Naczk-Cameron, Fan and Loxton2011
Paraphyletic Family Neodiplograptidae Melchin et al., Reference Melchin, Mitchell, Naczk-Cameron, Fan and Loxton2011
Genus Korenograptus Melchin et al., Reference Melchin, Mitchell, Naczk-Cameron, Fan and Loxton2011
Type species
Glyptograptus gnomus Churkin and Carter, Reference Churkin and Carter1970 (p. 24, pl. 2, fig. 16, text-fig. 11E). From southeastern Alaska, USA, by original designation.
Korenograptus bifurcus (Mu et al. in Nanjing Institute of Geology and Palaeontology, 1974)
Figures 8.8(?), 8.9, 13.7
- *1974
Diplograptus bifurcus Mu et al. in Nanjing Institute of Geology and Palaeontology, p. 212, pl. 98, fig. 12.
- Reference Chen and Lin1978
Diplograptus bifurcus; Chen and Lin, p. 22, pl. 2.3, 2.4, fig. 4a.
- Reference Štorch and Schönlaub2012
Rickardsograptus? bifurcus (Ye, Reference Ye1978); Štorch and Schönlaub, p. 762, figs. 3L–N, 4B–D, K.
- Reference Loxton2017
Korenograptus bifurcus (Ye, Reference Ye1978); Loxton, p. 311, pl. 45.13; fig. 42.1, 42.8.
- Reference Loydell, Udchachon and Burrett2019
Neodiplograptus bifurcus; Loydell et al., p. 370, fig. 6B.
- Reference Štorch, Bernal and Gutiérrez-Marco2019
Korenograptus bifurcus (Mu et al. in Nanjing Institute of Geology and Palaeontology, 1974); Štorch et al., p. 16, figs. 7c, k, 8g, ?j, 10a.
Holotype
Figured by Mu et al. in Nanjing Institute of Geology and Palaeontology (1974, p. 212, pl. 98, fig. 12) and by Chen and Lin (Reference Chen and Lin1978, p. 22, pl. 2.3, 2.4, fig. 4a.), specimen no. 21430.
Material
Poorly preserved specimens collected by the authors (LG7_09-1, questionably LG8_01-2). From the LG7 and possibly LG8 bed (equivalent to Jones's cherty bed) of the Tanjung Dendang Formation in Pulau Langgun.
Remarks
Korenograptus bifurcus (Mu et al. in Nanjing Institute of Geology and Palaeontology, 1974) can be easily recognized from its long, distally bifurcating virgella that split into two prominent branches. The DVW of the Langkawi specimens is 0.9 mm at the first thecal pair, 1.1 mm at the second thecal pair, and 1.8 mm at the fifth thecal pair; the maximum recorded DVW is ~2.2 mm at the eighth thecal pair. Most authors recorded a distal DVW of more than 2.0 mm for Korenograptus bifurcus (Mu et al. in Nanjing Institute of Geology and Palaeontology, 1974); for example, in specimens studied by Štorch and Schönlaub (Reference Štorch and Schönlaub2012), a DVW of 2.30–2.55 mm is achieved at the tenth thecal pair, and in Štorch et al. (Reference Štorch, Bernal and Gutiérrez-Marco2019) a DVW of 2.40–2.65 mm is recorded by the tenth–fifteenth thecal pairs. Štorch and Schönlaub (Reference Štorch and Schönlaub2012) previously differentiated the Ye (Reference Ye1978) species from the species described by Mu et al. in Nanjing Institute of Geology and Palaeontology (1974) by the former having more densely spaced thecae and a more robust proximal end. On further reexamination of the Chinese materials, Štorch et al. (Reference Štorch, Bernal and Gutiérrez-Marco2019) suggested that the Ye (Reference Ye1978) species, along with several other similar taxa, was conspecific with the Mu et al. (in Nanjing Institute of Geology and Palaeontology (1974) species. Štorch and Schönlaub (Reference Štorch and Schönlaub2012) referred the species to the genus Rickardsograptus; however, Loxton (Reference Loxton2017) and Štorch et al. (Reference Štorch, Bernal and Gutiérrez-Marco2019) instead assigned it to the genus Korenograptus, the former highlighting the lack of tapered proximal end and dramatic change in mesial thecal form, and the latter remarking on the pattern H astogeny and a largely glyptograptid thecae that lack any genicular thickening. Our present specimen unfortunately displays a relatively poorly preserved proximal end. Korenograptus bifurcus (Mu et al. in Nanjing Institute of Geology and Palaeontology, 1974) is very similar to Korenograptus bicaudatus (Chen and Lin, Reference Chen and Lin1978); however, the latter has a virgella that bifurcates only very slightly below the turning point of the first theca, usually less than 1 mm (Chen and Lin, Reference Chen and Lin1978; Loxton, Reference Loxton2017; Loydell et al., Reference Loydell, Udchachon and Burrett2019; Štorch et al., Reference Štorch, Bernal and Gutiérrez-Marco2019).
Korenograptus elegantulus (Mu and Ni, Reference Mu and Ni1983)
Figures 5.3–5.5, 11.1, 11.2, 11.5
- *Reference Mu and Ni1983
Glyptograptus elegantulus Mu and Ni, Reference Mu and Ni1983, p. 161, pl. 7, fig. 4, pl. 8, figs. 1, 2, 10–13, text-fig. 3b, d.
- Reference Štorch, Mitchell, Finney and Melchin2011
Normalograptus elegantulus (Mu and Ni, Reference Mu and Ni1983); Štorch et al., p. 370, figs. 25K, R, X–Z, 26A, B, D, I, J, 27J–M, table 22.
- Reference Maletz, Wang, Kai and Wang2021b
Korenograptus elegantulus (Mu and Ni, Reference Mu and Ni1983); Maletz et al., fig. 5a, c.
Holotype
Figured by Mu and Ni (Reference Mu and Ni1983, pl. 4, fig. 13), specimen no. 67970. Listed to be from the Diplograptus bohemicus Biozone, Riajue, Tibet.
Material
Three moderately preserved specimens (172F_3, 182F_4, and 183F_3) chosen for illustration and identification. Recovered below the shelly Hirnantia fauna bed of the Tanjung Dendang Formation in Pulau Langgun.
Remarks
The graptolite description matches well with Korenograptus elegantulus (Mu and Ni, Reference Mu and Ni1983) (Mu and Ni, Reference Mu and Ni1983; Štorch et al., Reference Štorch, Mitchell, Finney and Melchin2011). The DVW is 0.9–1.0 mm at the first thecal pair, 1.05–1.10 mm at the second thecal part, 1.2–1.3 mm at the third thecal pair, 1.5 mm at the fifth thecal pair, 1.65 mm at the tenth thecal pair, and reaches a maximum of 1.5–1.8 mm (distal part is mostly broken). Compared with Normalograptus temalaensis (Jones, Reference Jones1973), Korenograptus elegantulus (Mu and Ni, Reference Mu and Ni1983) are less parallel-sided and possess a much more inclined glyptograptid thecae. Korenograptus elegantulus (Mu and Ni, Reference Mu and Ni1983) have a more asymmetrical proximal end, slightly smaller DVW, and wider spaced thecae distally compared with the similar Normalograptus lungmaensis (Štorch et al., Reference Štorch, Mitchell, Finney and Melchin2011). Korenograptus laciniosus (Churkin and Carter, Reference Churkin and Carter1970) is another species that seems to be related to Korenograptus elegantulus (Mu and Ni, Reference Mu and Ni1983) but can be differentiated by a shorter sicula, a narrower DVW, and closer spaced thecae (Churkin and Carter, Reference Churkin and Carter1970; Chen et al., Reference Chen, Fan, Melchin and Mitchell2005a; Štorch et al., Reference Štorch, Mitchell, Finney and Melchin2011; Maletz et al., Reference Maletz, Wang, Kai and Wang2021b). Loxton (Reference Loxton2017) measured both the specimen of Korenograptus laciniosus (Churkin and Carter, Reference Churkin and Carter1970) from Blackstone River, Yukon, and South China (as previously described by Chen et al., Reference Chen, Fan, Melchin and Mitchell2005a) and noted that for most cases, the specimens reach a consistent distal width of or very close to 1.5 mm.
Genus Metabolograptus Obut and Sennikov, Reference Obut, Sennikov, Betechtuna and Zhuravleva1985, emend. Melchin et al., Reference Melchin, Mitchell, Naczk-Cameron, Fan and Loxton2011
Type species
Diplograptus modestus sibericus Obut (Reference Obut and Nikiforova1955, p. 138). From the Siberian Platform, Russia, by original designation.
Metabolograptus extraordinarius (Sobolevskaya, Reference Sobolevskaya and Obut1974)
Figures 4.10–4.20, 10.3, 10.4, 10.7, 10.9, 10.12, 10.13
- vpReference Jones1968
Glyptograptus persculptus (Salter, Reference Salter1873); Jones, p. 436 (specimen not figured).
- *Reference Sobolevskaya and Obut1974
Fenhsiangograptus extraordinarius Sobolevskaya, p. 69, pl. 3, figs. 6, 7, 25A, H, M, N, S–U, 27I, N–R, 28A, B, F, J, K, table 23.
- Reference Chen, Fan, Melchin and Mitchell2005a
Normalograptus extraordinarius (Sobolevskaya, Reference Sobolevskaya and Obut1974); Chen et al., p. 256, pl. 1, figs. 1, 2; pl. 2, fig. 12; text-figs. 6C, 7H, I, 8W (see for further synonymy).
- Reference Štorch, Mitchell, Finney and Melchin2011
Normalograptus extraordinarius (Sobolevskaya, Reference Sobolevskaya and Obut1974); Štorch et al., p. 372, figs. 25A, H, M, N, S–U, 27I, N–R, 28A, B, F, J, K, table 23 (see for further synonymy).
- Reference Chen, Chen, Aung and Muir2020
Metabolograptus extraordinarius (Sobolevskaya, Reference Sobolevskaya and Obut1974); Chen et al., p. 61, figs. 5Q, 6H, 7I, 9J, T, 11A.
Holotype
Figured by Sobolevskaya (Reference Sobolevskaya and Obut1974, p1. 3, fig. 6), specimen no. 602x/1, from the Ina River area of the Kolyma Basin, eastern Siberia.
Material
Eleven specimens figured; six specimens were previously collected by Jones (173F_3, 174F_1, 174F_2, 174F_3, 187F_6, and 190F_6), and five specimens were collected by the authors (LG1_01, LG1_02, LG1_03, LG1_04-1, and LG1_04-2). Recovered below the shelly Hirnantia fauna bed of the Tanjung Dendang Formation in Pulau Langgun. Specimens are preserved in mostly poor to moderately good condition.
Remarks
One of the most notable characteristics for Metabolograptus extraordinarius (Sobolevskaya, Reference Sobolevskaya and Obut1974) is its significantly wide maximum DVW. The DVW of the Langkawi specimens increases fairly rapidly, from 0.90–1.55 mm at the first thecal pair, to 1.15–1.75 mm at the second thecal pair, to 1.3–1.9 mm at the third thecal pair, to 1.50–2.05 mm at the fifth thecal pair, to a maximum of 2.25–3.20 mm usually attained after the tenth to twentieth thecal pair. Occasional DVW reduction can be observed in the mesial and/or distal section before returning to its maximum. The 2TRD measures ~1.2–2.2 mm on the proximal part and 1.4–2.4 mm on the distal part of the tubarium. Štorch et al. (Reference Štorch, Mitchell, Finney and Melchin2011) noted how papers describing Metabolograptus extraordinarius (Sobolevskaya, Reference Sobolevskaya and Obut1974) from around the world display a degree of morphological variety between and within the population. For example, the maximum DVW of specimens from Vinini Creek, USA, varies from 2.1 to 3.0 mm (Štorch et al., Reference Štorch, Mitchell, Finney and Melchin2011), from the Upper Yangtze region of China it measures 2.9–3.5 mm (Chen et al., Reference Chen, Fan, Melchin and Mitchell2005a), while in the Mandalay region of Myanmar a maximum of 3.6 mm is recorded (Chen et al., Reference Chen, Chen, Aung and Muir2020). The DVW of Metabolograptus extraordinarius (Sobolevskaya, Reference Sobolevskaya and Obut1974) from the Langkawi seems to be between the narrower Vinini Creek and the larger Upper Yangtze specimens, but all other parameters seem to match. A very similar graptolite, Neodiplograptus incommodus Legrand, Reference Legrand2009, was described to have a maximum DVW between 2.4 and 2.7 (sometimes 2.9 mm), and the thecae tend to become more glyptograptid in the medial and distal parts of the tubarium (Legrand, Reference Legrand2009). We consider that Neodiplograptus incommodus Legrand, Reference Legrand2009 might be instead a subspecies or at least the narrow variety of Metabolograptus extraordinarius (Sobolevskaya, Reference Sobolevskaya and Obut1974). So far, the only record of Neodiplograptus incommodus Legrand, Reference Legrand2009 is in the Algerian Sahara (Legrand, Reference Legrand2009) and the Mandalay region, Myanmar (Chen et al., Reference Chen, Chen, Aung and Muir2020), but further examination of the available specimens of Metabolograptus extraordinarius (Sobolevskaya, Reference Sobolevskaya and Obut1974) in other regions could potentially delineate these two graptolites. Metabolograptus extraordinarius (Sobolevskaya, Reference Sobolevskaya and Obut1974) can be distinguished from Metabolograptus ojsuensis (Koren’ and Mikhaylova in Koren et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980) and Metabolograptus persculptus (Elles and Wood, Reference Elles and Wood1907) by having a much wider tubarium overall, less protracted proximal end, and considerably more geniculate and overlapping thecae with relatively smaller apertural excavations and shorter, less inclined supragenicular walls (Štorch et al., Reference Štorch, Mitchell, Finney and Melchin2011).
Metabolograptus ojsuensis (Koren’ and Mikhaylova in Koren’ et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980)
Figures 4.1–4.7, 10.1, 10.2, 10.5, 10.6, 10.8
- vpReference Jones1968
Glyptograptus persculptus (Salter, Reference Salter1873); Jones, p. 436, pl. 2, fig. 6a, b.
- *Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980
Glyptograptus? ojsuensis Koren’ and Mikhaylova in Koren' et al., p. 143, pl. 41, figs. 1–8, pl. 42, figs. 1, 2; text-fig. 43.
- Reference Chen, Fan, Melchin and Mitchell2005a
Normalograptus ojsuensis (Koren’ and Mikhaylova in Koren’ et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980); Chen et al., p. 264, pl. 1, figs. 4, 7, pl. 2, fig. 8, text-figs. 9D, K, N, Q, 10I (see for the list of synonymous species Diplograptus bohemicus in China).
- Reference Štorch, Mitchell, Finney and Melchin2011
Normalograptus ojsuensis (Koren’ and Mikhaylova in Koren’ et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980); Štorch et al., p. 375, figs. 27A–D, 28G–I, table 25.
- Reference Chen, Chen, Aung and Muir2020
Metabolograptus ojsuensis (Koren’ and Mikhaylova in Koren et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980); Chen et al., p. 62, figs. 5K, L, 6G, 7A, C, N, 8A, I, 10I–K, 11C.
Holotype
Designated and figured by Koren’ and Mikhaylova in Koren’ et al. (1980, pl. 41, fig. 1), mature specimen 11586/96 housed in the CNIGR Museum, Saint Petersburg, Russia. From Ojsu Spring, Kazakhstan.
Material
Seven figured specimens (173F_1, 175F_3, 176F_2, 176F_4, 179F_1, 188F_1, and 191F-1_1). Recovered below the shelly Hirnantia fauna bed of the Tanjung Dendang Formation in Pulau Langgun. Specimens are preserved mostly in moderately good condition.
Remarks
The present material agrees well with specimens of Metabolograptus ojsuensis (Koren’ and Mikhaylova in Koren et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980) described by several authors such as Chen et al. (Reference Chen, Fan, Melchin and Mitchell2005a, Reference Chen, Chen, Aung and Muir2020) and Štorch et al. (Reference Štorch, Mitchell, Finney and Melchin2011). The DVW of the Langkawi specimens at the first thecal pair is 0.9–1.1 mm, at the second thecal pair is 1.15–1.30 mm, at the third thecal pair is 1.2–1.4 mm, at the fifth thecal pair is 1.3–1.7 mm, at the tenth thecal pair is 1.70–1.85 mm, and mostly after the tenth thecal pair reaches a maximum of 1.85–2.55 mm, which it maintains throughout the rest of the tubarium. The 2TRD measures 1.3–1.7 mm at the proximal part and 1.4–1.9 mm at the distal part of the tubarium. Štorch et al. (Reference Štorch, Mitchell, Finney and Melchin2011) distinguished Metabolograptus ojsuensis (Koren’ and Mikhaylova in Koren et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980) from a very similar Metabolograptus persculptus (Elles and Wood, Reference Elles and Wood1907) by the DVW of the proximal end and how fast it reaches the same maximum DVW. Metabolograptus ojsuensis (Koren’ and Mikhaylova in Koren et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980) is noted to have a narrower proximal end (first thecal pair of 0.9–1.1 mm) and reaches a maximum distal DVW of ~1.65–2.30 mm well above the tenth thecal pair, compared with Metabolograptus persculptus (Elles and Wood, Reference Elles and Wood1907), which has a slightly bigger DVW proximally (first thecal pair of 1.2–1.4 mm) and reaches the same maximum DVW by the sixth to ninth thecal pairs. On the basis of these quantifiable features, we reassigned the specimen of Metabolograptus persculptus (Salter, Reference Salter, Huxley and Etheridge1865) previously identified by Jones (Reference Jones1968, pl. 2, fig. 6a, b) to Metabolograptus ojsuensis (Koren’ and Mikhaylova in Koren et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980). Furthermore, Metabolograptus ojsuensis (Koren’ and Mikhaylova in Koren et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980) is also described as having more strongly geniculate thecae proximally and relatively taller apertural excavations (Štorch et al., Reference Štorch, Mitchell, Finney and Melchin2011). From Metabolograptus extraordinarius (Sobolevskaya, Reference Sobolevskaya and Obut1974), Metabolograptus ojsuensis (Koren’ and Mikhaylova in Koren et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980) can be differentiated by having a markedly narrower DVW, a more protracted proximal end, and considerably less geniculate and less overlapping thecae with relatively broad excavations and longer, slightly more inclined supragenicular walls (Štorch et al., Reference Štorch, Mitchell, Finney and Melchin2011).
Metabolograptus persculptus (Elles and Wood, Reference Elles and Wood1907)
Figures 7.2, 7.4, 12.1, 12.4
- Reference Salter, Huxley and Etheridge1865
Diplograptus persculptus (Salter?), Salter, p. 25 (see Strachan, Reference Strachan1971).
- *Reference Elles and Wood1907
Diplograptus (Glyptograptus) persculptus Salter; Elles and Wood, p. 257, pl. 31, fig. 7a–c, text-fig. 176a, b.
- Reference Williams1983
Glyptograptus persculptus (Salter, Reference Salter, Huxley and Etheridge1865); Williams, p. 622, pl. 66, figs. 1–3.
- Reference Štorch and Loydell1996
Normalograptus persculptus (Elles and Wood); Štorch and Loydell, p. 872, text-figs. 3–5 (see for further synonymy).
- Reference Chen, Fan, Melchin and Mitchell2005a
Normalograptus persculptus (Elles and Wood, Reference Elles and Wood1907); Chen et al., p. 266, text-figs. 5B, 9A, F, L (see for further synonymy).
- Reference Štorch, Mitchell, Finney and Melchin2011
Normalograptus persculptus (Elles and Wood, Reference Elles and Wood1907); Štorch et al., p. 376, figs. 25L, AA, 27E–H, 28C–E, table 26.
Material
Two specimens collected by the authors (LG4L_01 and LG4L_04). Recovered above the shelly Hirnantia fauna bed of the Tanjung Dendang Formation in Pulau Langgun.
Remarks
The current specimens are assigned to Metabolograptus persculptus (Elles and Wood, Reference Elles and Wood1907) on the basis of the diagnoses of the species by Williams (Reference Williams1983) and Štorch and Loydell (Reference Štorch and Loydell1996). The measured DVW at the first thecal pair is 0.95–1.05 mm, at the second thecal pair is 1.05–1.10 mm, at the third thecal pair is 1.1–1.2 mm, at the fifth thecal pair is 1.4–1.5 mm, at the tenth thecal pair is ~1.8 mm, and reaches a maximum of 2.25 mm after the tenth–eleventh thecal pair in a single specimen with preserved distal part. The 2TRD increases from 1.6–2.0 mm at the proximal part of the tubarium to 1.95–2.20 mm at the distal part. Metabolograptus persculptus (Elles and Wood, Reference Elles and Wood1907) is very similar to Metabolograptus ojsuensis (Koren’ and Mikhaylova in Koren’ et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980), and it is often difficult to discern these two species. Chen et al. (Reference Chen, Fan, Melchin and Mitchell2005a) recognized that not all specimens in China previously assigned to Diplograptus bohemicus (Marek, Reference Marek1955) are synonymous with Metabolograptus persculptus (Elles and Wood, Reference Elles and Wood1907) (Koren et al. in Koren’ et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980; Štorch and Loydell, Reference Štorch and Loydell1996) and instead consider all occurrences of Diplograptus bohemicus (Marek, Reference Marek1955) that either underlie or occur within the strata representing the main phase of glacioeustatic regression to be Metabolograptus ojsuensis (Koren’ and Mikhaylova in Koren’ et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980). The current specimens from Pulau Langgun were found directly above the shelly Hirnantia fauna bed and have a higher 2TRD compared with Metabolograptus ojsuensis (Koren’ and Mikhaylova in Koren’ et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980) specimens found below the shelly bed (Metabolograptus ojsuensis identified does not have 2TRD reaching more than 2 mm even in the distal section). The same differences can also be observed for the specimens from the Vinini Creek section (Štorch et al., Reference Štorch, Mitchell, Finney and Melchin2011), where the 2TRD of Metabolograptus ojsuensis (Koren’ and Mikhaylova in Koren’ et al., Reference Koren’, Mikhaylova, Tzai, Apollonov, Nikitin and Bandaletov1980) very rarely reached more than 2.0 mm except for the very distal thecae, while Metabolograptus persculptus (Elles and Wood, Reference Elles and Wood1907) reached the 2TRD of 2.0 mm more consistently even in the proximal section.
Genus Neodiplograptus Legrand, Reference Legrand1987, emend. Melchin, Reference Melchin1998
Type species
Diplograptus magnus Lapworth (Reference Lapworth1900, p. 132, fig. 21a–d). From the Llandovery of central Wales, UK, by original designation.
Neodiplograptus parajanus? (Štorch, Reference Štorch1983)
Figures 8.5, 8.6, 13.4, 13.5
- *Reference Štorch1983
Diplograptus parajanus Štorch, p. 168, pl. 4, figs. 1–3, text-fig. 3A, B.
- Reference Štorch and Serpagli1993
Neodiplograptus parajanus; Štorch and Serpagli, p. 19, pl.3, figs. 1, 5, 6, text-fig. 5A, F, G.
- Reference Loydell2007
Neodiplograptus parajanus; Loydell, p. 24, text-figs. 12C, D, 14A, B, E.
Holotype
Figured by Štorch (Reference Štorch1983, pl. 4, fig. 2, text-fig. 3A), number PŠ 54b. From the Parakidograptus acuminatus Biozone of Želkovice Formation, Praha-Řepy, Bohemia.
Material
Two specimens collected by the authors (LG7_08 and LG8_05). From the LG7 and LG8 bed (equivalent to Jones's cherty bed) of the Tanjung Dendang Formation in Pulau Langgun.
Remarks
The present specimens are preserved only up to the sixth and eighth thecal pairs. The DVW increases rapidly from 1.3–1.4 mm at the first thecal pair to 1.6–1.7 mm at the second thecal pair and from 1.65 mm at the third thecal pair to 2.0 mm at the fifth thecal pair. A recorded DVW maximum is 2.35 mm. The 2TRD measures 1.7–2.0 mm. The proximal morphologies somewhat agree with Neodiplograptus parajanus (Štorch, Reference Štorch1983) specimens described and illustrated by Štorch and Serpagli (Reference Štorch and Serpagli1993) and Loydell (Reference Loydell2007). Štorch and Serpagli (Reference Štorch and Serpagli1993) noted that the proximal thecae in some of their Sardinian specimens are glyptograptid (which can be observed in the Langkawi specimens) while those in the more mature tubarium tend to be more climacograptid in appearance. The distal thecae, however, are glyptograptid to orthograptid. Another discernible difference is that the Langkawi specimens do not have the long virgella that is usually recorded in Neodiplograptus parajanus (Štorch, Reference Štorch1983). Neodiplograptus parajanus (Štorch, Reference Štorch1983) can be differentiated from other similar Neodiplograptids by its tapering but distinctly wider proximal end and more orthograptid distal thecae (Štorch and Serpagli, Reference Štorch and Serpagli1993). Specimens of Neodiplograptus lanceolatus Štorch and Serpagli, Reference Štorch and Serpagli1993, which can occur together with those of Neodiplograptus parajanus (Štorch, Reference Štorch1983), have a shallower thecal excavation, a more orthograptid thecal appearance, and a distinct distal narrowing in more mature specimens. Loydell (Reference Loydell2007) remarked that some of his Jordanian specimens are similar to “Orthograptus” regularis Fang et al., Reference Fang, Liang, Zhang and Yu1990; however, specimens of the latter species differ in having a smaller 2TRD and a broader nema.
Neodiplograptus shanchongensis (Li, 1984)
Figures 5.1, 5.2, 11.3, 11.4
- *Reference Li1984b
Diplograptus shanchongensis Li, p. 344, pl. 13, figs. 9–11.
- Reference Chen, Fan, Melchin and Mitchell2005a
Neodiplograptus shanchongensis; Chen et al., p. 274, text-fig. 11B–E.
- Reference Štorch, Bernal and Gutiérrez-Marco2019
Neodiplograptus shanchongensis; Štorch et al., p. 17, figs. 7p, x, 10g, o.
Lectotype
Not yet designated. Li's (Reference Li1984b) material from the Kaochiapien Formation, Jiangxian, Anhui, China.
Material
Two figured specimens previously collected by Jones (173F_5 and 180F_3). Recovered below the shelly Hirnantia fauna bed of the Tanjung Dendang Formation in Pulau Langgun.
Remarks
Chen et al. (Reference Chen, Fan, Melchin and Mitchell2005a) noted that Neodiplograptus shanchongensis (Li, 1984) is distinguished from other Neodiplograptids of the Neodiplograptus modestus group by its close thecal spacing and tendency to slightly narrow distally. Jones's specimens are shorter compared with specimens described by Chen et al. (Reference Chen, Fan, Melchin and Mitchell2005a), which grow up to 18 mm long; as such, the distal trend is harder to compare. The DVW increases rapidly from 1.3–1.4 mm at the first thecal pair to 1.6–1.7 mm at the second thecal pair, 1.65 mm at the third pair, and 2.0 mm at the fifth pair to the maximum of 2.35 mm. The 2TRD measures 1.7–2.0 mm. Neodiplograptus shanchongensis (Li, 1984) has a smaller tubarium width overall compared with the similarly shaped Neodiplograptus modestus (Lapworth), which reaches a maximum of 3.0–3.3 mm at th 10 and is narrower distally with significantly closer thecal spacing compared with Neodiplograptus charis (Mu and Ni, Reference Mu and Ni1983) (Chen et al., Reference Chen, Fan, Melchin and Mitchell2005a). Neodiplograptus shanchongensis (Li, 1984) is suggested to be the stratigraphical precursor of the similar Neodiplograptus lanceolatus Štorch and Serpagli, Reference Štorch and Serpagli1993 (Štorch et al., Reference Štorch, Bernal and Gutiérrez-Marco2019). It is described by Štorch and Serpagli (Reference Štorch and Serpagli1993) to reach a higher maximum width of commonly 3.0–3.5 mm attained at the level of ninth–eleventh thecal pair; in addition, Neodiplograptus shanchongensis (Li, 1984) does not seem to form the diagnostic lanceolate shape of Neodiplograptus lanceolatus Štorch and Serpagli, Reference Štorch and Serpagli1993 when it narrows distally.
Acknowledgments
This research was made possible thanks to the generous funds provided by the Yayasan UTP (YUTP-FRG) research grant. Special thanks to M. Melchin for providing the camera lucida drawings of the holotype of Normalograptus radicatus (Chen and Lin, Reference Chen and Lin1978) and the best syntype of Normalograptus coremus (Chen and Lin, Reference Chen and Lin1978). A huge thanks to J. Maletz for his valuable comments on the systematic palaeontology of the graptolites. We would also like to give our utmost thanks to the reviewers including M. Melchin and an anonymous reviewer, and the editorial office of the Journal of Paleontology including E. Currano, P. Štorch and J. Kastigar for their tremendous help and insightful feedback that helped to further improve the manuscript.
Declaration of competing interests
The authors declare none.













