



INTRODUCTION
In September 2020, during excavations at the Volchia Griva (VG) site in Western Siberia, Russia, in the lower bone-bearing level dated to ca. 20–18 14C ka BP, the broken-off distal end of a mammoth femur was found containing bone fragments and teeth of foxes. All these remains form an artifactual bone assemblage (ABA). There are no analogues for such a find either for the period of the last glacial maximum (LGM) or for the entire Paleolithic of Siberia. This confirms the uniqueness of VG, which was discovered in 1957 (Polunin, Reference Polunin1961) and studied in the 1960s through early 1990s (Alexeeva and Vereshchagin, Reference Alexeeva and Vereshchagin1970; Okladnikov et al., Reference Okladnikov, Grigorenko, Alexeeva and Volkov1971; Maschenko and Leshchinskiy, Reference Maschenko, Leshchinskiy and Podobina2001; Zenin, Reference Zenin2002; Leshchinskiy et al., Reference Leshchinskiy, Kuzmin, Zenin and Jull2008), as a paleontological, geological, and archaeological site in Northern Eurasia.
This publication follows a series of articles based on a new stage of research at VG since 2015 (Leshchinskiy et al., Reference Leshchinskiy, Zenin, Burkanova, Dudko, Gulina, Fedyaev, Semiryakov and Kanishcheva2015). The results of seven fieldwork seasons indicate that VG today is the largest woolly mammoth locality in Asia, which also contains many Late Paleolithic artifacts. Excavations on an area >550 m2 since the 1950s, numerous test pits, manual exploratory drilling, inspection of house cellars and water pipe trenches, and data from local residents allowed us to determine a total of several hectares of space where mammoth faunal remains were found.
The bone-bearing horizon of VG, which is represented by a series of large lens-like strata that forms three levels in some places, contains tens of thousands of bones and teeth belonging to woolly mammoth, horse, bison, wolf, fox, rodents, and other mammals. The maximum concentration of only mammoth remains in some places exceeds 130 items/m2 (Leshchinskiy and Burkanova, Reference Leshchinskiy and Burkanova2022). Lithic artifacts are found throughout the site, but their concentration is usually low (<1 specimen/m2; Leshchinskiy et al., Reference Leshchinskiy, Zenin and Bukharova2021). Nevertheless, the 2020–2021 excavation pits exposed a marginal activity area of the Paleolithic site with a thick ash lens. In some parts of these new excavation pits the highest concentration of cultural materials (>1400 items/m2) known at VG was detected. Artifacts are usually small (>50% up to 1 cm in size). There is also a large amount of faunal remains (average concentration >200 items/m2), mainly in the form of fragments and whole bones, and teeth of the polar fox. In this paper, we present these data for the first time, including ABA.
REGIONAL SETTING AND GEOLOGICAL CONTEXT
The VG site is situated in the eastern part of the Baraba Lowland, 220 km north of the border of Russia and Kazakhstan. The mammal remains are found in the northeastern part of the eponymous (Volchia Griva) sloping mound, elongated along an azimuth of ~50–55°. This mound is ~11 km long and ~0.5–1 km wide, with a maximum altitude of ~149 m asl and a relative elevation of 10–15 m above the surrounding grounds. The spatial distribution of the currently known faunal and cultural materials is located within Mamontovoe Village, Kargat County, Novosibirsk Province (Fig. 1).
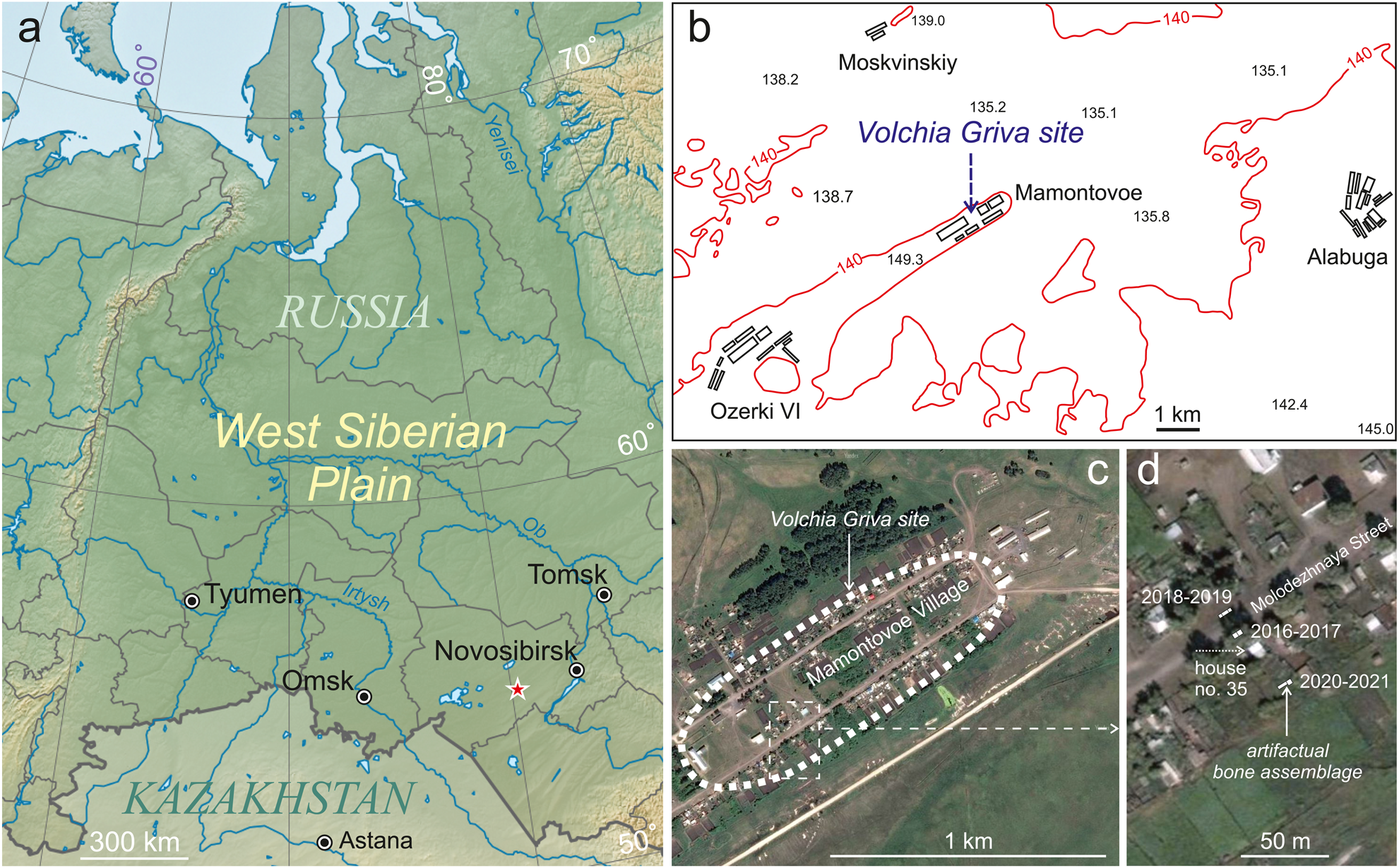
Figure 1. Location of the Volchia Griva. (a) Administrative map of the West Siberian Plain (the star indicates the site); (b) county map with local altitudes in meters; (c) the Volchia Griva site within Mamontovoe Village (satellite image); (d) location of 2016–2021 excavation pits with the position of the ABA.
Currently, the Baraba Lowland is located in the forest-steppe zone of the Ob-Irtysh interfluve. Open landscapes, unchanged by human activities, are covered with steppe halophytic-grass meadows. In forest outliers, birch dominates; willow and aspen are also found. The modern climate is markedly continental. According to observations from the middle of the twentieth century, the average air temperatures are: for July +18.3–19.7°С, and for January −19.1°С to −18.2 °С. The average annual temperature is +0.5–1.2°С, with a difference between minimum and maximum annual temperatures of ~80–95°С. The annual precipitation is 315–450 mm (Arhiv Klimaticheskih Dannyh, 2012; Pogoda i Klimat, 2020).
The Baraba Lowland is a large depression covering an area of ~117,000 km2. It is filled with predominantly loose sandy–clayey sediments, with a thickness of 500–2800 m or more. Sediment age is continuously traced from Jurassic to Quaternary (Babin et al., Reference Babin, Chernykh, Golovina, Zhigalov, Dolgushin, Vetrov and Korableva2015). The geological structure of the VG site and the territory adjacent to it shows a complete absence of lithic raw material sources. The nearest accessible bedrock outcrops and alluvial pebble deposits that potentially would have been suitable for lithic toolmaking are located 90 km to the southeast of VG, in the valley of the Ob River. Thus, all lithic artifacts found at VG were brought to the site by humans, but their exact sources are currently unknown (Leshchinskiy et al., Reference Leshchinskiy, Zenin and Bukharova2021).
SITE STRATIGRAPHY
Excavations in 2020 were carried out in an area of 6 × 2 m, on the periphery of the VG central faunal accumulation discovered in 2016. The ABA was located on the mound slope, ~38 m southeast of the 2016–2017 excavation pits (Fig. 1c, d). This area was identified as promising based on the results of exploratory drilling in 2017 (Leshchinskiy and Burkanova, Reference Leshchinskiy and Burkanova2022). In 2021, the excavations continued in an area of 3 × 3 m next to the 2020 pit. The spatial position of the ABA is shown on the stratigraphic section (Fig. 2), and in part of the 2020–2021 excavation plans, with the main paleontological and archaeological finds at a depth of 225–330 cm below the ground surface (Fig. 3). The presence of nearby outbuildings and a vegetable garden is reflected in anthropogenic disturbance of soil of the research area. A summary description of the section is given below.

Figure 2. Cross-section of the Volchia Griva southeastern slope along the line A-B-C/6-7 of 2020–2021 excavation pits (see Fig. 3 for map view). 1 = sand; 2 = silt; 3 = loess-like loam; 4 = soil; 5 = boundaries (a = certain, b = assumed); 6 = desiccation cracks; 7 = holes of burrowing animals; 8 = sedimentary ash cloud; 9 = faunal remains (No. 346 = ABA); 10 = artifacts (green crosses = from 2020 fieldwork, red crosses = from 2021 fieldwork). Not all artifacts and faunal remains found near the line A-B-C/6-7 are shown.

Figure 3. Sketch map of A-B/6-7 – C/7 grids of the Volchia Griva excavation pits in 2020–2021 (squares 1 × 1 m). The artifacts and selected faunal remains are shown from a depth below the zero mark of −225 cm.
Layer 1 is a black massive sandy loam soil of the chernozem type. It has a thickness of up to 0.5 m (the top was cut off).
Layer 2 is a light-brown, massive loess-like sandy loam that is 0.5–0.7 m thick. Rare lithic artifacts are found throughout the layer, mainly in the middle part. In the lower half of the layer, there are a few fragments of poorly preserved large-mammal bones; they all represent the upper level of finds.
Layer 3 is a gray-brown, thinly laminated clayey sand (whitish if dry due to precipitation of Ca and Na salts from pore waters). The sediments dip along a magnetic azimuth of 120–130° (down the mound slope); sediments at the top dip at an angle of ≤5–10°, and from the middle to the bottom at an angle of 10–15° to 35°. In the upper part of the layer, there are rare lithic artifacts and fragments of mammal bones (the middle level of finds). In the lower part of the layer, sediment deformation and erosion benches up to 0.3–0.4 m high are recorded. An ash lens (thickness up to 0.35 m) is clearly visible on the declining bottom unit. The entire base of the layer (especially the ash lens) includes concentrations of lithic artifacts and faunal remains (the lower level of finds). Among them, fragments and whole bones and teeth of foxes predominate; less common are wolves, mammoths, horses, and other mammals. Thickness is 0.8–1.3 m.
The sediments of Layer 3 are disturbed by chaotic cracks that are filled with loess-like sandy loam, beginning on the bedding surface of Layer 2. The cracks are from ≤3 cm to ≥20 cm wide at the mouth, and 0.4–1.3 m long (mean values are 0.7–0.8 m). One crack that is ≥1.7 m long penetrates deep into Layer 4. Near the boundary with Layer 4, including the ash lens, there are numerous holes of burrowing animals 10–14 cm in diameter, mostly filled with motley black-brown sandy loam. Some segments of the holes remain unfilled, and sometimes they cross the cracks.
Layer 4 is a light-brown thinly laminated clayey sand. Lithic artifacts and mammal bones and teeth are distributed throughout the layer; small ash spots ≤20 × 20 cm in size are also found. All of this is a continuation of the lower level of finds. Often there are washout signs (including erosion benches up to 0.5 m), sliding, and deformation of sediments. The bedding surface with Layer 5 dips along a magnetic azimuth of 100–130° at an angle of 10–35° (lines A and B) to 70–80° (line C). Thickness of Layer 4 is 0.1–0.6 m (down the slope, >1 m).
Layer 5 is a light-brown thinly laminated sand with many deformations (breaks in lamination, subsidence of microblocks, and other kinds of distortion). The top is broken by small cracks (length up to 0.3 m, width up to 2–6 cm) that are filled with sand from the overlying Layer 4. The sediments dip along a magnetic azimuth 120–150° at an angle of 5–10° to 20–30°. The visible thickness is >1.3 m.
The described section can be divided into two members: member 1, which is subaerial, consisting of Layers 1–2; and member 2, which is subaerial-subaqueous, consisting of Layers 3–5. The eluvial-aeolian genesis of Layers 1–2 is beyond doubt. The stratigraphic hiatus between the members is obvious and is marked by cracks that probably formed during the drying of sandy-clayey sediments. Similar cracks also were recorded in other VG areas (Leshchinskiy et al., Reference Leshchinskiy, Zenin, Burkanova, Dudko, Gulina, Fedyaev, Semiryakov and Kanishcheva2015, 2021; Leshchinskiy and Burkanova, Reference Leshchinskiy and Burkanova2022). Layers 3–5 reflect the transition from shore to lacustrine shallow water. Signs of shore slope erosion and surf activity on the beach area were accompanied by migration of the shoreline, which is marked by erosion benches. Moreover, the presence of cracks and deformations at the top of Layer 5 may indicate another sedimentation hiatus, preceding the appearance of large mammals, including humans, at the VG site. Member 2 displays a very complex relationship of sediments. It is possible that in this part of the section a larger number of layers with well-founded boundaries will be identified in the future. The holes of burrowing animals described above are of Holocene age, as indicated by their filling with a sedimentary mixture from Member 1, partial hollowness, and intersection of desiccation cracks.
The stratigraphic position of archaeological and paleontological materials found during the 2020–2021 excavations generally corresponds to the previously identified three levels of the VG bone-bearing horizon (Leshchinskiy et al., Reference Leshchinskiy, Kuzmin, Zenin and Jull2008; Leshchinskiy and Burkanova, Reference Leshchinskiy and Burkanova2022, fig. 7). It is important to emphasize that the lithic artifacts and faunal remains at the base of Layer 3 and in Layer 4 represent a single lower level. Figure 2 shows that most of these finds were locally redeposited by temporary streams or were washed away by rain and meltwater down the paleoslope. As a result, many of the finds, including the ABA, are buried in the shallow lake inshore zone. In addition to geological data, this is exemplified by the different degree of preservation of mammalian bones found along the excavation lines of squares A (weathering stages of 3–5 dominate) and B–C (weathering stages of 0–2 prevail).
MATERIALS AND METHODS
The artificial modification of the mammoth femur became apparent immediately upon its discovery (Fig. 4). Preliminary cleaning of the bone revealed a human-made cavity filled with brownish-gray sandy loam that differed sharply from the light brown sand of the enclosing sediments. This fact suggested a high value of the find, so the femur was placed in a sealed plastic bag to prevent contamination. Subsequent processing of the sample was carried out at the Laboratory of Mesozoic and Cenozoic Continental Ecosystems, Tomsk State University (LMCCE TSU). During careful preparation, the remains of a medium-sized carnivore that were found in the bone cavity were documented in detail by sequential digital photography (Fig. 5).

Figure 4. The Volchia Griva section base along the line A-B/6-7 in 2020 (frame: polar fox hemimandible and the ABA in situ, arrow indicates an artificial hole in the mammoth femur metaphysis).
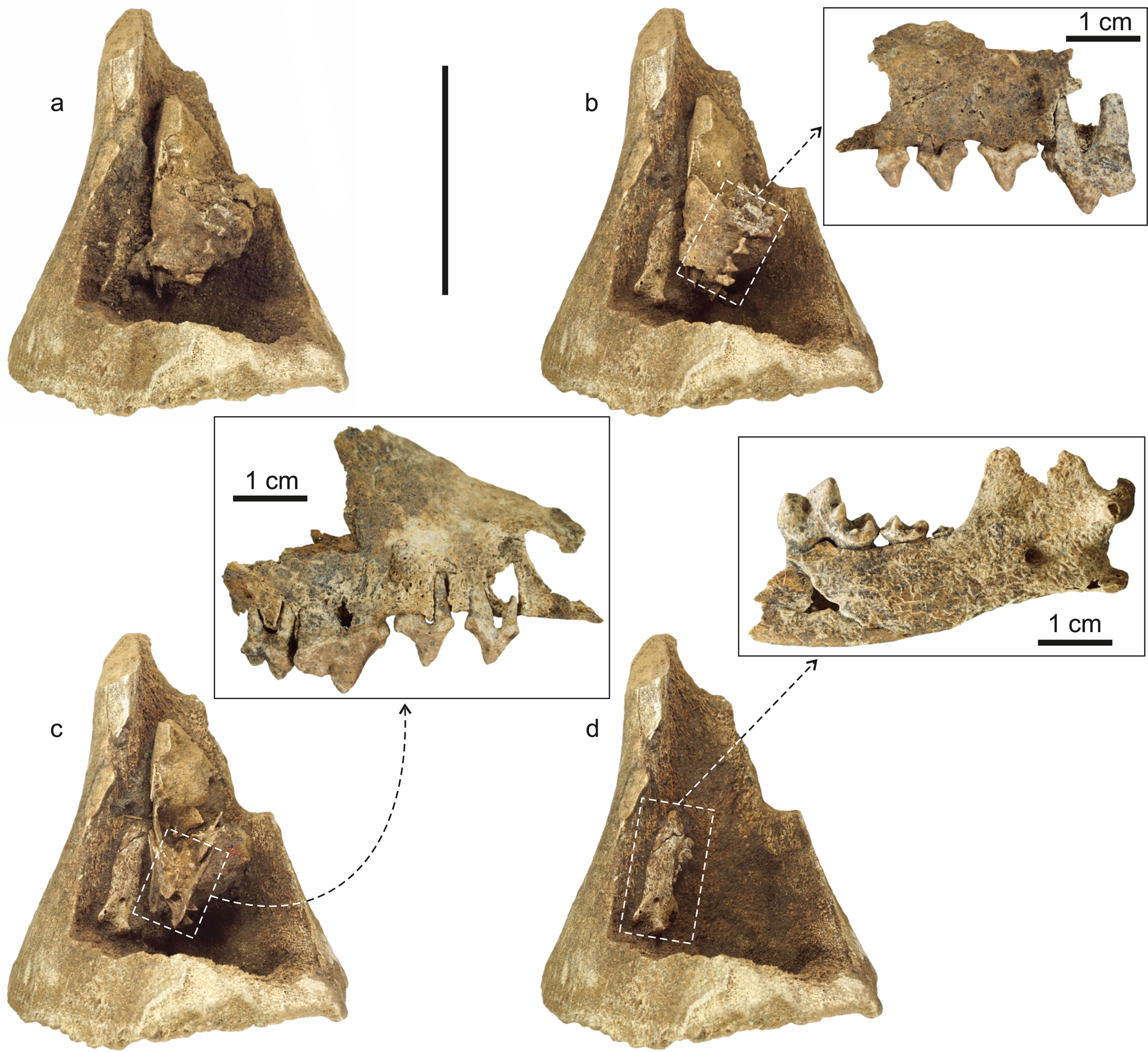
Figure 5. Preparation stages of the ABA (scale = 10 cm). (a) The cavity was opened and the main part of the sandy loam fill was removed; (b) left side of the polar fox partial cranium and the fox hemimandible fragment are partially exposed (frame = left maxilla fragment cleaned, lateral view); (c) polar fox cranial cavities are opened (frame = right maxilla fragment cleaned, lateral view); (d) all sandy loam fill is removed and the hemimandible fragment is completely exposed (frame = right hemimandible fragment is cleaned, lingual view; note full coverage of bone surface by root marks). Some maxillary teeth are not shown because they fell out of the destroyed alveoli.
Thus, the primary materials for this paper are faunal remains representing the described assemblage: (1) a human-modified fragment of a mammoth femur; (2) portions of the cranium and hemimandible, and an isolated tooth of a medium-sized carnivore; and (3) a fragment of rib of a large mammal placed in the cavity. Secondary material was obtained during preparation (cleaning) of the ABA. This is a sandy loam with a high ash content (visible with the naked eye) that filled the rest of the cavity space. Additional material include data for the geological section of the enclosing sediments, and the associated paleontological remains and lithic artifacts.
Taphonomic analysis of the fossil remains began in the field, and was completed at the LMCCE TSU. Examination was performed visually using a 10× magnifying glass, and a stereomicroscope Leica M 205 С (with digital camera FLEXACAM C1) with magnification up to 200×. The preservation of bone surfaces was assessed according to the weathering stages established by Behrensmeyer (Reference Behrensmeyer1978). The taxonomy and individual ages of the animals were determined by comparison with the collections stored at the LMCCE TSU, and with published data on modern and Pleistocene mammals of the families Elephantidae and Canidae (Baryshnikov et al., Reference Baryshnikov, Kuzmina, Hrabrii and Starobogatov1977; Kuzmina and Sablin, Reference Kuzmina, Sablin, Baryshnikov and Kuzmina1993; Sablin, Reference Sablin, Kuzmina and Baryshnikov1994; Lister, Reference Lister1999; Maschenko, Reference Maschenko2002; Baryshnikov, Reference Baryshnikov2006; Gimranov et al., Reference Gimranov, Kosintsev and Gasilin2015; Gasilin et al., Reference Gasilin, Nanova, Kosintsev and Gimranov2017; Gimranov, Reference Gimranov2021).
Palynological analysis of the sandy loam material that filled the cavity in the femur was performed at the LMCCE TSU. Chemical preparation was carried out according to the standard procedure for this type of sediment: the sample was successively treated with Na4P2O7, and then with 10% HCl (Zaklinskaya and Panova, Reference Zaklinskaya and Panova1986). Demineralization was carried out by fractionation in a heavy liquid (density of 2.2 g/cm3) based on a K-Cd mixture.
The prepared cover-glass samples were viewed under a Leica DM 1000 microscope with magnifications of 400, 630, and 1000×. Atlases, published articles, and the LMCCE TSU comparative collections were used to determine pollen and spores. The pollen spectrum (PS) documents the composition and ratio of individual seed plant taxa. The main percentage sum of PS includes the pollen of trees and shrubs (arboreal pollen “AP”), and herbs and dwarf shrubs (non-arboreal pollen “NAP”). Reconstruction of the environment corresponding to the time of the ABA burial was carried out, taking into account the contextual geological data and results of taphonomic studies.
Typological analysis of the associated lithic artifacts was carried out at the Institute of Archaeology and Ethnography (IAET), the Siberian Branch of the Russian Academy of Sciences (Novosibirsk, Russia). A standard methodology was used to describe the Paleolithic materials based on the morphology and metric characteristics of each specimen (Derevianko et al., Reference Derevianko, Markin and Vasil'ev1994, Reference Derevianko, Molodin, Zenin, Leshchinskiy and Mashchenko2003). For this, we used a 10× magnification glass, and optionally a stereomicroscope with magnification up to 300×.
Radiocarbon dating and measurements of stable isotope ratios (δ13C and δ15N) for the bone samples were conducted at the Center for Collective Use “Laboratory of Radiocarbon Dating and Electron Microscopy,” Institute of Geography, Russian Academy of Sciences (Moscow, Russia; laboratory code IGANAMS), and at the Center for Applied Isotope Studies, University of Georgia (Athens, GA, USA). First, the bone was cleaned with a wire brush, and washed using an ultrasonic bath. After physical cleaning, the dried bone was gently crushed into small fragments. The crushed bone was treated with 1N HCl at 4°C for 24 hours. The residue was filtered, treated with 0.1N NaOH on filter to remove humic acid contamination, rinsed with deionized water, and under slightly acid conditions (pH = 3) heated at 80°C for six hours to dissolve collagen and leave humic substances in the precipitate. The collagen solution was then filtered to isolate pure collagen and freeze-dried. The dried collagen was combusted at 575°C in an evacuated/sealed Pyrex ampoule in the presence of CuO. The resulting carbon dioxide was cryogenically purified from the other reaction products and catalytically converted to graphite using the method of Vogel et al. (Reference Vogel, Southon, Nelson and Brown1984). Graphite 14C/13C ratios were measured with the CAIS 0.5 MeV Accelerator Mass Spectrometer (AMS). Sample ratios were compared to the ratio measured from the Oxalic Acid I (NBS SRM 4990). The quoted uncalibrated 14C dates have been given in radiocarbon years before 1950 (years BP), applying the 14C half-life of 5568 years. The error is quoted as one standard deviation and reflects both statistical and experimental errors. The 14C dates have been corrected for isotope fractionation. Calibration of radiocarbon dates was performed using the Calib 8.1.0 program (http://calib.org/calib/), based on the IntCal20 dataset (Reimer et al., Reference Reimer, Austin, Bard, Bayliss, Blackwell, Ramsey and Butzin2020), with ± 2σ rounding calibrated dates to the next 10 years, and combining all calibrated intervals if necessary. The collagen 13C/12C and 15N/14N ratios were measured separately using a stable isotope ratio mass spectrometer and expressed as δ13C and δ15N with respect to standards (PD belemnite and AIR, respectively), with an error of less than ± 0.1‰.
LOCATION AND ACCESSION OF MATERIALS
The described paleontological and archaeological materials are stored at the LMCCE TSU. Fieldwork was carried out based on licenses NOV 02895 PD and NOV 02902 PD, issued by the Department for Subsurface Use, Siberian Federal District of the Russian Federation for TSU.
RESULTS
The ABA structure
The basis of the described assemblage is the distal portion of the left femur belonging to a young mammoth (field No. 346), ~180 mm long, without the epiphysis, which had not been fused. The maximum lateral and anteroposterior diameters are ~150 mm and >110 mm, respectively (the anterior portion of the metaphysis is locally destroyed). On the anterior side of the femur, a cavity close to rectangular in shape, ~165 × 100 mm in size, is hollowed out up to 80 mm deep (Fig. 6). The edges of the cavity were shaped by at least seven large impact-made notches ~1–2.5 cm wide. It should be noted that radial and ring cracks up to 40 mm long diverge from the transverse impact marks, penetrating deeply into the compact tissue (Fig. 6a, b). The main part of the cancellous tissue was removed during cavity construction, and the metaphysis was perforated with two holes ~44 × 20 mm and ~8 × 5 mm maximum sizes (Fig. 7b). On the opposite side of the femur (posterior surface), there are several deep longitudinal cracks (maximum length ~55 mm) running from the broken edge of the diaphysis towards the metaphysis. These cracks indicate at least five strong blows to the midshaft end (Fig. 7c, d).

Figure 6. Completely prepared cavity in the anterior side of a mammoth femur with signs of artificial modeling (scale = 10 cm). Solid white arrows indicate impact marks, solid black arrows indicate root marks, dotted white arrows indicate cut marks. (a) Impact-made notch >2 cm wide with impact mark, one small ring, and two large radial cracks; (b) impact-made notch >2 cm wide with two impact marks and small radial cracks; (c) impact-made notch <1 cm wide with impact mark, around several ring cracks; (d) cut marks; (e) two impact-made notches, each >1 cm wide, with two impact marks and several ring cracks; (f) impact-made notch >1 cm wide with impact mark, small ring, and semi-radial cracks. Note the almost right angles of the cavity base (c, f).
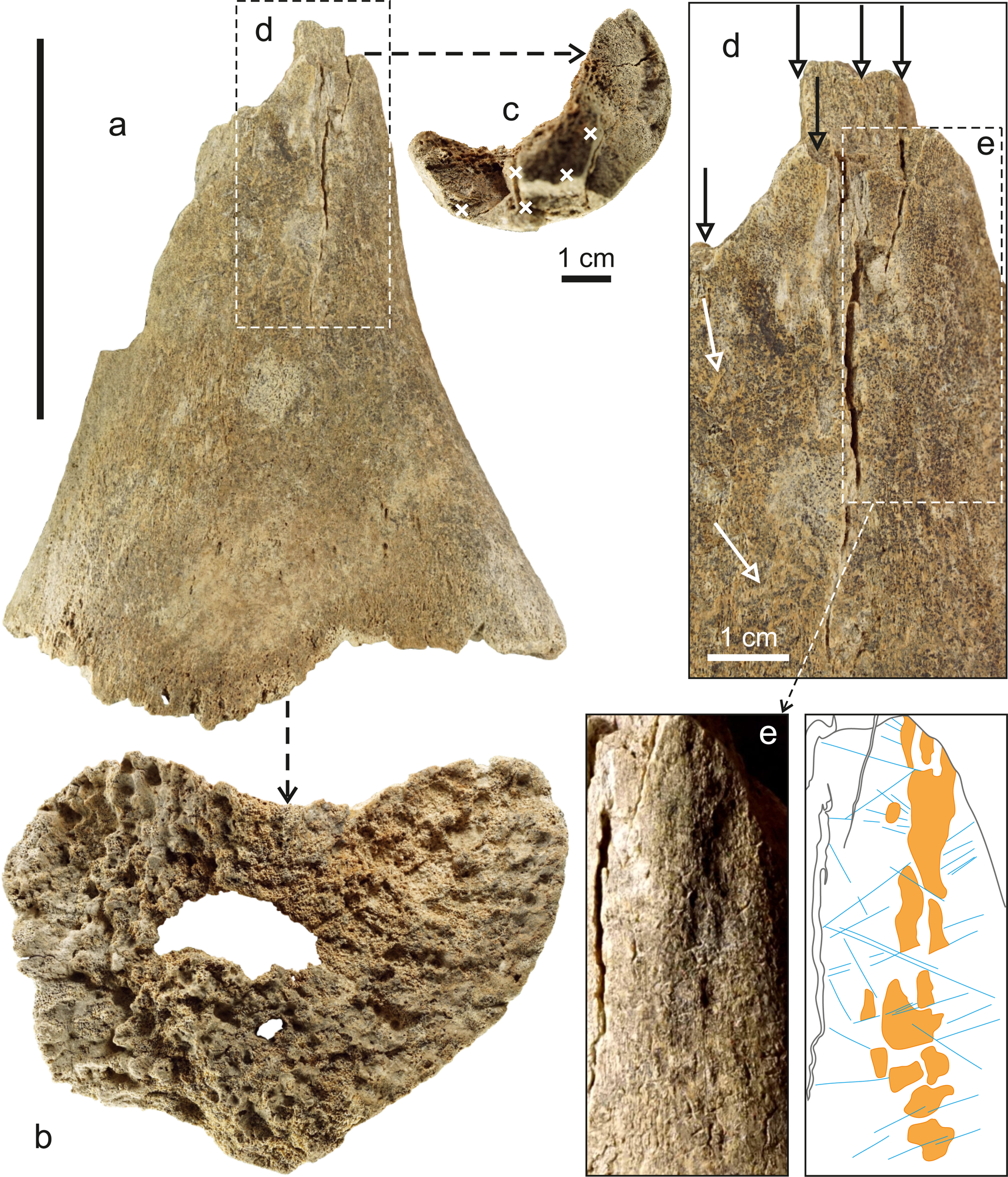
Figure 7. Posterior side of mammoth femur with signs of artificial processing and utilization (scale = 10 cm). (a) General view; (b) perforated metaphysis; (c) transverse midshaft fracture (crosses indicate impact points); (d) area with deep longitudinal cracks (black arrows indicate impact points, white arrows indicate root marks); (e) planoconvex bone portion used as a passive retoucher, drawing shows impressions (orange spots) and straight cuts (blue lines).
Other remains of the described assemblage were found in the cavity (Fig. 5). These are portions of a cranium and mandible, and an isolated tooth, all belonging to foxes, and a fragment of a large rib. A portion of the fox cranium (field No. 347) is represented by nasalia, frontalia, maxillaria, palatinum, and mesethmoideum bones, and the vomer. Most of them are not complete. In the left maxilla, five teeth (all premolars and M1), and in the right maxilla, six teeth (all premolars and molars) remain. There is no visible tooth wear (Fig. 5b, c). The morphology and sizes of the bones and teeth point to species identification as the polar fox (Vulpes lagopus L.).
A fragment of a gracile mandible (field No. 348) is represented by the posterior portion of the right hemimandible with three molars; there is no visible tooth wear (Fig. 5d). Taxonomy due to incomplete bone size and similar morphology of teeth for Eurasian foxes can be tentatively determined as Vulpes cf. V. lagopus L.
An isolated tooth (field No. 350) found in the sandy loam fill is a left M1 of Vulpes sp. The similarity in morphology and size of this tooth for Eurasian foxes does not allow us to give an exact species determination. There is no visible tooth wear.
A fragment of a rib (field No. 349) was also found in the sandy loam fill; it has a length of 14 mm and a cross-section of >17 × 9 mm. These parameters do not allow us to assign it to a species, but it clearly belongs to a large mammal (e.g., mammoth, horse, bison, etc.).
Taphonomic analysis
The femur fragment with the artificial cavity is generally well preserved. Weathering of bone surfaces is at the zero stage. There are no signs that the bone was burned. At the same time, slight chemical etching of the outer compact layer is recorded (edges of fractures and cracks are rounded), including rare marks of plant roots (Figs. 6b, 7d). This made it difficult to diagnose other possible use traces. Nevertheless, they were identified on the lateral inner surface of the femur as a string of impressions of different depths over an area of ~62 × 10 mm (Fig. 7d, e). Some impressions have linear, cut-like edges. There are also several tens of linear discontinuous cut marks (usually <10 mm long, but some are >25 mm), regularly going in two directions relative to each other, at angles from ~40° to ~90°. Cut marks are barely noticeable, and sometimes they lie on top of the impressions (Fig. 7e). A separate clear and straight cut mark ~15 mm long (one end bifurcates) is located on the bone surface near the edge of the cavity (Fig. 6d).
The cranial bones of the fox are small and thin. They are not very well preserved; therefore, determining the weathering stage of such remains is problematic. The bone surfaces, both outside (to a greater extent) and inside the cranium, are covered with marks of plant roots, which are also present on the teeth. On the right maxilla, in a localized area above P3 and P4 (~0.5 cm2), at least 10 short linear incisions with a length of up to 10 mm were identified. These linear incisions can be divided into two distinct groups: 0.01–0.02 mm wide, and 0.07–0.15 mm wide (Fig. 8). Similar marks, up to 10 mm long and <0.09–0.2 mm wide, were also recorded on the right frontale (Fig. 9). Signs of true cut marks on green bone for the ABA in general include a mostly V-shaped transverse profile, linear sharp edges, discontinuous and non-parallel marks of most incisions (Figs. 6d, 7e, 8, 9), and the presence of rare double (Fig. 8) and triple (Fig. 9) marks. There are no signs that the cranium was burned.

Figure 8. Skinning signs on the polar fox maxilla, lateral view (P1 glued into place, M2 not shown). Solid and dotted arrows indicate root and cut marks, respectively.

Figure 9. Skinning signs on the polar fox frontalia, dorsal view. Solid and dotted arrows indicate root and cut marks, respectively.
The hemimandible portion, together with its teeth, is completely covered with root marks. This does not allow us to determine the weathering stage or possible use indicators (cut marks, bone perhaps used as a retoucher, etc.). The same can be said about the isolated M1 tooth, which also has root marks. The fragment of the large mammal rib is poorly preserved. Its weathering cannot be assessed because almost all of the compact tissue has been destroyed. There are no signs that any of these remains were burned.
Associated paleontological material
More than 1000 faunal remains were found in proximity to the ABA (Figs. 2, 3). Most of them are represented by small fragments (up to 10 mm) of bones and teeth recovered during the screen washing of bone-bearing sediments. Only a few dozen large fragments of ribs, vertebrae, limb bones, and tusks belong to woolly mammoth. The fully preserved forelimb of a wolf in anatomical position (excavation squares A6–7) coincides with the ABA level (245–250 cm below the ground surface). Almost all other remains belong to foxes (including the cranium articulated with the mandible in square C7, and isolated portions of the maxillaria and mandibles), of which >90% belong to the polar fox. Cut marks, which indicate skinning, are found on some well-preserved fox bones.
The described deposit differs sharply from most of the VG central faunal accumulation, where there is a record high concentration of large-mammal remains, the vast majority of which belongs to woolly mammoth (Leshchinskiy and Burkanova, Reference Leshchinskiy and Burkanova2022). Thus, the taphonomic picture of human activity in the area of the Paleolithic site clearly defines its functional significance and fundamental difference from the natural paleontological burial of megafauna, although there are only a few tens of meters between them.
Associated archaeological material
More than 750 artifacts were found in the immediate vicinity of the ABA, of which 99 lithics and one bone flake were precisely recorded in space and indicated on the excavation plan. The rest of the specimens were found during screen washing of sediments, and these are mainly small waste products. The main concentration of artifacts is observed within the ash lens (Figs. 2, 3). The petrographic diversity of the lithic industry is limited to two large rock groups: (1) flint-jasperoid and (2) rock crystal-smoky quartz (Table 1). The collection includes products of primary splitting (cores, flakes, chunks, chips, blades, and bladelets and their fragments), and items with secondary treatment defined as tools (Fig. 10).

Figure 10. The Volchia Griva lithic artifacts (2020 and 2021 excavations). (a, c) Cores; (b) chisel; (d–g) burins (arrows indicate directions and quantity of burin blows); (h) flake; (i, l) retouched blades; (j) double-end scraper; (k, m, n) backed bladelets. Petrographic composition: smoky quartz (c), rock crystal (e, m), flint (g–i), jasperoid (a, b, d, f, j–l, n).
Table 1. The petrographic composition and categories of the Volchia Griva lithic artifacts around the ABA.

Cores are maximally residual and represent two reduction types: (1) edge-faceted (Fig. 10.1) and (2) flat (Fig. 10.3). Both types were aimed to obtain blade-like debitage. This is confirmed by the high proportion of blades and bladelets among both the waste products and tools (Fig. 10.4, 10.6, 10.9, 10.11–10.14). Flakes and their fragments are usually <2 cm long, but sometimes reach a length of ≥3 cm (Fig. 10.8).
The ratio of tools in the lithic collection is very high (>10%). They (in total, 12 items) are represented by burins (Fig. 10.4–10.7), blades with continuous retouch on one or two edges (Fig. 10.9, 10.12), a retouched flake, backed bladelets (with a fine marginal lateral retouch; Fig. 10.11, 10.13, 10.14), a chisel (Fig. 10.2), and a double-end scraper (Fig. 10.10).
Radiocarbon dating and stable isotope analysis
Direct 14C dating of the mammoth femur and polar fox cranium (vomer, mesethmoideum, and palatinum bones) was performed. The collagen content of these samples is 17.00% and 15.05%, respectively, which is in the range for high reliability of the analysis. The results are (probability 95.4%, ± 2σ): femur—19,125 ± 40 yr BP (IGANAMS-8477); cranium—19,245 ± 45 yr BP (IGANAMS-8478). Calibrated ages for these 14C values are 22,930–23,120 cal yr BP and 22,980–23,660 cal yr BP, respectively. The atomic C/N ratio of the collagen for both samples is 3.14, within the range of 2.9–3.6, which is recommended for reliable data (Ambrose, Reference Ambrose1990; Guiry and Szpak, Reference Guiry and Szpak2021). Stable isotope data for bone collagen samples from the ABA are as follows: woolly mammoth, δ13C = −20.02‰, δ15N = 12.50‰; polar fox, δ13C = −13.13‰; δ15N = 16.81‰.
Palynological analysis
A sample from the sandy loam fill of the ABA yielded a sufficient amount of pollen for reliable palynological analysis, the results of which are shown on Figure 11. PS consists of 315 grains belonging to AP and NAP. The amount of AP is only 7%; among which Pinus s/g Dyploxylon dominates. Pollen of other trees (Piceae sp., Betula sp.) and shrubs (Alnus sp., Salix sp.) is very rare. Thus, NAP strongly dominates in the PS; Tricolpopollenites (tricolpate pollen with an unclear exine ornamentation) has the largest share, 31.4%. A large amount of pollen belongs to the families Brassicaceae (18.4%), Asteraceae (14.2%, including 7.9% of Artemisia sp.), and Rosaceae (10.2%). The pollen of Chenopodiaceae constitutes 2.9% of total NAP. Pollen of Fabaceae, Cichoriaceae, Polygonaceae, and Scrophulariaceae is grouped into “other herbs”; the share for each of these taxa is <2%. One pollen grain of Potamogeton sp., a typical representative of aquatic plants, is identified. No spores were found. Non-pollen palynomorphs are represented by rare Pediastrum sp., fungal fruiting bodies of Glomus sp., and hyphae and fungal spores of indeterminate taxa. Fragments of sponge spicules and phytoliths of dicotyledonous plants are also found.

Figure 11. Pollen diagram for the ABA fill (AP = arboreal pollen; NAP = non-arboreal pollen; AP and NAP of the main percentage sum).
A large amount of finely dispersed charcoal/ash, visually recorded in the ABA fill, was confirmed in the palynological cover-glass preparations. Especially indicative is the contrast with the sediments of the lower bone-bearing level in the 2016 excavation pit (Leshchinskiy and Burkanova, Reference Leshchinskiy and Burkanova2022), in the samples of which there are almost no charcoal particles (Fig. 12).

Figure 12. Palynological cover-glass preparations of the ABA fill (a, b) and sediments of the lower bone-bearing level in the 2016 excavation pit (c, d). Solid, dashed, and dotted arrows indicate charcoal/ash particles, fruit bodies of Glomus sp. fungi, and plant tissue remains, respectively.
DISCUSSION
The presence of humans in the West Siberian Plain during the LGM has been considered debatable (e.g., Goebel, Reference Goebel, Elston and Kuhn2002; Graf, Reference Graf, Camps and Chauhan2009; Chlachula, Reference Chlachula2017; Kuzmin and Keates, Reference Kuzmin and Keates2018). By modeling the process of human colonization of Eurasia in the Late Pleistocene, it was clearly demonstrated that at ca. 20 ka BP, only in the extreme south of the region, a small population with a density of ≤2 persons per 100 km2 can be assumed. As for the rest of the West Siberia Plain, a density of ≤1 person per 100 km2 or completely uninhabited has been suggested (Timmermann and Friedrich, Reference Timmermann and Friedrich2016). This conclusion is generally supported by the limited reliable traces of human presence in most of the West Siberian Plain at that time. The short-term sites of Tomsk and Shikaevka, with a few 14C dates of ca. 18.3–18 ka BP, were recorded in the southeastern and southwestern parts of the region, respectively (Zenin, Reference Zenin2002). These sites are clearly associated with mammoth remains, which implies a direct dependence of ancient humans on these animals. The same connection is obvious for the largest sites of this period (Shestakovo and Volchia Griva) associated with a natural mass accumulation of mammoth remains in the southeastern and southern peripheries of the region. The large Achinsk site and a promising mammoth site with Paleolithic artifacts called Krasnoyarskaya Kurya are in relative proximity to these sites. Verification of the geological age for all of these sites is represented by >40 14C dates tightly bunched within the interval of 25.7–17 ka BP (Derevianko et al., Reference Derevianko, Zenin, Leshchinskiy and Mashchenko2000, Reference Derevianko, Molodin, Zenin, Leshchinskiy and Mashchenko2003; Seuru et al., Reference Seuru, Leshchinskiy, Auguste and Fedyaev2017; Leshchinskiy et al., Reference Leshchinskiy, Zenin and Bukharova2021; Leshchinskiy and Burkanova, Reference Leshchinskiy and Burkanova2022). A similar range can be seen using 14С dates of the mammoth fauna and the Late Paleolithic sites in the Gari area, situated in the western part of the West Siberian Plain (Chlachula and Serikov, Reference Chlachula and Serikov2010). Thus, there is no doubt that human populations existed in the southeastern, southern, southwestern, and western parts of the region during the first half of MIS 2. The absence of LGM sites in the central, northern, and eastern parts of the West Siberian Plain is commonly assumed to be the result of various geographical barriers (e.g., severe climate, ice sheets, ice-dammed lakes, etc.). However, this absence also can be explained by the relatively poor degree of survey of this vast territory.
Taphonomy and paleoenvironmental conditions
The ABA from the VG site has no current analogues either in the Paleolithic of Siberia or, possibly, the entire Northern Hemisphere. The base of the ABA from the VG site is made from a mammoth femur shaft; a medium-sized bone with an unfused epiphysis that indicates a young adult animal, with an age estimate of 12–20 years (Baryshnikov et al., Reference Baryshnikov, Kuzmina, Hrabrii and Starobogatov1977; Lister, Reference Lister1999; Maschenko, Reference Maschenko2002). The zero stage of weathering of the bone surfaces indicates a relatively rapid burial. In the arid African savanna, with extreme temperature fluctuations due to high ultraviolet radiation, this corresponds to a period of up to one year (Behrensmeyer, Reference Behrensmeyer1978; Haynes and Wojtal, Reference Haynes and Wojtal2022). In the tropical rainforest, with very little direct light and high humidity, weathering stage zero can sometimes last >15 years; and in temperate and subarctic climatic conditions, weathering stage zero probably can last >7 years, but mostly no more than three years (Tappen, Reference Tappen1994; Haynes, Reference Haynes1999).
What were the environmental conditions in the southern part of the West Siberian Plain around the LGM? This topic has been discussed for over 50 years. Many paleogeographic reconstructions suggest radically opposite environments: from swampy tundra or forest tundra flooded by the waters of the giant ice-dammed Lake Mansi to loess- and sand-covered cold desert with regosolic gleysols, extremely low winter air temperatures, and deep ground freeze (Arkhipov, Reference Arkhipov1971; Arkhipov et al., Reference Arkhipov, Astakhov, Volkov, Volkova and Panychev1980; Arkhipov and Volkova, Reference Arkhipov and Volkova1994; Chlachula, Reference Chlachula2003; Hubberten et al., Reference Hubberten, Andreev, Astakhov, Demidov, Dowdeswell, Henriksen and Hjort2004; Astakhov, Reference Astakhov2006; Zykin and Zykina, Reference Zykin and Zykina2009; Chlachula and Little, Reference Chlachula and Little2011; Velichko et al., Reference Velichko, Timireva, Kremenetski, MacDonald and Smith2011). Based on the most recent data, for the last 10,000 years of the Pleistocene, this region, including the Baraba Lowland, was characterized by widespread development of forb-grass steppes (Leshchinskiy and Burkanova, Reference Leshchinskiy and Burkanova2022), which indicates a dry climate and high summer insolation. Consequently, the weathering stages of large-mammal bones, described in modern thanatocenoses, indicate that burial of the mammoth femur's fragment at VG should have occurred within 1–3 years after death of the animal. This is consistent with the contemporary environmental conditions of northwestern Mongolia, which are the closest to those at the LGM in the southern part of the West Siberian Plain (Leshchinskiy, Reference Leshchinskiy2015). However, the described specimen could have remained on the ground surface longer under favorable conditions (e.g., the shore zone of a lake with high humidity, and shade, etc., including the factor of human influence). Root marks on the femur's surface could have formed both in subaerial and subaqueous environments, since the ABA was finally covered with sediments in lacustrine shallow water.
The special conditions at the periphery of the Paleolithic site may reflect the PS of sandy loam fill of the ABA. The ratio of AP and NAP shows that it generally corresponds to the PSs from the coeval sediments of the lower bone-bearing level, which was excavated in 2016 (Fig. 1d). Those PSs reflect the widespread steppes, as well as the presence of a lake with developed coastal and aquatic vegetation at the foot of the VG mound. In general, this situation persisted until the beginning of loess-like sandy loam accumulation (Leshchinskiy and Burkanova, Reference Leshchinskiy and Burkanova2022). However, the PS of the ABA shows a more than threefold increase in the pollen content of the cabbage family (Brassicaceae)—18.4% of AP+NAP. At the same time, its share among all detected herbs is unusually high, and amounts to almost 20%.
For comparison, published PSs from Late Pleistocene deposits in the central and southern parts of the West Siberian Plain, including mammoth fauna and Paleolithic sites, have a share of Brassicaceae of ≤10% of the herb composition, and even less of AP+NAP (Volkova et al., Reference Volkova, Bukreeva, Poleshchuk, Grichuk and Saks1970; Arkhipov et al., Reference Arkhipov, Votah, Levina, Saks, Arkhipov, Volkov, Volkova and Troitskiy1973, Reference Arkhipov, Dovgal’, Levina, Lipagina, Suhorukova and Hlonova1976; Levina, Reference Levina and Volkova1979; Volkova and Volkov, Reference Volkova, Volkov, Saks, Volkova and Hlonova1980; Krivonogov et al., Reference Krivonogov, Bakhareva, Kim, Orlova and Skabichevskaya1993; Volkova and Mikhailova, Reference Volkova and Mikhailova2001; Mikhailova, Reference Mikhailova2004; Burkanova, Reference Burkanova, Rozanov, Lopatin and Parkhaev2006). The proportion of Brassicaceae pollen as ≤10% of AP+NAP also has been reported for Late Paleolithic sites in Siberia and Eastern Europe (Gladilin and Pashkevich, Reference Gladilin, Pashkevich, Ivanova and Praslov1977; Gubonina, Reference Gubonina, Ivanova and Praslov1977; Pashkevich, Reference Pashkevich, Goretskiy and Tseytlin1977; Praslov et al., Reference Praslov, Levkovskaya, Kulkova, Ivanova and Praslov1977; Mochanov et al., Reference Mochanov, Fedoseeva, Alekseev, Kozlov, Kochmar’ and Shcherbakova1983; Tseytlin, Reference Tseytlin and Molodin1985; Lbova et al., Reference Lbova, Kolomietc, Dergacheva, Fedenyova and Klementyev2005; Spiridonova, Reference Spiridonova and Voeykov2006; Velichko et al., Reference Velichko, Pisareva, Sedov, Sinitsyn and Timireva2009; Tashak et al., Reference Tashak, Reshetova and Antonova2017, Reference Tashak, Antonova and Reshetova2019). Based on contemporary data, 58 species of this economically important family grow in the territory of Novosibirsk Province, 12 of which are found in steppe and alkali meadows, 10 species along the river and lake shores, and one species is an aquatic plant (Krasnoborov et al., Reference Krasnoborov, Lomonosova, Shaulo, Vibe, Zhirova, Korolyuk, Krasnikov, Snytko and Tupitsyna2000). Our studies of various surface PSs at VG showed that Brassicaceae pollen is rare in recent samples (Leshchinskiy and Burkanova, Reference Leshchinskiy and Burkanova2022).
Thus, the available data indicate that the natural background of Brassicaceae pollen in the Late Pleistocene of Siberia and Eastern Europe probably amounted to ≤10% of AP+NAP. The anomalously high content of this pollen in the ABA fill, in our opinion, could be due to two factors: (1) the specific plant composition of the area (dry sandy loam slope and proximity to the lake) and (2) human activity (collection of certain plants or their parts off site for bedding, nutrition, use as a medicine, or other purposes, including aesthetic and ritual ones). Moreover, both factors could have acted either separately or simultaneously.
The mammoth femur and the polar fox cranium from the ABA were studied for δ13C and δ15N stable isotopic composition of bone collagen. The results are almost identical to the data on the mammoth and canine bones from the 2016 excavation pit, which revealed an abnormal biogeochemical composition of the VG mammalian diet (Leshchinskiy and Burkanova, Reference Leshchinskiy and Burkanova2022). Thus, the elevated δ15N value (12.50‰) for the mammoth femur is close to the range of 12.86–14.67‰ previously determined for mammoths from the VG lower bone-bearing level. In other parts of Eurasia and Eastern Beringia, the average δ15N values for mammoths dated to 20–10 14C ka BP are <10‰ (Bocherens et al., Reference Bocherens, Pacaud, Lazarev and Mariotti1996; Szpak et al., Reference Szpak, Gröcke, Debruyne, MacPhee, Guthrie, Froese, Zazula, Patterson and Poinar2010; Seuru et al., Reference Seuru, Leshchinskiy, Auguste and Fedyaev2017; Drucker et al., Reference Drucker, Stevens, Germonpré, Sablin, Péan and Bocherens2018; Kuzmin et al., Reference Kuzmin, Bondarev, Kosintsev and Zazovskaya2021). The δ13C and δ15N values for the polar fox cranium are also unusual: −13.13‰ and 16.81‰, respectively. They go far beyond the general range for Eurasian and Eastern Beringian populations: δ13C from approximately −23‰ to −17‰; and δ15N from ~4–10‰ (Angerbjörn et al., Reference Angerbjörn, Hersteinsson, Lidén and Nelson1994; Dalerum et al., Reference Dalerum, Perbro, Magnusdottir, Hersteinsson and Angerbjörn2012). The new data obtained on δ13C and δ15N for mammalian bone collagen at VG confirm the trophic enrichment of the carnivores (Bocherens, Reference Bocherens2015), and indicate that carrion of large herbivores could have been the basis of the polar fox diet. In turn, an elevated δ15N value for mammoth bone collagen is expected for the ecosystem of fertile forb-grass steppe under conditions of a favorable geochemical landscape formed on the VG mineral oasis (Leshchinskiy and Burkanova, Reference Leshchinskiy and Burkanova2022).
Some or even all of the components of the ABA could have been obtained from the VG thanatocenosis that already existed by that time. This is perhaps reflected in the varying coverage of the bone surfaces with root marks, which is why it has not yet been established if the cranium and hemimandible belong to one or two individuals. However, the presence of an isolated left M1 tooth and another M1 in the preserved portion of the cranium indicate that the remains of at least two foxes were buried in the bone cavity. Dates of 19,245 ± 45 14C yr BP and 19,125 ± 40 14C yr BP for the polar fox cranium and mammoth femur, respectively, are in good agreement with previous results for the main period of formation of the VG lower bone-bearing level at ca. 19.6–19.0 14C ka BP (Leshchinskiy and Burkanova, Reference Leshchinskiy and Burkanova2022).
Late Paleolithic activity
Considering the ABA in context, it can be argued that it is clearly human made. The regular impact notches on the edges of the femur's cavity indicate the purposeful use of a heavy hammerstone, with a working part width probably ≥25 mm. In this case, the work was carried out on green bone, as can be seen from the pattern of the accompanying cracks. Experimental study on the splitting of elephant long bones fully supports this conclusion (Haynes et al., Reference Haynes, Krasinski and Wojtal2021). At the same time, the almost right angles of the external base of the cavity (Fig. 6c, f) may indicate use of a thin tusk, bone, or antler as an intermediate tool with a width of the working part of ≤10 mm. The mechanism of formation and significance of the large hole in the metaphysis are unclear, but it is obvious that the large hole was made intentionally, and the small hole may have appeared accidentally during formation of the cavity. All these damages do not compare with the gnawing marks of bones by large carnivores: “…Gnawing animals may lick and grind fracture edges smooth, or abrade fracture edges against paws or ground surfaces while gnawing opposite ends. After long bones have been broken apart by carnivores, parts of bones unmarked by teeth will have step-fractures on edges that might also be rounded…” (Haynes and Stanford, Reference Haynes and Stanford1984, p. 225).
The mammoth femur was used previously as a heavy passive retoucher for relatively large lithic tools. This is evinced by typical widespread chipping from pressure retouching in the form of impressions, which mark a planoconvex zone of active use with a great length (>6 cm). The mammoth femur is not completely preserved, and part of it apparently was broken off during or before formation of the cavity (Fig. 7e). At the same time, discontinuous linear cut marks, sometimes overlying impressions, may reflect the beveling process. A combination of similar marks is often found on Paleolithic bone retouchers in Northern Eurasia (Moigne et al., Reference Moigne, Valensi, Auguste, García-Solano, Tuffreau, Lamotte, Barroso and Moncel2016; Costamagno et al., Reference Costamagno, Bourguignon, Soulier, Meignen, Beauval, Rendu, Mussini, Mann, Maureille, Hutson, García-Moreno, Noack, Turner, Villaluenga and Gaudzinski-Windheuser2018; Mozota, Reference Mozota, Hutson, García-Moreno, Noack, Turner, Villaluenga and Gaudzinski-Windheuser2018; Martellotta et al., Reference Martellotta, Delpiano, Govoni, Nannini, Duches and Peresani2020; Bello et al., Reference Bello, Crété, Galway-Witham and Parfitt2021). In this aspect, the functional significance of blows to the midshaft butt, which led to the appearance of longitudinal cracks, becomes clear. The blows probably were produced by a heavy (≥2 kg) hammerstone for the purpose of chipping long and thick cortical blanks for retouchers with a regular convex surface. According to some researchers (Mozota, Reference Mozota2013; Kolobova et al., Reference Kolobova, Kharevich, Chistyakov, Kolyasnikova, Kharevich, Baumann, Markin, Olsen and Krivoshapkin2022), the diaphyses of large mammal long bones are the best material for this purpose. Impressions and linear cut marks caused by using the mammoth femur as a retoucher are fundamentally different from the gnawing marks made by predators and scavengers, as well as from marks made by trampling. In this case, the scratches occur singly or as sets of parallel marks, whose outer edges are generally rounded (Haynes, Reference Haynes1982, Behrensmeyer et al., Reference Behrensmeyer, Gordon and Yanagi1986; Haynes et al., Reference Haynes, Krasinski and Wojtal2020). The impressions noted on the mammoth femur are not anatomical features. Diagnosis of mammoth femurs from natural Siberian sites, stored in the LMCCE TSU collections, did not reveal similar patterns of bone topography.
The short linear incisions in the frontal region of the polar fox cranium from the ABA were certainly made with a lithic tool during the skinning process. The tendons were cut in the skull region, near the eye sockets and on the frontal bones. Thus, the typical parameters of incisions and their clear localization on certain skeletal elements are characteristic features of Paleolithic complexes (Charles Reference Charles1997; Landeck and Garriga, Reference Landeck and Garriga2017; Konidaris et al., Reference Konidaris, Athanassiou, Tourloukis, Thompson, Giusti, Panagopoulou and Harvati2018; Wojtal et al., Reference Wojtal, Wilczyński, Wertz and Svoboda2018, 2020). In addition, the fragility of the bones does not support the option of trampling, in which case the skull would have been crushed.
Careful preparation of the polar fox remains as part of the ABA showed that the premaxillae, with incisors and the anterior portions of the maxillae together with the canines, previously were separated from the cranium, which is why they are not in the structure of the assemblage. This can be seen from the position of the preserved cranial portion at the base of the cavity, opposite the large hole (Fig. 5). The absence of parietale, jugale, squamosum, auditory bulla, occipital, and other bones of the braincase and palate may also indicate their deliberate removal by humans. However, some of them (especially the occipital bone) could have been partially destroyed during the burial process due to microbial and chemical degradation (Leshchinskiy, Reference Leshchinskiy2012; Calábková et al., Reference Calábková, Chlachula, Ivanov, Hložková, Czerniawska, Vašinová-Galiová, Prokeš and Gadas2022). Only the posterior portion of the hemimandible, broken between the premolars and molars, was also inserted into the cavity. The position of an isolated tooth and a large rib fragment were not clearly recorded. Nevertheless, we are inclined to think that in the final stage of the creation of the assemblage all remains were sealed by humans in the cavity in a compound made of a mixture of sand, clay, and ash. The artistic reconstruction of the ABA (without the infill, isolated tooth, and rib fragment) is shown in Figure 13.

Figure 13. Artistic reconstruction of the ABA (illustration by Anastasiia S. Leshchinskaya, idea by Sergey V. Leshchinskiy).
One question about the ABA burial remains open: was it carried into the lake by natural agents such as temporary streams, colluvial processes, or wave activity, or was it deliberately placed by humans in shallow water? Movement down the slope and farther into the lacustrine shallow zone for most of the paleontological and archaeological materials recovered during the 2020–2021 excavations is beyond doubt. Although this does not necessarily mean a significant redeposition of paleontological remains and artifacts, it is currently not possible to identify regularities in the distribution of the cultural layer in the studied area from a geological point of view. Therefore, we consider these finds as a single complex of the lower bone-bearing level and associate the chaotic arrangement of artifacts with the destruction of the cultural layer on the VG slope.
The VG lithic collection (Fig. 10) represents a combination of exhausted cores and blades with curved profiles, including items converted into tools. The scar pattern of the blades and flakes determines variants of cores with longitudinal and counter knapping. Backed bladelets and bladelets with a truncated transverse edge, as well as micropoints with a straight profile, are the most numerous among the tools. The lateral and transverse burins are illustrative. Scrapers, chisel-like tools, and borers are rare. Notched tools are typical, including items with a notch on the striking platform. Tools with fine dorsal retouch predominate (counter and opposite kinds of retouch are very rare). Thus, the typological composition and a large percentage of tools (Table 1), within a background of the thick cultural layer saturated with ash and burnt bone fragments, suggest a long-term occupation of the site with the active use of fire. Intensive activity at the working area was aimed mainly at toolmaking, along with butchering of animals and processing of their skins, bones, and teeth. In this case, inset tools were used widely. These data significantly supplement the previous geoarchaeological studies (Leshchinskiy et al., Reference Leshchinskiy, Zenin and Bukharova2021).
The described lithic industry belongs to the bladelet-based complexes of Siberia and the Middle Urals that existed during the LGM. They are characterized by the production of bladelets (2–5 cm long), modified by marginal one-sided retouch. Micropoints, bladelets with a backed longitudinal edge, carinated scrapers, borers, burins, and chisel-like tools are common. Cores, as a rule, are represented by edge-faceted and flat varieties. Wedge-shaped cores are absent, as well as large scrapers. Overall, the VG lithic inventory has the closest analogues in the materials from the West Siberian sites of Shestakovo (geological layers V–VII) and Achinsk, probably reflecting a common cultural tradition (Anikovich, Reference Anikovich and Larichev1976; Derevianko et al., Reference Derevianko, Zenin, Leshchinskiy and Mashchenko2000, Reference Derevianko, Molodin, Zenin, Leshchinskiy and Mashchenko2003). In Eastern Siberia, similar technologies have been observed at the Volchikha (Akimova et al., Reference Akimova, Stasiuk, Tomilova, Tihomirov, Yu, Derevianko and Molodin1997) and Mal'ta sites (layers 8, 9) (Medvedev et al., Reference Medvedev, Slagoda, Lipnina, Berdnikova, Generalov, Rogovskoy, Oshchepkova, Vorobieva and Shmygun2001); and in the Altai region, at the Ushlep 6 site (layer 3) (Kungurov, Reference Kungurov and Kiryushin1996). West of the VG site, they are known at the Talitski site in the Middle Urals (Shcherbakova, Reference Shcherbakova1994).
The commonalities of LGM industries in Siberia are clearly seen in the knapping technique and the main tool categories. Differences are expressed in the details of the secondary treatment of tools and typological diversity. At the same time, there are artifacts that have a striking similarity but belong to sites situated very far apart, including micropoints, short carinated scrapers, bladelets with a notch on the striking platform, backed bladelets, and blades with retouched notches. In addition to VG, these artifacts are found at the Mal'ta, Shestakovo, and Talitski sites. Thus, in a space of almost 3000 km (from Lake Baikal to the Urals and farther to the west) in the interval of 24–17 14С ka BP, Paleolithic technocomplexes with a representative and similar microinventory can be traced.
Despite this similarity, in the Late Paleolithic sites of Siberia, no bone assemblages similar to the ABA from VG have been found so far. The presence of the polar fox cranium without nasal, orbital, and ear regions in a human-made bone cavity may indicate a funeral rite (a mammoth femur could serve as a coffin), or a ritual associated with depriving the head of the sense organs: smell, sight, and hearing. Similar examples are known among the native people of Western Siberia. For example, at the bear festival, the Khanty and Mansi nations perform a ritual ceremony: a hat is put on the head of a killed male bear (a scarf is tied to a female bear), and birch bark circles are placed on the eyes and nose; quite often, coins or metal plates are used instead of the circles. These actions are designed to close all the skull openings in order to interrupt any connection with the real world. At the end of the ceremony, the rhinarium together with the upper and lower lips are cut off from the bear muzzle (Markov and Lukina, Reference Markov and Lukina1987; Gemuev and Baulo, Reference Gemuev and Baulo1999; Kulemzin and Lukina, Reference Kulemzin and Lukina2006; Sokolova, Reference Sokolova2009).
It is important to note that modern aboriginal hunters consider as a sacred animal not only the bear, but also the fox, and some other carnivores. Their skulls, noses, teeth, paws, claws, and other remains may be kept as parts of a totem animal. An example is the cult assemblages of the guardian spirit among the indigenous peoples of Siberia, which include the nose portions of the “sacred” animal heads in association with minerals and metal artifacts (Fig. 14). Similar symbolism is well known in ancient societies of various regions of Eurasia since the Neolithic (Kulemzin et al., Reference Kulemzin, Matyushchenko and Bobrova1994; Baulo, Reference Baulo2002; Kulemzin, Reference Kulemzin2004; Sokolova, Reference Sokolova2009). Information about the possible ritual behavior of Paleolithic people associated with carnivores is rare and contradictory. Such examples include the practice of deliberate burial of bear remains (primarily skulls and long bones) in niche-caches of caves of the Middle–Late Paleolithic in Central Europe (Bednarik, Reference Bednarik2020), and debatable finds of “chopped off” bear craniums with cut marks in the Acheulian cache of the Azykh Cave in the Caucasus (Guseynov, Reference Guseynov1973).

Figure 14. Portions of the assemblage of the family guardian spirit (Mansi People, Yasunt Village, Berezovo District, Khanty-Mansi Autonomous Okrug; field materials of Prof. A.V. Baulo, 2009). The assemblage was made in the middle of the eighteenth century AD and represents two “robes” put on each other (the external one is yellow-brown); between “robes” the following items were found: (a) the mummified noses of an otter (top) and a wolverine (bottom); (b) rock crystal, triangular brass plate with holes, and a 20 kopeck coin belonging to the reign of Empress Elizabeth, wrapped in textile.
Were any rituals performed in Western Siberia at 20–19 14C ka BP, and how complex were the relationships between humans and carnivores at that time? We have yet to receive detailed answers to these questions, but one of the key areas for investigating this problem now is the Baraba Lowland and adjacent regions. New excavations of the VG site demonstrate the great importance of the polar fox in the Late Paleolithic economy of this territory. Comparison with the main LGM sites in Eastern Siberia, Eastern Europe, and Central Europe clearly indicates that the mass procurement of these fur-bearing animals was a paleoecological trend for the human population of Northern Eurasia. Moreover, the polar fox, like other foxes, was hunted not only for its fur, but also for food, and its bones and teeth were used to make household items, tools, and personal ornaments (Sukachev et al., Reference Sukachev, Gromov and Bader1966; Gvozdover and Grigoryev, Reference Gvozdover, Grigoryev, Ivanova and Praslov1977; Velichko et al., Reference Velichko, Gribchenko, Markova, Udartsev, Ivanova and Praslov1977; Verestchagin and Kuzmina, Reference Verestchagin, Kuzmina and Skarlato1977; Ermolova, Reference Ermolova1978; Kuzmina and Sablin, Reference Kuzmina, Sablin, Baryshnikov and Kuzmina1993; Sablin, Reference Sablin, Kuzmina and Baryshnikov1994; Charles Reference Charles1997; Germonpre et al., Reference Germonpre, Sablin, Khlopachev and Grigorieva2008; Camarós et al., Reference Camarós, Münzel, Cueto, Rivals and Conard2016; Baumann et al., Reference Baumann, Wong, Starkovich, Münzel and Conard2020; Pryor et al., Reference Pryor, Pospuła, Nesnídalová, Kowalik, Wojtal and Wilczyński2020; Wojtal et al., Reference Wojtal, Svoboda, Roblíčková and Wilczyński2020; Demay et al., Reference Demay, Julien, Anghelinu, Shydlovskyi, Koulakovska, Péan and Stupak2021). However, despite the abundant fox remains at these sites, there is no evidence of their ritual use. The only case that can be considered as a potential ritual use is recorded at the Předmostí site. During excavations at the end of the nineteenth century, many polar fox bones were found here inside and around human burials, including the authentic “… association of a fox skull that lay across a human skeleton… but given the frequency of fox remains over the whole area it loses any kind of exceptionality…” (Svoboda, Reference Svoboda2008, p. 31).
CONCLUSIONS
The material aspects of human paleoecology in the form of lithic tools, bones, and teeth provide evidence that is useful for solving some problems of physiological adaptation to the LGM environment. At the same time, the spiritual sphere of prehistoric culture is the least tangible and most difficult part of human history to reconstruct. Nevertheless, attempts are made to isolate such sophisticated elements as magic, shamanism, totemism, fetishism, or art in its structure (Braun, Reference Braun, Putová and Soukup2015; Chlachula and Serikov, Reference Chlachula and Serikov2022).
The subtlety and inconsistency of reasoning regarding ritual behavior or aesthetic perceptions in the world of the Late Pleistocene people is obvious. An example is Franco-Cantabrian cave art, much of which can be interpreted as being made by children or adolescents (Bednarik, Reference Bednarik2008). Thus, the ABA from the VG site could well be either a totem doll (imitation) of a guardian spirit or an ordinary or unusual toy for children. In any case, the main features of the ABA include (1) a series of impact notches forming a rectangular cavity in the mammoth femur; (2) removed cancellous tissue from the human-made cavity; (3) the perforated femur metaphysis; (4) the polar fox cranium with cut marks and the anterior portion removed; (5) the fox hemimandible with the anterior portion removed; and (6) longitudinally oriented position of fox remains in a bone cavity. The combination of these features denies the accidental burial of fox remains in the mammoth femur shaft. This creates confidence that the described assemblage is not a product of nature. The choice of mammoth and polar fox bones for its creation was deliberate, since the role of both species in the life of Late Paleolithic humans was extremely great.
Acknowledgments
We are grateful to Alexandra S. Samandrosova, Artur T. Dzhumanov, Svyatoslav S. Perfilev, Alexey S. Klimov, Dmitriy Y. Kadochnikov, Nikolay A. Vlasov, Ekaterina A. Osipova (all from TSU) and Ivan G. Krivokorin (Tallinn University of Technology, Estonia) for assistance in fieldwork. We are indebted to Arkadiy V. Baulo (IAET, SB RAS) for consultations on the ethnography of Western Siberia, and to Svetlana V. Gayduk as the head of the local administration for the opportunity to conduct excavations in 2020–2021 within Mamontovoe Village. We also are grateful to Susan G. Keates (Düsseldorf, Germany) for her help in correction of the English text. Special thanks to Anastasia S. Leshchinskaya (Tomsk, Russia) for the artistic drawing of the reconstructed ABA.
Financial Support
The study was funded by the Russian Science Foundation (RNF), Project No. 20-17-00033.















