



INTRODUCTION
Archaeological excavations of early commercial fishing sites can provide information on past economies, and the zooarchaeological material recovered at these sites allows biochemical and biological investigations on historical marine ecology (Hambrecht et al., Reference Hambrecht, Anderung, Brewington, Dugmore, Edvardsson, Feeley and Gibbons2018). There are many well-preserved historical fishing sites in Iceland, and the island's geographical position, on the interface between temperate and subarctic seas, makes zooarchaeological investigations on these sites particularly suitable to examine the ecological effects of past climate fluctuations in the North Atlantic. Opposing oceanic fronts, the warm North Atlantic Current and the cold Arctic East Greenland and East Iceland Currents, result in substantial differences in sea temperature and concomitant variation in the marine environment across the island's latitudinal gradient (Ingólfsson, Reference Ingólfsson2006; Astthorsson et al., Reference Astthorsson, Gislason and Jonsson2007).
The commercialization of Atlantic cod (Gadus morhua) fisheries commenced in the North Sea in the eleventh to thirteenth centuries, expanding to the North Atlantic and then to the Newfoundland banks (Starkey et al., Reference Starkey, Reid and Ashcroft2000; Barrett et al., Reference Barrett, Orton, Johnstone, Harland, Van Neer and Ervynck2001; Barrett et al., Reference Barrett, Locker and Roberts2004b, Reference Barrett, Orton, Johnstone, Harland, Van Neer and Ervynck2011). The Atlantic cod's life history as a highly fecund, long-growing, top predator with a broad natural distribution set the scene for commercial exploitation at that scale. However, Atlantic cod distribution is sensitive to variation in water temperatures. Fisheries research has demonstrated that Atlantic cod feeding migrations, spawning distributions, and larval transport are determined by climate and oceanographic factors (Drinkwater, Reference Drinkwater2005). The Icelandic cod stock, for example, exhibited varying migration and foraging ranges in response to sea temperature changes (Astthórsson et al., Reference Astthorsson, Gislason and Jonsson2007). However, groups or populations within cod stocks commonly display varying migratory tendencies (Campana and Casselman, Reference Campana and Casselman1993; Pálsson and Thorsteinsson, Reference Pálsson and Thorsteinsson2003; Robichaud and Rose, Reference Robichaud and Rose2004; Neat et al., Reference Neat, Wright, Zuur, Gibb, Gibb, Tulett, Righton and Turner2006; Thorsteinsson et al., Reference Thorsteinsson, Pálsson, Tómasson, Jónsdóttir and Pampoulie2012). Some populations complete their life cycle in nearshore waters, whereas others engage in long-range feeding migrations, only returning to coastal waters to spawn. The different Atlantic cod migratory ecotypes may respond differently to fishing pressure and climate forcing (Árnason et al., Reference Árnason, Hernandez and Kristinsson2009; Jakobsdóttir et al., Reference Jakobsdóttir, Pardoe, Magnússon, Björnsson, Pampoulie, Ruzzante and Marteinsdóttir2011).
Biochemical and biological analysis of zooarchaeological material has been increasingly used to estimate ecological variables of past animal populations, for example, population genetic analysis (Ólafsdóttir et al., Reference Ólafsdóttir, Westfall, Edvardsson and Pálsson2014; Star et al., Reference Star, Boessenkool, Gondek, Nikulina, Hufthammer, Pampoulie and Knutsen2017), stable isotope reconstructions of historical ecology (Emslie and Patterson, Reference Emslie and Patterson2007; Misarti et al., Reference Misarti, Finney, Maschner and Wooller2009; Clark et al., Reference Clark, Horstmann and Misarti2017), and estimates of growth and age structure using otoliths (Van Neer et al., Reference Van Neer, Löugas and Rijnsdorp1999, Reference Van Neer, Ervynck, Bolle, Millner and Rijnsdorp2002; Geffen et al., Reference Geffen, Høie, Folkvord, Hufthammer, Andersson, Ninnemann, Pedersen and Nedreaas2011; Ólafsdóttir et al., Reference Ólafsdóttir, Pétursdóttir, Bárðarson and Edvardsson2017). In particular, δ13C values of marine organisms are known to reflect sea temperature, as primary production is directly affected by lower temperatures, and this is carried to higher trophic levels (Fry, Reference Fry1988). Slower growth and metabolic rates of ectotherms will also result in more variable δ13C values (Gorokhova, Reference Gorokhova2017), perhaps augmenting the effects of sea temperature. Otoliths, calcified structures of the inner ear of fish (Reibisch, Reference Reibisch1899), are occasionally recovered in archaeological excavations. Otolith annuli record the annual growth of individual fish and are often used in fish biology as measures of fish growth (Campana, Reference Campana2005) and less commonly in zooarchaeology (Van Neer et al., Reference Van Neer, Löugas and Rijnsdorp1999, Reference Van Neer, Ervynck, Bolle, Millner and Rijnsdorp2002; Geffen et al., Reference Geffen, Høie, Folkvord, Hufthammer, Andersson, Ninnemann, Pedersen and Nedreaas2011; Disspain, Reference Disspain2016; Ólafsdóttir et al., Reference Ólafsdóttir, Pétursdóttir, Bárðarson and Edvardsson2017). Fish growth is not only reflected in annulus size but also in otolith shape (Campana and Casselman, Reference Campana and Casselman1993), and otolith shape analysis is now commonly used to analyze fish ecology and environmental conditions and, in turn, to assign individual fish to stocks or subpopulations (Campana and Casselman, Reference Campana and Casselman1993; Jónsdóttir et al., Reference Jónsdóttir, Campana and Marteinsdottir2006). In both Iceland and Norway, otolith shape variation can be used to distinguish migratory and coastal Atlantic cod ecotypes (Stransky et al., Reference Stransky, Baumann, Fevolden, Harbitz, Høie, Nedreaas, Salberg and Skarstein2008; Geffen et al., Reference Geffen, Høie, Folkvord, Hufthammer, Andersson, Ninnemann, Pedersen and Nedreaas2011; Bardarson et al., Reference Bardarson, McAdam, Thorsteinsson, Hjörleifsson and Marteinsdottir2017).
Changes in the distribution and migrations of cod around Iceland in the twentieth century support predictions of similar patterns in historical times. Such climate-driven changes in migration may result in overfishing (as migratory stocks boom) and subsequent crashes in fisheries. Centennial records of climate-driven migratory shifts could therefore provide a valuable baseline for conservation and management. The substantial cooling of the North Atlantic, succeeding the Medieval Warm Period and cumulating in severe cold periods in the sixteenth to nineteenth centuries, the Little Ice Age (Ogilvie and Jónsson, Reference Ogilvie, Jónsson, Ogilvie and Jónsson2001), may have specifically resulted in changes in migrations and distributions of marine populations. Previous studies of archaeological Atlantic cod bones, for example, support population changes (Ólafsdóttir et al., Reference Ólafsdóttir, Westfall, Edvardsson and Pálsson2014) and changes in fish growth (Geffen et al., Reference Geffen, Høie, Folkvord, Hufthammer, Andersson, Ninnemann, Pedersen and Nedreaas2011; Ólafsdóttir et al., Reference Ólafsdóttir, Pétursdóttir, Bárðarson and Edvardsson2017) in the sixteenth to seventeenth centuries. In the current paper, we examine parallels in multidisciplinary proxies reflecting migration or distribution shifts in Atlantic cod, sea temperature, and concomitant shifts in the distribution and occupancy of historical fishing sites. We use biochemical, biological, archaeological, and historical analysis spanning the time since the onset of cod fisheries in Iceland ca. AD 1000 to modern times. We expect periods of low sea temperature, and the concomitant ecological effects of nearshore sea ice, lower primary production, and less prey availability, to be reflected in lower carbon stable isotope values (δ13C) of Atlantic cod and lower frequency of migratory cod in the historical catch. Moreover, we expect the occupancy of historical fishing sites to reflect our prediction of environmentally forced shifts in cod availability, with fishing sites shifting south during cold periods and establishment of northern fishing sites during warmer periods.
The history and archaeology of the Icelandic subarctic fisheries
Iceland was settled in the late ninth century by settlers who were predominantly from Scandinavia but also from the British Isles. The settlement of the island was completed by the late tenth to early eleventh centuries (Vésteinsson, Reference Vésteinsson1998, Reference Vésteinsson, Fitzhugh and Ward2000; McGovern et al., Reference McGovern, Vésteinsson, Fririksson, Church, Lawson, Simpson and Einarsson2007). During early settlement the economy was based on agriculture, that is, animal husbandry and pastoralism, and agriculture remained the main source of income until the early thirteenth century (McGovern et al., Reference McGovern, Vésteinsson, Fririksson, Church, Lawson, Simpson and Einarsson2007; Edvardsson, Reference Edvardsson2010). Archaeological evidence suggests that in the eleventh to thirteenth centuries, agriculture was encumbered by soil erosion (Þórarinsson, Reference Þórarinsson1961; Dugmore and Buckland, Reference Dugmore, Buckland, Maizels and Caseldine1991; Ólafsdóttir and Guðmundsson, Reference Ólafsdóttir and Guðmundsson2002; Dugmore et al., Reference Dugmore, Church, Buckland, Edwards, Lawson, McGovern and Panagiotakopulu2005; McGovern et al., Reference McGovern, Vésteinsson, Fririksson, Church, Lawson, Simpson and Einarsson2007), and decreasing demand for agricultural products in Europe caused prices to drop (Þorláksson, Reference Þorláksson1992; Þorsteinsson, Reference Þorsteinsson1995), which caused a decline in the Icelandic economy.
Historical and archaeological evidence suggests that subarctic fisheries in Iceland were established concurrent with the settlement of the island in the tenth century. During early settlement, most coastal farmers fished from their farms, and fishing was primarily aimed at a domestic market (Bjarnason, Reference Bjarnason1883; McGovern et al., Reference McGovern, Vésteinsson, Fririksson, Church, Lawson, Simpson and Einarsson2007; Edvardsson, Reference Edvardsson2010). The thirteenth century saw a steady increase in the demand for dried fish, that is, Atlantic cod, compelling European merchants to seek new sources in the North Atlantic, especially Norway, Iceland, and the Faroe Islands. In turn, fish prices increased rapidly, and the economy of Iceland became more reliant on fishing (Baasch, Reference Baasch1889; Þorsteinsson, Reference Þorsteinsson1995; Starkey et al., Reference Starkey, Reid and Ashcroft2000; Barret et al., Reference Barrett, Locker and Roberts2004a; Gardiner, Reference Gardiner, Barrett and Orton2016; Vésteinsson, Reference Vésteinsson, Barret and Orton2016). The early thirteenth-century expansion in commercial fishing in Iceland is recorded in the historical and archaeological record, and many new commercial fisheries were established during this period (Edvardsson, Reference Edvardsson2010). The Icelandic commercial cod fisheries thrived in the period between AD 1250 and AD 1550, as there was a steady European demand for dried fish, especially by German and English merchants (Heath, Reference Heath1968; Þorsteinsson, Reference Þorsteinsson1995; Starkey et al., Reference Starkey, Reid and Ashcroft2000).
Atlantic cod fisheries developed primarily off the west coast of Iceland, with the primary fishing grounds located off the Reykjanes and Snæfellsnes peninsulas and in the Westfjords (Fig. 1). The historical record categorizes four types of fisheries in Iceland, that is, home base (Icelandic: heimver), camp (Icelandic: viðlegurver), mixed base (Icelandic: blandað ver), and outlying base (Icelandic: útver) (Kristjánsson, Reference Kristjánsson1980). Home base was the place from which a fisher-farmer set out during fishing trips and was used generally only by himself and his farmhands. Camp is a place where many farmers from the region used a home base of a particular farm during the fishing season. Outlying base is a particular place on one's farmland, not close to the farm itself, where fishermen set out on their fishing trips during the fishing season. Mixed base is a fishing station that seems to be a mixture of the other three types of fishing sites. Archaeological research has demonstrated that the two latter types generally display preserved structures, but the former do not necessarily preserve any remaining structures (Edvardsson, Reference Edvardsson2010). Commercial fishing was mostly carried out from outlying and mixed bases, with excavations indicating that Atlantic cod were the primary targeted fish, representing 80%–90% of the catch landed. Other species, such as saithe (Pollachius virens), haddock (Melanogrammus aeglefinus), redfish (Sebastes sp.), Atlantic wolffish (Anarhichas lupus), and Atlantic halibut (Hippoglossus hippoglossus), were also targeted, primarily for domestic consumption (Amorosi et al., Reference Amorosi, McGovern and Perdikaris1994; Amundsen et al., Reference Amundsen, Perdikaris, McGovern, Krivogorskaya, Brown, Smiarowski, Storm, Modugno, Frik and Koczela2005; Harrison, Reference Harrison2014). The typical commercial fishing station consisted of fishing booths, small oval or rectangular structures made of turf where fishermen lived during the fishing season, boat landings, boathouses, winches, fish-drying platforms, and sheds. The number of structures at each site varies between regions, with the greatest number occurring on the west coast and fewer in the north and on the east coast. The general layout remained invariant, although fishing booths tend to grow larger over time (Edvardsson, Reference Edvardsson2010). The most common boat was a small open boat, powered by oars and sails, with a crew of 7–12 men; no decked vessels were used by Icelanders until the late eighteenth and early nineteenth centuries (Kristjánsson, Reference Kristjánsson1980).
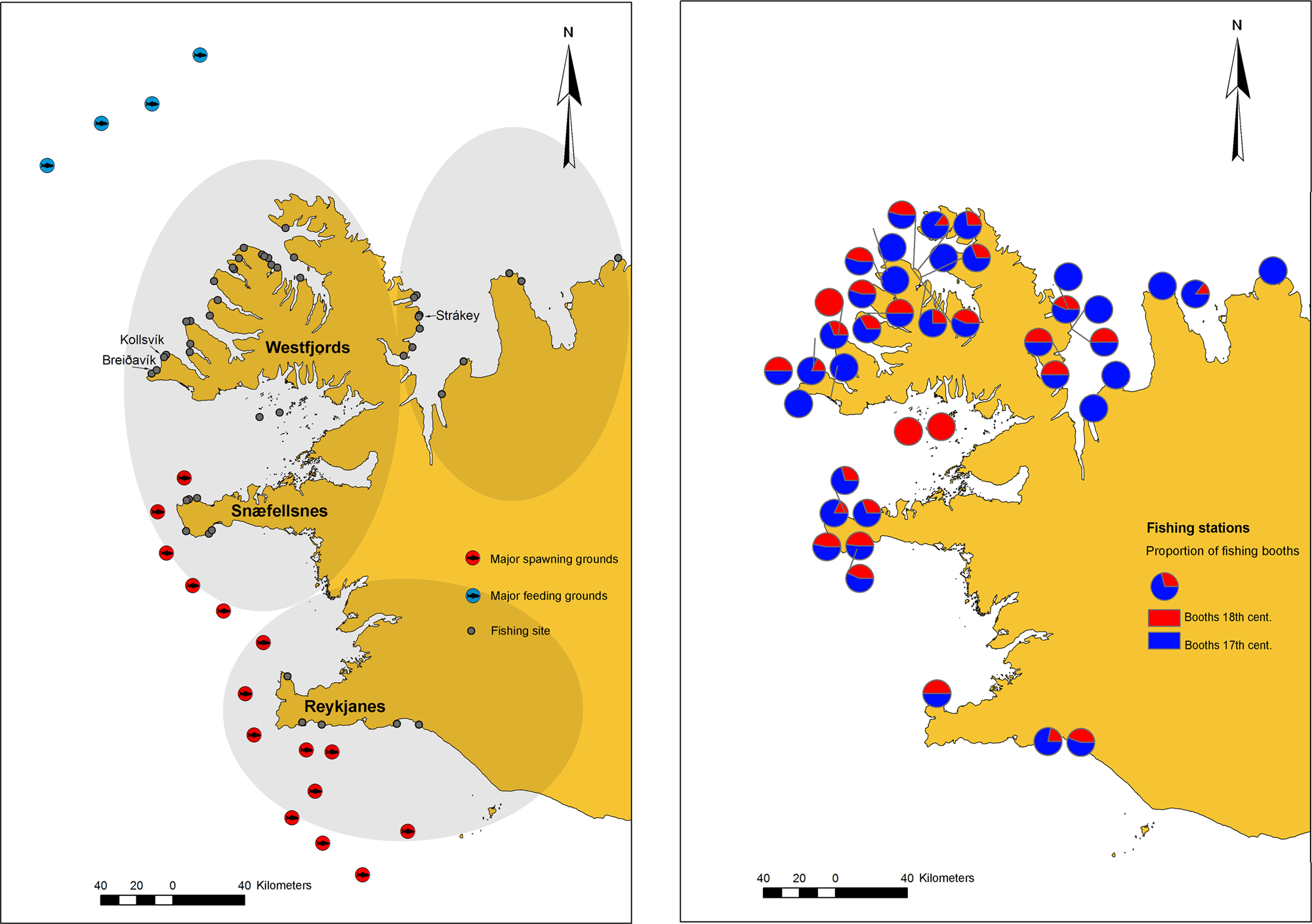
Figure 1. Left, A map showing western Iceland and the major spawning (red circles) and feeding (blue circles) grounds of Atlantic cod (note that the map excludes smaller spawning grounds found along the coast and does not depict variation in feeding grounds). Historical fishing sites are marked (gray circles), specifying the archaeological sample sites of Breiðavík (BRV), Kollsvík (KOV), and Strákey (STR). Right, A map showing changes in fishing-site occupancy between the seventeenth and eighteenth centuries as described in the land registry of Magnússon and Vídalín (Reference Magnússon and Vídalín1940).
The position and occupancy of historical fishing stations, interpreted in light of modern-day knowledge of Atlantic cod ecology (Sólmundsson et al., Reference Sólmundsson, Jónsdóttir, Ragnarsson and Björnsson2017), suggest that the stations were strategically positioned to target Atlantic cod migrations, specifically feeding migrations in the north and northwest and spawning migrations, and perhaps spawning sites, in the west and southwest.
METHODS
Archaeological data
Archaeological excavations were carried out at the fishing sites of Breiðavík (BRV) and Kollsvík (KOV) on the southern part of the Westfjords (Fig. 1). Trenches were excavated into midden deposits, and the single context method was employed. All contexts were cleaned, photographed, planned, and finally removed. The minimum size of each trench was 1 m × 1 m, although some were slightly larger. All excavated contexts were dry-sieved with a 5 mm mesh, and all finds were removed and analyzed. Fish bones were removed from the bone assemblage and analyzed separately. Terrestrial herbivore bones, when present, were used to date the deposits by 14C dating (Scottish Universities Environmental Research Centre).
A subset of the Atlantic cod vertebrae and all whole Atlantic cod otoliths were used for subsequent analysis. Statistical analysis of the zooarchaeological data was accomplished by pooling material into groups (each approximately representing one century) using 14C dates, finds, and/or stratigraphic sequence information (Table 1). In addition to the material excavated at BRV and KOV, 14 otoliths from the fishing site at Strákey (STR; Fig. 1) were included in the analysis. These otoliths were removed from contexts dated by finds to the sixteenth century (Edvardsson, Reference Edvardsson2018).
Table 1. Summary information of the zooarchaeological material used for analysis.
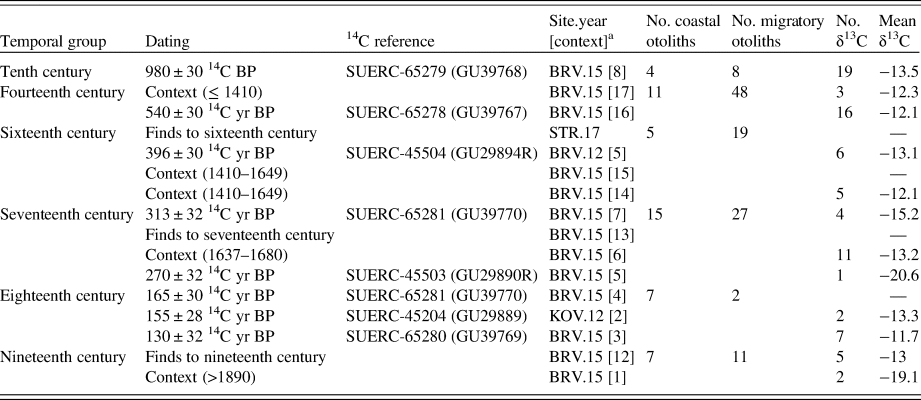
a “Site.year [context]” refers to excavation site, year excavated, and archaeological context (e.g., BRV015[8] = Breiðavík, 2015, context 8).
Stable isotope analysis
Loose sediment was removed from vertebrae by brush, while fine cleaning was performed in an ultrasonic bath (model 550D, VWR), with samples immersed in deionized water for 4 minutes. Samples were oven-dried overnight at 60°C. Dry samples were powdered in a swing mill. Collagen was extracted according to Leyden et al. (Reference Leyden, Wassenaar, Hobson and Walker2006). Stable isotope values were obtained using a Thermo Finnigan Flash 1112 EA coupled to a Thermo Finnigan Delta Plus XL via a Conflo III interface. Carbon isotope ratios were corrected for 17O contribution using the Craig correction and reported in per mil notation relative to the Vienna PeeDee Belemnite scale (VPDB). Nitrogen isotope ratios, used for quality control of the δ13C values, were reported in per mil notation relative to the Atmospheric Air Reference Scale (AIR). Carbon data were calibrated against the international standards L-SVEC (δ13C = −46.6‰VPDB) and IAEA-CH6 (δ13C = −10.45‰VPDB). Nitrogen data were calibrated against the international standards USGS-25 (δ15N = −30.4‰AIR) and IAEA-305A (δ15N = 39.8‰AIR).
Samples were tested for collagen quality by confirming that their atomic C/N ratio fell within the range of 2.9 and 3.6, resulting in 81 δ13C values used for analysis (Supplementary Table 1).
Otolith shape analysis
One hundred sixty-four otoliths could be used for shape analysis (Table 1). All otoliths used were well preserved, with no breaks or wear that could affect shape determination. Shape analysis was done “blinded,” that is, without information on 14C dating or stratigraphic sequence.
High-contrast digital images were captured using a Leica IC80 HD (Digital Camera Module by Leica Microsystems) under a stereomicroscope at 10× magnification. Background lighting was used, resulting in dark two-dimensional otolith images with bright backgrounds. The otoliths were orientated such that the rostrum was placed in the lower left corner for otoliths from the right side of the fish; for otoliths from the left side the image was flipped. The outlines were automatically traced using a black-and-white pixel gradient. Fourier descriptors (FD) were obtained from the outlines using the ‘shapeR’ (Libungan and Pálsson, Reference Libungan and Pálsson2015) and ‘lme4’ packages in R (Bates et al., Reference Bates, Mächler, Bolker and Walker2015). The otolith images are normalized in regard to the size and orientation of the otolith by fixing the first FD to 1 and the second FD to 0, respectively.
Bardarson et al. (Reference Bardarson, McAdam, Thorsteinsson, Hjörleifsson and Marteinsdottir2017) showed that otolith shape correlated with migration patterns of Atlantic cod that had been tagged with data storage tags. Specifically, otoliths from migratory cod are more streamlined or elongated compared with the more deeply shaped otoliths of the coastal ecotype (Fig. 2). Following Bardarson et al. (Reference Bardarson, McAdam, Thorsteinsson, Hjörleifsson and Marteinsdottir2017), FD3 and FD8 and three shape variables, roundness, aspect ratio, and circularity, were used as factors in a generalized linear model (GLM) to estimate the migratory value (ranging from 0 to 1) of each archaeological otolith. Values from 0 to 0.50 indicate Atlantic cod with intermediate to high migratory tendency, and values from 0.50 to 1 indicate cod with intermediate to high resident (coastal) tendency. This GLM has previously been selected as the best model for classification of otoliths to migratory ecotypes, correctly predicting the migratory ecotypes of Atlantic cod around Iceland in more than 90% of cases (Bardarson et al., Reference Bardarson, McAdam, Thorsteinsson, Hjörleifsson and Marteinsdottir2017). See Supplementary Table 2 for specifics of assignment.

Figure 2. Examples of the Atlantic cod otoliths used in the current study. The top two otoliths represent the migratory ecotype, and the lower two otoliths the coastal ecotype.
Historical analysis
To get estimates of fishing site occupancy (Fig. 1), we used the land registry of Árni Magnússon and Páll Vídalín (Reference Magnússon and Vídalín1940). This land registry, which was intended for tax purposes, was compiled in the period AD 1702–1712 by the order of the Danish king and gives the value of each individual farm and rent, the number of domestic animals, farm benefits, and other pertinent information. It contains descriptions of fishing stations, including the number of boats and booths occupied in the early eighteenth century and the number of booths and boats occupied in the seventeenth century.
Historical references to cod fishing were extracted from the Icelandic Annals (Íslenskir Annálar sive Annales Islandici ab anno Christi ad annum 1400 and Íslenskir Annálar 1400–1800, Annales Islandici Saeculorum Posteriorum). The Icelandic Annals are commonly designated as ancient annals, covering the years AD 803 to AD 1430, and later annals, covering AD 1430 to AD 1800. The ancient annals are based upon secondary written sources, whereas the more recent annals, especially in the period AD 1600–1800, were written exclusively about contemporary events. The annals cover different topics of current interest, including deaths of eminent people, natural disasters, weather, political events, and fishing. The annals, therefore, offer historical documentation of economic and natural events that affected the lives and livelihood of local people. All references to Atlantic cod fishing were extracted. For a comparable data set on fishing (although not including weather or sea ice) in AD 1800–1878, we used the published compilation of Bjarnason (Reference Bjarnason1883). The references were assigned to a region (north, west, or south) following the designation of regions shown in Figure 1, and all references to bad weather or sea ice hindering fishing were noted.
Statistical analysis
To test whether the Atlantic cod vertebrae δ13C values (reflecting lifetime temperature exposure) varied between centuries, we used a Kruskal-Wallis test with post hoc Mann-Whitney U-tests with Benjamini-Hochberg correction for multiple comparisons. To test whether the proportion of migratory-type otoliths differed between centuries, we used a two proportions z-test.
To examine variation in the number of references to good, average, or bad fishing between centuries and regions we used a GLM (general linear model). The number of references formed the dependent variable, with region, type of references (good, bad, average), and temporal group (century) being the fixed effects, with interaction between both the type of effect and temporal group and region and temporal groups (the size of the data set did not allow a model that estimated all interaction effects). All statistical analysis was done in R v. 3.4.2 (R Core Team, 2017) using RStudio v. 1.0.153 (RStudio Team, 2015).
RESULTS
Stable isotope analysis
The Atlantic cod vertebrae δ13C values varied between −10.0‰ and −20.6‰ and had a mean of −13.1‰. These values are akin to previously published values from Atlantic cod zooarchaeological material (Orton et al., Reference Geffen, Høie, Folkvord, Hufthammer, Andersson, Ninnemann, Pedersen and Nedreaas2011). There was significant variation in δ13C values between centuries (Kruskal-Wallis χ2= 22.10, df = 5, P < 0.001) (Fig. 3). Post hoc Mann-Whitney U-tests with Benjamini-Hochberg correction showed that the δ13C values were significantly lower in the tenth, seventeenth, and nineteenth centuries, reflecting cooler sea temperatures and/or lower primary production in the cod environment, than in the fourteenth century (P < 0.01).

Figure 3. The box plots show δ13C values from Atlantic cod vertebrae in the historical catch in each of the temporal groups, specifically the median (horizontal black line) and first (box) and third (whiskers) quartiles. Outliers are presented as black circles. The δ13C scale is on left y-axis. The frequency of coastal otoliths in the historical catch is depicted as figures of otoliths, with the scale on the right y-axis. Reference to sea ice or bad weather hindering fishing in the Icelandic annals (before AD 1800) are depicted as light blue vertical lines. Finally, reconstructed summer (blue line) and winter (red line) sea-surface temperatures off North Iceland (from Jiang et al., Reference Jiang, Eiríksson, Schulz, Knudsen and Seidenkrantz2005) are shown in the background.
Otolith shape analysis
Most of the archaeological Atlantic cod otoliths were assigned to the otolith shape characteristic of the modern-day migratory cod type (Table 1). Migratory otolith shape was predominant in the sample until the seventeenth century (Fig. 3, Table 1). The percentage of otoliths assigned to the migratory otolith shape was significantly lower in the seventeenth, eighteenth, and nineteenth centuries (36%, 78%, and 40%, respectively) than in the fourteenth and sixteenth centuries (19% and 21%, respectively) (z = 2.88, df = 1, P < 0.01), suggesting that at the peak of historical fisheries, migratory cod was much more prevalent in the catch. The percent of migratory otoliths in the tenth century (33%) did not differ significantly from other periods.
Historical analysis
Data from the eighteenth-century land registry clearly signal a decline in the number of fishing booths, particularly in the north, between the seventeenth and eighteenth centuries (Fig. 1). From a total of 404 general references to Atlantic cod fishing in the Annals, 232 could be assigned to either the northern, western, or southern regions (Table 2). The results from the GLM show that references to fishing were more frequent in west Iceland than in north Iceland and that references are most frequent in the eighteenth century (Table 3). References to bad fishing were least likely to refer to west Iceland, and references to good fishing increased in the nineteenth century (Table 2). A total of 16 references were made to sea ice or bad weather hindering cod fishing, most commonly in the eighteenth century (Fig. 3).
Table 2. Overview of the number of regional references to Atlantic cod fishing in each century.

Table 3. Results from the generalized linear model depicting estimates and significance of type of reference (good and bad vs. average) and geographical region (with an interaction term) and type of reference and century (with an interaction term).a

a Significant estimates, at the 0.05 level, are in bold type.
DISCUSSION
Carbon stable isotope values (δ13C) reflect rates of primary production and photosynthetic fractionation (Fry, Reference Fry1988) and are generally lower in poikilotherms inhabiting colder waters (Ramos and González-Solís, Reference Ramos and González-Solís2012). Carbon stable isotope values also correlate to metabolic rates in poikilotherms (Wurster and Patterson, Reference Wurster and Patterson2003; Wurster et al., Reference Wurster, Patterson, Stewart, Bowlby and Stewart2005) and may reflect growth rate and food source limitation (Gorokhova, Reference Gorokhova2017). The current results of lower δ13C values in the seventeenth and nineteenth centuries (Fig. 3) are consistent with either decreased sea temperature and slower growth (related to lower temperature decreasing metabolism) or, alternatively, shifts in Atlantic cod migration to colder water that would result in the same effect. The lower δ13C values coincide with recorded cold periods and harsh climate conditions in the North Atlantic (Ogilvie and Jónsdóttir, Reference Ogilvie and Jónsdottir2000; Dahl-Jensen et al., Reference Dahl-Jensen, Mosegaard, Gundestrup, Clow, Johnsen, Hansen and Balling1998) (Fig. 3), supporting a hypothesis of local cooling.
Other variations in the current δ13C data, for example, the relatively low values in the tenth century and high values in the eighteenth century, may reflect fluctuations rather than consistent cooling during the Little Ice Age (Jiang et al., Reference Jiang, Eiríksson, Schulz, Knudsen and Seidenkrantz2005; Patterson et al., Reference Patterson, Dietrich, Holmden and Andrews2010). Studies of seasonality as well as winter and summer sea-surface temperatures on the Iceland shelf suggest earlier and more severe fluctuations of summer temperature at the onset of the Little Ice Age (Jiang et al., Reference Jiang, Eiríksson, Schulz, Knudsen and Seidenkrantz2005; Patterson et al., Reference Patterson, Dietrich, Holmden and Andrews2010). In particular the summer sea-surface temperatures of the North Iceland shelfs appear to coincide well with the current δ13C data (Fig. 3). Moreover, the δ13C data represent lifetime values of a highly mobile species. Migratory cod engage in northern feeding migrations and are exposed to colder waters than coastal cod (Pálsson and Thorsteinsson, Reference Pálsson and Thorsteinsson2003; Thorsteinsson et al., Reference Thorsteinsson, Pálsson, Tómasson, Jónsdóttir and Pampoulie2012). However, an alternative hypothesis, that the lower δ13C values represent not lower local sea temperatures but increases in migration and migratory cod in the catch, is contradicted by the observed lower frequency of migratory-type otoliths in seventeenth to nineteenth-century catches (Fig. 3).
Both migratory and coastal populations of Atlantic cod are common at the northern distribution range of the cod distribution, such as around Iceland (Robichaud and Rose, Reference Robichaud and Rose2004). The archaeological otolith analysis showed that migratory-type otoliths in the catch were most frequent in the fourteenth to sixteenth centuries, at the peak of land-based Atlantic cod fisheries in Iceland (Fig. 3). The lower proportion of migratory cod to coastal cod in the catch after AD 1600 may reflect three nonexclusive mechanisms: (1) population changes, for example, lower frequency of migratory cod in the Atlantic cod stock; (2) climate effects, such as climate forcing of Atlantic cod migration patterns; or (3) weather conditions that hindered fishers from targeting the cod migrations but had no effect on migration patterns or population composition. Certainly, historical references suggest that conditions for fishing were often unfavorable during the eighteenth and early nineteenth centuries and that sea ice was common in coastal regions (Fig. 3). However, the north Iceland Atlantic cod fisheries deteriorated before any increase in references to weather detrimental to fishing (Fig. 3). Modern-day fisheries research has documented reoccurring shifts in the migration patterns of fish species in Icelandic waters in response to temperature change (reviewed in Valdimarsson et al., Reference Valdimarsson, Astthorsson and Palsson2012). Feeding migrations may be particularly flexible, as the Atlantic cod responds to changes in both sea temperature and prey distribution; this happened during a warm period of the twentieth century when foraging Atlantic cod followed the Arctic capelin north (Astthorsson et al., Reference Astthorsson, Gislason and Jonsson2007). Shifts in feeding migrations may well explain the archaeological and historical data presented in the current paper, that is, during periods of significant cooling, the Atlantic cod foraging grounds north of Iceland were no longer reliable fishing grounds. On the other hand, the large spawning grounds and associated spawning migrations in the west and southwest (Sólmundsson et al., Reference Sólmundsson, Jónsdóttir, Ragnarsson and Björnsson2017) were more consistently favorable.
In addition to climate-driven shifts in migration routes, latitudinal shifts in the spawning grounds of migratory cod in response to sea temperature change are known to have occurred in the twentieth century, for example, when Atlantic cod that originated in Iceland started spawning along the Greenland coast (Bonanomi et al., Reference Bonanomi, Pellissier, Therkildsen, Hedeholm, Retzel, Meldrup and Olsen2015). Such shifts may explain the significantly lower frequency of migratory cod in the local catch. Ólafsdóttir et al. (Reference Ólafsdóttir, Westfall, Edvardsson and Pálsson2014) showed lowered frequency of the Pan IB allele, characteristic of migratory cod (Fevolden and Pogson, Reference Fevolden and Pogson1997; Pampoulie et al., Reference Pampoulie, Ruzzante, Chosson, Jörundsdóttir, Taylor, Thorsteinsson, Daníelsdóttir and Marteinsdóttir2006), in zooarchaeological Atlantic cod material from Iceland after AD 1550. The same study also reported lowered genetic diversity, which again supports fishing from a truncated Atlantic cod stock after AD 1550.
The current analysis identifies the proximity to rich fishing grounds as key to the development of the Icelandic commercial fisheries during the medieval period. The locations of fishing sites (Fig. 1) suggest that fishers targeted feeding and spawning migrations, and potentially spawning grounds, of Atlantic cod and that these sites provided the most consistently favorable fishing throughout the period. However, environmental change resulted in fisher movement on varying scales. Small-scale changes, such as coastal erosion destroying boat landings, forced fishermen to relocate to new sites, often in close proximity to earlier locations, which the archaeological record suggests occurred at different times in the northwest of Iceland (Edvardsson, Reference Edvardsson2010). On a larger scale, depletion of in-fjord fishing grounds forced many fisher-farmers to abandon their home bases and move to the larger commercial stations to fish (Magnússon and Vídalín, Reference Magnússon and Vídalín1940). Large-scale changes, such as reoccurring lack of catch over long periods of time, forced fishermen to relocate nationally to fish seasonally, and from the late seventeenth century, fishermen from the north of Iceland traveled to the commercial fishing stations in the southwest (Kristjánsson, Reference Kristjánsson1980; Þór, Reference Þór2002). The current historical analysis shows that commercial fishing stations in North Iceland had been abandoned long before the eighteenth century and replaced by opportunistic subsistence fishing by local farmers. By the early eighteenth century, the number of boats and booths at commercial fishing stations in the Snæfellsnes peninsula and the Westfjords had dropped by 50%–60%, and several sites had been abandoned (Fig. 1), but the abandonment of fishing sites was less pronounced in the south (Fig. 1). After the seventeenth-century collapse, the number of booths and boats does not begin to increase again until the early nineteenth century, and by the mid-nineteenth century, the number of booths exceeded their original number in most regions (Kristjánsson, Reference Kristjánsson1980). This is also reflected in the data set from Bjarnason (Reference Bjarnason1883), which shows increased references to favorable fishing in the nineteenth century (Table 2).
Although the current multidisciplinary results support a significant role for climate-driven ecological shifts in a historical collapse of Atlantic cod fisheries in Iceland, the fisheries cannot be examined solely within an environmental context. The earliest written sources concerning European fishers in Icelandic waters date to the late fourteenth century (Íslenskir Annálar, 1847). From the period AD 1408 onward, the English sent large fishing fleets to Iceland annually. Other nations followed, primarily the Dutch, French, and Germans, and many European nations had fishing fleets in Icelandic waters by the late nineteenth century (Bjarnason, Reference Bjarnason1883; Þorsteinsson, Reference Þorsteinsson1976, Reference Þorsteinsson1995; Simon-Thomas, Reference Simon-Thomas1935; Pálmadóttir, Reference Pálmadóttir1989; Jones, Reference Jones, Starkey, Reid and Ashcroft2000; Starkey et al., Reference Starkey, Reid and Ashcroft2000). Icelandic fishers frequently protested to local authorities about foreign fleets ruining fishing grounds and fishing gear (Bjarnason, Reference Bjarnason1883), suggesting that local fishermen attributed reduced Atlantic cod catches to the presence of European fishing fleets. The European fishing fleets may in fact have had the advantage of mobility, allowing them to respond to changing migrations and moving stocks, while the Icelandic fishing boats were limited to a fixed point (i.e., a fishing station) and had a limited sphere of operation. The Icelandic fishers had to risk the costly wait for the annual migrations, and when the migrating fish did not materialize in a particular area, fishers were forced to move to a new site. The relative certainty of the Atlantic cod catches around Iceland, procured during consistent annual spawning migrations, secured the early success of the land-based subarctic fisheries, but ultimately the local fisheries did not adapt sufficiently for Icelandic cod fisheries to flourish in a changing climate and an increasingly competitive environment. These results add to accumulating evidence for successful colonization and exploitation of subarctic environments in the Viking Age, followed by the creation of a flexible subsistence system (Simpson et al., Reference Simpson, Dugmore, Thomson and Vésteinsson2001; Dugmore et al., Reference Dugmore, Keller and McGovern2007, Reference Dugmore, McGovern, Vésteinsson, Arneborg, Streeter and Keller2012; McGovern et al., Reference McGovern, Vésteinsson, Fririksson, Church, Lawson, Simpson and Einarsson2007), but then the eventual failure to adapt an established economy to technological and climate-driven changes.
To conclude, the current results show that lower sea temperatures in the North Atlantic coincided with lower lifetime ambient temperatures of Atlantic cod and lower frequency of migratory cod in the Icelandic Atlantic cod catches. These results support a shift in Atlantic cod migration routes or, more drastically, migratory cod spawning grounds. These ecological changes, perhaps in concert with increased international fishing pressure, appear to have resulted in a temporary collapse of Atlantic cod fisheries in Iceland, despite local fishers responding with increased mobility.
ACKNOWLEDGMENTS
We would like to acknowledge Tom McGovern for his conceptual input in developing this work and Ben Fitzhugh, David Orton, and James Barrett for useful comments on this article. We thank Leifur Þór Þorvaldsson, Gylfi Björn Helgason, Zuhaitz Akizu, Marion Durand, Kjartan Ari Theoódórsson, Benedikt Kári Theódórsson, and Emma Lilja Ragnarsdóttir for field assistance and Einar Pétur Jónsson for his work on the otolith shape assignment.
SUPPLEMENTARY MATERIAL
To view supplementary material for this article, please visit https://doi.org/10.1017/qua.2018.147









 Open access
Open access