


INTRODUCTION
Oil palm is the most important source of edible oil, accounting for 0·30 of global production of oils and fats (Oil World 2015). Cultivation of oil palm has expanded over the last few decades, especially in Southeast Asia (Indonesia and Malaysia). There are two species of oil palm, Elaeis guineensis which originated from Central and West Africa and E. oleifera which originated from South and Central America. E. guineensis is the species commercially cultivated in Southeast Asia.
Lipases (triacylglycerol acylhydrolases) play an important role in lipid metabolism, regulating the synthesis and hydrolysis of lipids. They catalyse the hydrolysis of ester compounds, primarily neutral lipids such as triacylglycerols (TAG) at the oil and water interface, resulting in the release of long chain aliphatic acids from glycerol (Brockman Reference Brockman, Borgstrom and Brockman1984). Incomplete hydrolysis results in the release of monoacylglycerols (MAG) and diacylglycerols (DAG) in addition to free fatty acids (FFA).
Studies have confirmed the presence of active endogenous lipases in various crops including oats (Hoi et al. Reference Hoi, Holland and Hammond1999), rice bran (Goffman et al. Reference Goffman, Pinson and Bergman2003), wheat seed (Pierozan et al. Reference Pierozan, da Costa, Antunes, Oestreicher, Oliveira, Cansian, Treichel and de Oliveira2009) and oil palm (Sambanthamurthi et al. Reference Sambanthamurthi, Chong, Oo, Yeo and Rajan1991, 1995). Palm oil which originates from the mesocarp is the most important vegetable oil, accounting for almost half of the world's edible oil (Singh et al. Reference Singh, Ong-Abdullah, Low, Manaf, Rosli, Nookiah, Ooi, Ooi, Chan, Halim, Azizi, Nagappan, Bacher, Lakey, Smith, He, Hogan, Budiman, Lee, DeSalle, Kudrna, Goicoechea, Wing, Wilson, Fulton, Ordway, Martienssen and Sambanthamurthi2013). However, the high oil content in the mesocarp makes it a rich substrate for lipase activity. Oil palm lipase activity is synchronized with the biosynthesis of palm oil, first detected at ~16 weeks after anthesis (WAA) and reaching its peak at 21 WAA (Sambanthamurthi et al. Reference Sambanthamurthi, Oo and PArman1995). Lipase in the mature palm mesocarp hydrolyses TAG to FFA, MAG and DAG (Sambanthamurthi et al. Reference Sambanthamurthi, Rajanaidu and Parman2000a ) and since FFA contribute to acidity and rancidity of the oil, oil palm genotypes with low lipase activity are desirable in order to produce oil with increased shelf-life. Conversely, genotypes with high lipase activity may be an attractive source of lipase for fat splitting to produce FFA and glycerol for the production of detergents and other industrial products.
The Malaysian Palm oil Board (MPOB) has the largest ex situ collection of natural oil palm populations in the world, with materials from most of its provenances including Nigeria, Cameroon, Democratic Republic of Congo (Zaire), Tanzania, Madagascar, Angola, Senegal, Gambia, Sierra Leone, Guinea and Ghana (Rajanaidu et al. Reference Rajanaidu, Jalani, Kushairi, Rao, Rajanaidu and Jalani1999). The germplasm collection is a rich genetic resource for oil palm breeding and serves to broaden the genetic base of the present breeding material, which is extremely narrow.
The MPOB germplasm collection has been screened extensively using isozymes and molecular markers such as restriction fragment length polymorphism (RFLP), amplified fragment length polymorphism (AFLP) and simple sequence repeats (SSR). Studies using SSR markers have shown a high level of genetic divergence in the germplasm and strong association between genetic distance and geographical location. Higher diversity was also observed in the germplasm material compared with a standard commercial Deli dura population, demonstrating the utility of the germplasm material in crop improvement (Rajanaidu et al. Reference Rajanaidu, Jalani, Kushairi, Rao, Rajanaidu and Jalani1999).
The present study aimed to screen the MPOB oil palm germplasm collection for lipase activity using a simple and accurate method. Oil palm lipase is activated at low temperature (Sambanthamurthi et al. Reference Sambanthamurthi, Chong, Oo, Yeo and Rajan1991, Reference Sambanthamurthi, Oo and PArman1995) and a novel method of screening for lipase activity developed by Sambanthamurthi et al. (Reference Sambanthamurthi, Chong, Oo, Yeo and Rajan1991, Reference Sambanthamurthi, Oo and PArman1995) was independently validated by Cadena et al. (Reference Cadena, Prada, Prerea and Romero2013). A total of 148 palms from various countries of origin were screened. The assay was subsequently used for screening the germplasm.
MATERIALS AND METHODS
Plant materials
Ripe fruits from several E. guineensis palms from the MPOB oil palm germplasm collection at its research station in Kluang, Malaysia were sampled. One bunch was selected per palm and ten spikelets sampled per bunch. A total of 148 palms originating from eight countries, namely Angola, Cameroon, Guinea, Nigeria, Senegal, Sierra Leone, Tanzania and Zaire were used. Of the 148 palms, 128 were grouped into 34 populations comprising 33 families. The 20 remaining palms did not fall into any population or family but were useful for analysis at country level. In all cases, ripe oil palm fruits were used and lipase activity of the mesocarp measured. All assays comprised at least three technical replicates.
Assay for lipase activity using the radioactive method
Freshly harvested fruits were sterilized in 10% Clorox™ solution for 10 min and washed thoroughly in sterile distilled water. Five to ten mesocarp slices were cut from the fruits and the exocarp discarded. The slices were ground to a fine powder in a mortar with liquid nitrogen using a pestle. A mixture comprising 100% ethanol, 5% gum Arabic, 5% 3-[{3-cholamidopropyl} dimethylammonio]-1-propanesulfonate (CHAPS), 1% Tween20 and phosphate buffer pH 6·8 containing 5% bovine serum albumin and 12 mm dithiothreitol (DTT) in the proportion 1 : 1 : 1 : 1 : 2 was vortexed for 5 min to a smooth emulsion. The emulsion had to be freshly prepared as it is only stable for up to 2 h. Two µl 27·3 nmol [14C] trioleate substrate were added to a clean eppendorf tube and 35 µl of the emulsion were added to the tubes containing the radioactive substrate. Then, 10 mg of the cold mesocarp powder were added quickly to the tubes. The mixtures were vortexed to a homogenate and incubated at ambient temperature (26 °C) for 30 min. The reaction was stopped by adding 650 µl methanol : chloroform : hexane in the ratio 1·41 : 1·25 : 1·00, repectively. For the blank, the stop reagent was added before the mesocarp powder to prevent any lipolysis. The [14C] oleic acid released by the lipase activity was converted into potassium salt by adding 210 µl 50 mm potassium carbonate (K2CO3) containing 0·1 m potassium hydroxide (KOH), pH10. The tubes were vortexed, then centrifuged for 60 s and 200 µl of the upper aqueous layer counted in a scintillation counter.
Assay for lipase activity using the cold-activated titration method
Freshly harvested fruits were stored at low temperature (4 °C) for 18 h to activate lipase. The fruits were then sterilized in 10% Clorox™ solution for 10 min and washed in sterile distilled water. They were then autoclaved for 10 min at 10 pounds per square inch (psi) to deactivate the lipase and soften their mesocarp. The oil was extracted by physical pressing and 1 g of oil was dissolved in 40 ml neutralized isopropanol. The mixture was warmed to about 40 °C to ensure that the oil was fully dissolved and then titrated with 0·1 M sodium hydroxide (NaOH) (AOCS Reference Firestone1998). For the samples with extremely low FFA contents, 2 g oil were used. The FFA content in the palm oil was expressed as a percentage based on palmitic acid (AOCS 1998).
Correlation study and data analysis
Comparison of lipase assay using radioactive and non-radioactive cold activation method
A total of 49 palms were randomly selected for carrying out both the conventional radioactive and non-radioactive cold activation (titration) methods to test for correlation of the two methods.
Data analysis
Analysis for population and family differences was done using SPSS 15·0 (Argyrous Reference Argyrous2011). Analysis of variance (ANOVA) was carried out to evaluate the significance of differences between and within populations and families of the material.
Differences between and within groups are outlined below:
 $$\eqalign{& \sigma _z^2 = \sigma _z^2 /\sigma _t^2 \times 100\% \cr & \sigma _z^2 = \sigma _w^2 /\sigma _t^2 \times 100\% \cr & \sigma _t^2 = \sigma _z^2 + \sigma _w^2} $$
$$\eqalign{& \sigma _z^2 = \sigma _z^2 /\sigma _t^2 \times 100\% \cr & \sigma _z^2 = \sigma _w^2 /\sigma _t^2 \times 100\% \cr & \sigma _t^2 = \sigma _z^2 + \sigma _w^2} $$
where
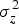 $\sigma _z^2 $
= between group variance,
$\sigma _z^2 $
= between group variance,
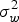 $\sigma _w^2 $
= within group variance and
$\sigma _w^2 $
= within group variance and
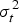 $\sigma _t^2 $
= total variance.
$\sigma _t^2 $
= total variance.
Heritability is the ratio of genetic to total phenotypic variance (Falconer & Mackay Reference Falconer and MacKay1996). The genetic variance is estimated from the variance between families. The total phenotypic variance is contributed by both genetic and environmental factors. Environmental variance is estimated from the variance within families. In this case, heritability was computed using intra-class correlation, as shown below.
Intra-class correlation,
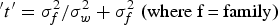 $$'t' = {\sigma_f^2} / {\sigma_w^2} + {\sigma_f^2} \;\hbox{(where f = family)}$$
$$'t' = {\sigma_f^2} / {\sigma_w^2} + {\sigma_f^2} \;\hbox{(where f = family)}$$
The broad sense Heritability is 2t if the seedlings are full-sibs and 4t if the seedlings are half-sibs.
Effect of long-term storage of the fruits from selected oil palm germplasm on the free fatty acid contents of their mesocarp oil
Fruits found to have low- and high-lipase activity were analysed for the FFA contents of their mesocarp oils after long-term storage at ambient temperature (26 °C). The fruit spikelets were left on a laboratory bench for up to 2 weeks. Every 2 days, a few fruits from the spikelets were tested for their mesocarp oil FFA content by titration. The fruits were not exposed to low temperature at all, as the aim was only to ascertain how long-term storage at ambient temperature affects FFA in their mesocarp oil.
Lipase expression in palms exhibiting different levels of lipase activity
Palms from four different levels of lipase activity (material from Angola, Guinea, Sierra Leone and Zaire) were selected. A modified method of Prescott & Martin (Reference Prescott and Martin1987) was used for RNA extraction: 2 µg RNA samples were reverse-transcribed using MultiScribe Reverse Transcriptase, available in the High-Capacity cDNA Archive Kit (Applied Biosystems). Real time polymerase chain reaction (PCR) was carried out on the first strand cDNA generated in a Mastercycler® ep realplex (Eppendorf, Hamburg, Germany). The 25 µl reaction mixture contained 1 µg cDNA (first strand) in 4 and 21 µl of 1 × TaqMan® Universal PCR Master Mix containing 0·2 µm TaqMan® Probe (Applied Biosystem) and 1·25 µm reverse and forward primers in wells of a MicroAmp optical 96-well Reaction Plate (Applied Biosystem, Foster City, CA, USA). Real-time PCR was carried out following PCR protocol: Uracil-N-glycosylase activation: 50 °C for 2 min (1 cycle); DNA polymerase activation: 95 °C for 10 min (1 cycle); amplification: 95 °C for 15 s, 60 °C for 1 min (40 cycles). For each 96-well plate, non-template and non-reverse transcription controls were included and tested concurrently with the samples. A calibrator sample was selected and included in every plate during the experiment. All the reactions were carried out in triplicate. Three MPOB in-house reference genes (Chan et al. Reference Chan, Rose, Abdul Murad, Zainal, Low, Ooi, Ooi, Yahya and Singh2014) and two genes of interest, GDSL and GXSXG lipases of the oil palm, were used. The expression stability of these genes was determined using the geNorm algorithm (Vandesompele et al. Reference Vandesompele, De Preter, Pattyn, Poppe, Van Roy, De Paepe and Speleman2002). All normalization and validation for quantitative real time PCR (qPCR) complied with the Minimum Information for Publication of qPCR Experiments (MIQE) guidelines. Quantification analysis was based on the formula E = 10−1/slope (Pfaffl Reference Pfaffl2001). In the present study, the relative expression ratio was calculated based on the expression of the lipase genes v. a calibrator in comparison with expression of the reference gene selected. At least four biological replicates were done for each of the four levels of lipase activity. All the reactions were carried out in triplicate (technical replicates)
RESULTS
Comparison of lipase assay using radioactive and non-radioactive cold activation methods
In total 49 palms were randomly selected for carrying out both the conventional radioactive and non-radioactive cold activation (titration) methods. The data obtained from the 49 palms were transformed to their Log values to minimize their variance and plotted together as shown in Fig. 1. The results from both assay methods were significantly correlated with a Pearson correlation r-value of 0·529 (P < 0·01). With the significant correlation, the less expensive and simpler non-radioactive cold-activation titration was used for the subsequent screening.

Fig. 1. Correlation of results obtained from radioactive and non-radioactive cold-activation methods. X-axis represents free fatty acid (FFA) content (percentage) following exposure of fruits at 4 °C for 18 h. Y-axis represents the radioactive count as measured in a scintillation counter.
Lipase activity in oil palm germplasm collection
Lipase activity was investigated in oil palm germplasm at the country, population, family and individual levels. The detailed results are given in Supplementary Table 1 (available from http://journals.cambridge.org/AGS). A range of lipase activity was observed. The overall FFA percentage of the 148 palms collected from eight countries was, however, skewed towards low levels with a mean of 16·82% under cold-activation (Fig. 2). Most of the low lipase palms were from Guinea, Nigeria and Senegal with mean FFA values <10% under cold activation while palms from Angola, Tanzania and Zaire had higher activity with mean FFA values >20%. This distribution pattern was studied using a histogram for individual palms. The distribution was skewed to the left for Guinea, Nigeria and Senegal, i.e. the three countries where palms had the lowest percentage of FFA in their oils. On the other hand, the distribution was skewed to the right for palms from Angola, Tanzania and Zaire (Figs 3(a)–(h)). On an individual level, Guinea (palm number 03·04) had the lowest lipase activity, exhibiting FFA content of only 0·26% even under cold activation. Conversely, Zaire (palm number 33·17) had highest lipase activity among the palms studied, showing FFA content of 65·15% under cold activation.

Fig. 2. Frequency distribution of free fatty acid (FFA) content in mesocarp oil from 148 palms from eight countries of origin. X-axis represents FFA content following exposure of fruits at 4 °C for 18 h. Y-axis represents the frequency of palms in each class.

Fig. 3. Distribution of free fatty acid (FFA) content in mesocarp oil from palms from different countries of origin: (a) Angola (b) Cameroon (c) Guinea (d) Nigeria (e) Senegal (f) Sierra Leone (g) Tanzania and (h) Zaire) X-axis represents FFA content following exposure of fruits at 4 °C for 18 h. Y-axis represents the frequency of palms in each class. The best fitted distribution curve is also shown.
The ANOVA showed significant differences between countries. The variance component between countries contributed 26·6% to the total phenotypic variance (Table 1). A multiple comparison of means was made using least significant difference (LSD). In the present study, the LSD value was 9·63 for the 148 palms. The highest mean values of 26·16 and 26·84% were observed for Tanzania and Zaire, respectively. The lowest were for Nigeria (2·55%), Guinea (7·99%) and Senegal (8·38%) (Table 2).
Table 1. Analysis of variance (ANOVA), mean square and variance components for 148 palms from eight countries of origin

Table 2. Means, range of free fatty acids (FFA) and least significant difference (LSD) of 148 palms from eight countries of origin

LSD = 9·6331.
The ANOVA results also showed significant differences between populations. The variance between populations
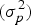 $(\sigma _p^2 )$
contributed nearly 40% to the total phenotypic variation (Table 3), implying that selection can be done at the population level. The overall FFA mean was 16·93% with LSD value of 20·50. For example, population Senegal 02 with FFA value of 1·50% was much lower than Sierra Leone 13 (40·99%), Sierra Leone 14 (39·66%) and Tanzania 04 (37%) (Table 4). The present study showed wide genetic variation in oil palm lipase activity at country and population levels.
$(\sigma _p^2 )$
contributed nearly 40% to the total phenotypic variation (Table 3), implying that selection can be done at the population level. The overall FFA mean was 16·93% with LSD value of 20·50. For example, population Senegal 02 with FFA value of 1·50% was much lower than Sierra Leone 13 (40·99%), Sierra Leone 14 (39·66%) and Tanzania 04 (37%) (Table 4). The present study showed wide genetic variation in oil palm lipase activity at country and population levels.
Table 3. Analysis of variance (ANOVA), means, mean squares and variance components for 34 populations

Table 4. Means, range of free fatty acids (FFA) and least significant difference (LSD) of 34 populations

ANG, Angola; CAM, Cameroon; GUI, Guinea; NGR, Nigeria; SEN, Senegal; TAN, Tanzania; ZRE, Zaire.
LSD = 20·5.
The ANOVA results also showed significant differences between families. Heritability for lipase activity was estimated by the variance component of 33 families (Table 5a ). The overall FFA mean was 15·15% with LSD value of 20·70. The family means showed Guinea family 04·03 to have the lowest (Mean FFA: 0·45%) and Tanzania family 11·01 the highest lipase activity (Mean FFA: 56·24%) (Table 5b ). The heritability estimate for lipase activity was high (Table 5c ), indicating the possibility of selecting for lipase activity at the family level as well.
Table 5. Analysis of variance (ANOVA), Means, mean squares and variance components for 148 palms from different families

variance components are based on ANOVA Table 5a.
Effect of long-term storage on fruits from palms exhibiting varying levels of lipase activity
Fruits from Guinea (palm number 0·353/190) and Nigeria (palm number 0·399/34), (palm number 0·399/36) exhibiting low lipase activity and Angola (palm number 0·311/607) having high lipase activity were left at ambient temperature for 2 weeks and analysed for FFA every 2–3 days (by sampling different fruits). The FFA content was initially low but increased considerably, reaching a peak on days 7–9, thereafter remaining approximately constant (Fig. 4). It then resumed its gradual increase, reaching 8–14%, at which stage the fruits were rotting. The Guinea palm 190, which exhibited low lipase activity based on the cold activated titration method, exhibited the slowest increase. The Nigerian palm which had low lipase activity also showed a slow rise in FFA during long-term storage. Conversely, the Angolan material, which was identified to have high lipase activity, showed the steepest increase.

Fig. 4. Effect of long-term storage of oil palm fruits on the free fatty acid (FFA) content of their mesocarp oil. (AT 607 – Angola palm 0·311/607; GD190 – Guinea palm 0·353/190; ND34 – Nigeria palm 0·399/34; ND36 – Nigeria palm 0·399/s6). X-axis represents number of days that ripe oil palm spikelets were left on the bench prior to FFA analysis. Y-axis represents percentage of FFA.
Lipase expression in palms exhibiting various levels of lipase activity
Oil palm contains at least two lipase classes, GXSXG and GDSL (unpublished data). The quantitative PCR data indicate that the gene expression levels of both the GDSL and GXSXG genes are in agreement with the lipase activity. Angola palms, which had the highest FFA content, also exhibited highest GDSL and GXSXG expression. Similarly, Guinea palms, which had the lowest FFA content, also had the lowest expression for both GDSL and GXSXG genes (Fig. 5). The PCR amplification efficiencies are given in Supplementary Fig. 1 (available from http://journals.cambridge.org/AGS).

Fig. 5. GDSL and GXSXG lipase expression in palms with different levels of lipase activity. X-axis shows the countries of origin of palm with different lipase activities. Y-axis shows the relative expression of GDSL(■) and GXSXG genes (![]() ) genes based on the expression of the lipase gene in a calibrator in comparison with expression of the reference genes selected.
) genes based on the expression of the lipase gene in a calibrator in comparison with expression of the reference genes selected.
DISCUSSION
Previous studies have used both radioactive and non-radioactive assays to determine lipase activity in oil palm (Henderson & Osborne Reference Henderson and Osborne1991; Sambanthamurthi et al. Reference Sambanthamurthi, Chong, Oo, Yeo and Rajan1991, Reference Sambanthamurthi, Oo and PArman1995, Reference Sambanthamurthi, Sundram and Tan2000b ). In the conventional radioactive method, 14C oleic acid (or another fatty acid, depending on the substrate) is released by hydrolysis of the radio-labelled lipid substrate. The 14C oleic (or other) acid released is converted to a potassium salt, which is then separated from the non-reacted substrate and counted in a liquid scintillation counter (Sambanthamurthi et al. Reference Sambanthamurthi, Oo and PArman1995). However, the radioactive method is difficult, tedious and expensive. It also requires extra safety precautions. In the non-radioactive approach, lipase activity is calculated from the percentage FFA released from the oil by titration with NaOH (AOCS Reference Firestone1998). The method is convenient, but less sensitive and accurate and only reliable with appreciable quantities of FFA, i.e. when lipase activity is high. It is not sensitive enough to detect FFA at low concentrations. When 0·1 M NaOH is used as titrant, it is not sufficiently sensitive to detect activity < 0·1 µmol/min (Beisson et al. Reference Beisson, Tiss, Rivière and Verger2000). In the present study, the accuracy was improved by exposing the fruits to low temperature as oil palm lipase is activated at low temperature. This enhanced titration was used to screen the germplasm as the results had been shown to correlate well with the results from radioactive assay. Although the level of correlation fell to a moderate level (0·529), it was nonetheless statistically significant (P < 0·01).
Previous genetic diversity studies in oil palm using both phenotypic and molecular markers showed significant differences between countries, populations and families (Singh et al. Reference Singh, Zaki, Ting, Rosli, Tan, Low, Ithnin and Cheah2008; Ting et al. Reference Ting, Zaki, Rosli, Low, Ithnin, Cheah, Tan and Singh2010; Zaki et al. Reference Zaki, Singh, Rosli and Ismail2012; Arias et al. Reference Arias, González, Prada, Restrepo and Romero2013). Similar genetic structure was observed in the present study for lipase activity.
Ecogeographical distribution, owing to the different environments, contributes to the phenotypical adaptation and differences in plants (Allard Reference Allard and Tigerstedt1996). The results of the present study indicate that the ecogeographic variation of the population may have contributed to the variation in lipase activity. Nigeria, Cameroon, Zaire and Tanzania are just above the equator, while Angola is south of it. Senegal, Guinea and Sierra Leone are on the Northern coastal line and the west of Africa. From the results, the low lipase palms are from the northern-most oil palm areas (Nigeria, Guinea, Sierra Leone and Senegal), bordering the Sahara desert which are generally low-lying, whereas the high lipase palms are from Angola, Tanzania and Zaire (now known as Democratic Republic of Congo), which have higher rainfall. As lipolysis is a hydrolysis reaction involving the participation of water, the palms probably adapted to the rainfall pattern and it is not surprising that higher lipase activity was observed in palms originating from countries with higher rainfall while those from drier regions exhibited lower lipase activity. Furthermore, the lower temperatures at the higher elevation may also have contributed to adaptation of palms. This pattern of ecogeographic variation in lipase activity is corroborated by genetic variation of natural populations of the palms (with the exception of Nigerian palms) across the same region assayed using isozymes (Hayati et al. Reference Hayati, Wickneswari, Maizura and Rajanaidu2004) and a number of DNA based markers viz. AFLP (Kularatne Reference Kularatne2000), RFLP (Maizura et al. Reference Maizura, Rajanaidu, Zakri and Cheah2006) and SSR (Bakoumé et al. Reference Bakoumé, Wickneswari, Siju, Rajanaidu, Kushairi and Billotte2015). Earlier studies using SSR markers had shown a high level of genetic divergence in the germplasm and strong association between genetic distance and geographical location. It was reported that the E. guineensis germplasm material was divided into two clusters – material originating from Madagascar and those from the African mainland (Rajanaidu et al. Reference Rajanaidu, Jalani, Kushairi, Rao, Rajanaidu and Jalani1999; Hayati et al. Reference Hayati, Wickneswari, Maizura and Rajanaidu2004). The mainland germplasm was subsequently further divided into a few sub-clusters (Singh et al. Reference Singh, Zaki, Ting, Rosli, Tan, Low, Ithnin and Cheah2008; Ting et al. Reference Ting, Zaki, Rosli, Low, Ithnin, Cheah, Tan and Singh2010). Generally the studies showed that the populations from drier countries such as Senegal and Gambia were clustered together compared with humid countries such as Nigeria, Cameroon and Zaire. The present study on lipase showed a similar trend. Human intervention may also have affected the lipase pattern in different countries. Although low lipase is preferred for producing quality oil for commercial use, certain tribes in Africa favour high FFA oil for culinary purposes. Thus, there may have been selection for high lipase in specific sites in Africa.
Taking into consideration the wide variation in oil palm lipase activity at country and population levels, a key question was whether the observed variation was due to environmental or genetic factors. Heritability is a concept that describes how much of the variation in a trait is attributable to genetic factors. Oil palm is largely cross-pollinated. The seedlings derived from a bunch are considered an open-pollinated family. The pollination may be from a single or multiple sources of pollen. Thus, the heritability estimation can be complicated. In the present study, an intra-class correlation was used to compute heritability (Falconer & Mackay Reference Falconer and MacKay1996). If an inflorescence was pollinated by a single male, the seedlings would be full-sibs (2t) and if by multiple pollen sources, half-sibs (4t). The heritability estimate computed was very high, irrespective of whether they were assumed to be half- or full-sibs. The high heritability estimates suggest that the lipase activity is under strong genetic control, allowing detection of stable quantitative traits loci or functional markers for lipase activity. This paves the way for selection and breeding for low and high lipase palms.
In the study on the effect of long-term storage on fruits from palms exhibiting varying levels of lipase activity, the initial gradual increase in FFA that reached a peak before plateauing off was caused by the endogenous oil palm lipase(s). Any increase in FFA thereafter was probably caused by microbial lipase(s) (fungus), as visually evidenced by the presence of fungus on the fruits. Sambanthamurthi et al. (Reference Sambanthamurthi, Oo and PArman1995) previously showed that endogenous mesocarp lipase was the main contributor to acidity in the oil palm mesocarp and fungal lipases only play a role once the fruits start rotting. The recent findings by Morcillo et al. (Reference Morcillo, Cros, Billotte, Ngando-Ebongue, Domonhédo, Pizot, Cuéllar, Espéout, Dhouib, Bourgis, Claverol, Tranbarger, Nouy and Arondel2013) are in agreement with this. The palms identified in the present study as having low lipase activity showed low increases on long-term storage, while those with high lipase activity exhibited higher increases. In producing palm oil, the fruit bunches have to be transported from the field to the mills for processing. The post-harvest handling and processing of the oil-rich palm fruits would result in increased contact between the lipase and fat substrate, thus resulting in increased hydrolysis of TAG to FFA. To minimize FFA release, these bunches have to be heat-sterilized and processed quickly. Delays in transporting and processing the fruits would result in an appreciable increase in FFA. The results obtained in the present study indicate that the low lipase palms would not encounter such drastic increases. While low lipase palms are desirable for maintaining the quality of the oil for marketing, nevertheless, high lipase palms have specific uses, for example, the high lipase palm from Zaire 33·17 showing 65% FFA would be highly valuable for fat splitting in the chemical industry.
The presence of at least two lipase classes in the oil palm, GXSXG and GDSL, was recently confirmed (unpublished data). The GXSXG lipase is mesocarp-specific, and expressed only in the later developmental stages of the fruit, while GDSL is expressed in the leaf, root and mesocarp with higher expression at the early developmental stages of the fruit. The present results suggest that both lipases are involved in release of FFA in the oil palm. However, since high FFA occurs when the oil palm fruits are injured or exposed to low temperature, it is suggested that both bruising and low temperature can trigger stress-responsive gene(s) in the palm, in this case, the GDSL lipase. In addition, bruising will release lipases, bringing them into contact with the substrate. Morcillo et al. (Reference Morcillo, Cros, Billotte, Ngando-Ebongue, Domonhédo, Pizot, Cuéllar, Espéout, Dhouib, Bourgis, Claverol, Tranbarger, Nouy and Arondel2013) recently reported that acidity in the oil palm mesocarp is controlled by a single gene. However, the present data suggest that both the GDSL and GXSXG lipases are at play. The GDSL lipases are involved in regulation and defence under extreme conditions, e.g. regulation of Oxytropis (an arctic plant) under cold treatment (Archambault & Stromvik Reference Archambault and Stromvik2011), as well as systemic resistance signalling by supressing auxins, which are an important disease susceptibility factor in Arabidopsis (Lee et al. Reference Lee, Kim, Kwon, Jin and Park2009).
In conclusion, the non-radioactive low temperature titration method is a convenient and accurate tool for measuring lipase activity in oil palm mesocarp. There were significant differences in lipase activity in palms from different countries of origin, populations and families. Palms with low and high lipase activities were identified. The three palms with the lowest FFA values were from Nigeria, Senegal and Guinea, whereas the highest were palms from Angola, Tanzania and Zaire. The MPOB collection comprises wild oil palms from the diverse regions of Africa. The present study showed wide genetic variation in oil palm lipase activity at country and population levels. Heritability estimates were very high irrespective of whether they were assumed to be half or full sibs. This paves the way for selection and breeding for low- and high-lipase palms. In selecting for palms of desired lipase phenotype, yield also has to be taken into consideration.
SUPPLEMENTARY MATERIAL
To view supplementary material for this article, please visit http://dx.doi.org/10.1017/S0021859615001112.
The authors thank the Director-General of Malaysian Palm Oil Board, Datuk Dr Choo Yuen May for permission to publish this paper and the Breeding Group of Malaysian Palm Oil Board for providing samples.













 Open access
Open access