



The chicken (Gallus gallus domesticus) is the most widespread domestic food animal of our time. It is one of the most efficient farm animals for protein production—more so than dairy cattle (Redding Reference Redding2015). Yet, chickens were not always used for meat and eggs. When introduced to the Americas, domesticated fowl were often used for feathers, for cockfighting, and as pets (Pitt et al. Reference Pitt, Gillingham, Maltby and Stewart2016; Sykes Reference Sykes2012). Their use for meat consumption is a fairly recent innovation in many regions of the world. In Japan, for example, there is little evidence that chickens were consumed regularly before the nineteenth century (Sykes Reference Sykes2012). In other parts of the world, such as Central Europe and the Pacific Islands, zooarchaeological and ethnographic data indicate that the primary motivation for introducing chickens to these regions was for meat but probably not for eggs (Benecke Reference Benecke1993; Langdon Reference Langdon1989).
Chickens were domesticated from wild jungle fowl (Gallus gallus) in Asia, and introduced to the New World by European settlers (Thomson et al. Reference Thomson, Lebrasseur, Austin, Hunt, Burney, Denham, Rawlence, Wood, Gongora, Flink, Linderholm, Dobney, Larson and Cooper2014). Neither domestic chickens nor wild jungle fowl have been recovered from prehistoric North American sites. According to Reitz and colleagues (Reference Reitz, Scarry and Seifert1985:28), chickens arrived in what is now the southeastern United States with Spanish soldiers during the sixteenth century. According to Lawler (Reference Lawler2014:193), they arrived in the northeastern United States (e.g., Jamestown) with British settlers in the early seventeenth century.
Historical accounts indicate that in the first two decades after their establishment (ca. AD 1769–1790), Upper California presidios and missions were regularly provisioned with goods and livestock from Mexico (Forbes Reference Forbes1839:11; Geiger Reference Geiger1963; Hackel Reference Hackel1997; Palóu Reference Palóu1926:356; Serra Reference Serra and Tibesar1955:361). In an effort to ensure livestock (e.g., cattle, sheep) proliferation, the Spanish government largely forbade slaughter of these animals (Archibald Reference Archibald1978:177; Cheever Reference Cheever1983:40). When California missions became economically viable, agricultural surplus was provisioned to presidio military personnel against their wages (Archibald Reference Archibald1978:11; Forbes Reference Forbes1839:145; Serra Reference Serra and Tibesar1955:365). While the Spanish crown promoted self-sufficiency among missions and presidios, California presidios do not appear to have reached the goal of agricultural sustainability (Aiken Reference Aiken1983:9; Bancroft Reference Bancroft1888:240; Cheever Reference Cheever1983:47; Hittell Reference Hittell1897:507). In light of the economic uncertainty and irregular food supply, small-scale poultry husbandry practiced at the presidios would have provided soldiers and their families a form of economic independence. Although historical documentation does not indicate whether chickens and eggs played an integral part of the provisioning network, zooarchaeological analysis of San Diego Presidio chicken remains sheds notable light on this question.
Our knowledge of chicken husbandry in colonial North America is fairly limited. Analyses of chicken remains from North American colonial sites commonly discuss only their relative frequency (Blind et al. Reference Blind, Voss, Kenton and Barker2004; Lucido Reference Lucido2013; Lyman Reference Lyman1977; Reitz et al. Reference Reitz, Scarry and Seifert1985:67; Schulz et al. Reference Schulz, Quinn and Fulmer1987; Schweitzer Reference Schweitzer2010; Simons Reference Simons and Felton1984; Szuter Reference Szuter and Orser1996). Questions related to male/female ratios, age profiles, or whether chickens were managed for their eggs have not been addressed. In contrast to North America, chicken remains from Classical and post-Classical periods in Europe and the ancient Near East are well studied. The archaeological literature is accessible and allows comparative inferences about chicken husbandry through time and space (Boessneck and von den Driesch Reference Boessneck and von den Driesch1979; De Cupere et al. Reference De Cupere, Van Neer, Monchot, Rijmenants, Udrescu and Waelkens2005; Gál Reference Gál, Jerem, Mester and Réka2006; Kyselý Reference Kyselý2010; Maltby Reference Maltby1997; Moreno-García and Detry Reference Moreno-García, Detry, Prummel, Zeiler and Brinkhuizen2010; Perry-Gal et al. Reference Perry-Gal, Erlich, Gilboa and Bar-Oz2015; Redding Reference Redding2015; Slavin Reference Slavin2009; Van Neer et al. Reference Van Neer, Noyen, De Cupere and Beuls2002).
As discussed below, historical data on chickens are also sparse. While large livestock animals—such as cattle, horses, mules, and sheep—are commonly enumerated in California mission inventories, and information about their age category and sex is often available (e.g., the number of mares, colts, calves, or lambs), this is not the case with chickens (Archibald Reference Archibald1978:159; Bancroft Reference Bancroft1888:337; Forbes Reference Forbes1839:265–266; Palóu Reference Palóu1926:216–217; Richman Reference Richman1911:31).
Chicken remains from the San Diego Presidio represent the earliest known utilization of domestic fowl on the western coast of what is now the United States. These remains serve as an excellent case study for addressing the following questions: What role did chickens play in the colonial Spanish subsistence economy, particularly at the time of their introduction to the Pacific coast of North America? Were the presidios provisioned with poultry, or did they raise and manage them on-site? Were chickens systematically raised for meat, eggs, or both? Using zooarchaeological methods and the limited historical accounts available, our study attempts to provide the first comprehensive assessment of chicken husbandry in Upper California.
Although the zooarchaeological sample available for this study is relatively small (n = 790), the number of identified species is strikingly high, with 20 avian and 14 mammalian taxa represented (Table 1; Supplemental Table 1). The number of identified chicken bones is also relatively high, compared to those identified from other presidio and mission sites (Benté et al. Reference Benté, Tordoff and Hilderman-Smith1982; Pavao-Zuckerman Reference Pavao-Zuckerman2011; Voss Reference Voss2008:307–324; Walker Reference Walker, Costello and Gasco1985). For instance, while nearly 56,000 animal bones are reported from various zooarchaeological studies of the San Francisco Presidio, only 157 chicken bones were identified among them (Smith-Lintner Reference Smith-Lintner2007; Valente Reference Valente, Ramsay and Voss2002a, Reference Valente, Ramsay and Voss2002b; Wake Reference Wake1996). In contrast, the San Diego Presidio yielded 303 chicken bones, making this sample especially useful for evaluating chicken husbandry in the late eighteenth to early nineteenth century (Table 2). Although it remains unclear why the number of chicken remains from other sites is relatively low, Table 2 demonstrates that chicken bones from the San Diego Presidio represent one of the largest samples to date and warrant comprehensive study.
Table 1. Avian NISP, MNI, and Modification Marks.
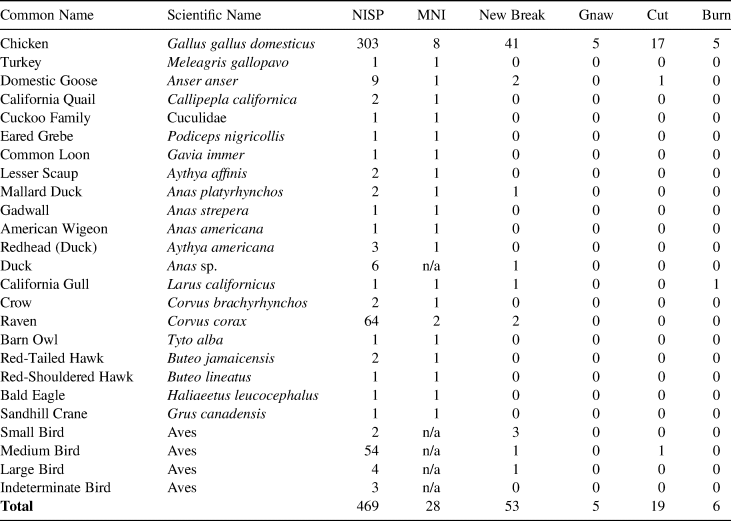
Table 2. Chicken Bones in California and Arizona Presidios and Missions.
Site Background
Established in AD 1769, the San Diego Royal Presidio was the earliest European settlement along the western coast of the future United States (Figure 1). Constructed on a hill, overlooking San Diego Bay and the San Diego River, it was the first of four military colonies (San Diego, Monterey [1770], San Francisco [1776], and Santa Barbara [1782]) established in the province of Upper (Alta) California (Voss Reference Voss2008:54–65; Williams Reference Williams2004a). The San Diego Presidio was a fortified military base for only two-thirds of its history (ca. AD 1780–1821). Along with Spanish military officers and Mexican soldiers, the garrison population included many civilians, representing an ethnically mixed community with a large number of Native American people. At its height, the outpost sheltered more than 500 inhabitants, and by AD 1790, it served as the chief administrative and judicial center for the surrounding region. The presidio of San Diego was abandoned in AD 1835 (Williams Reference Williams1997, Reference Williams2004b).
Figure 1. San Diego Presidio location. (Color online)
In the first decade of its history, the San Diego Presidio was unwalled. About 10 years after its establishment, it was rebuilt as a fortified adobe quadrangle with a chapel, areas assigned to soldiers' barracks, a commandant's house, a guardhouse, homes for civilians and troops with dependents, and a residential area for visiting padres (Figure 2; Williams Reference Williams2004b).
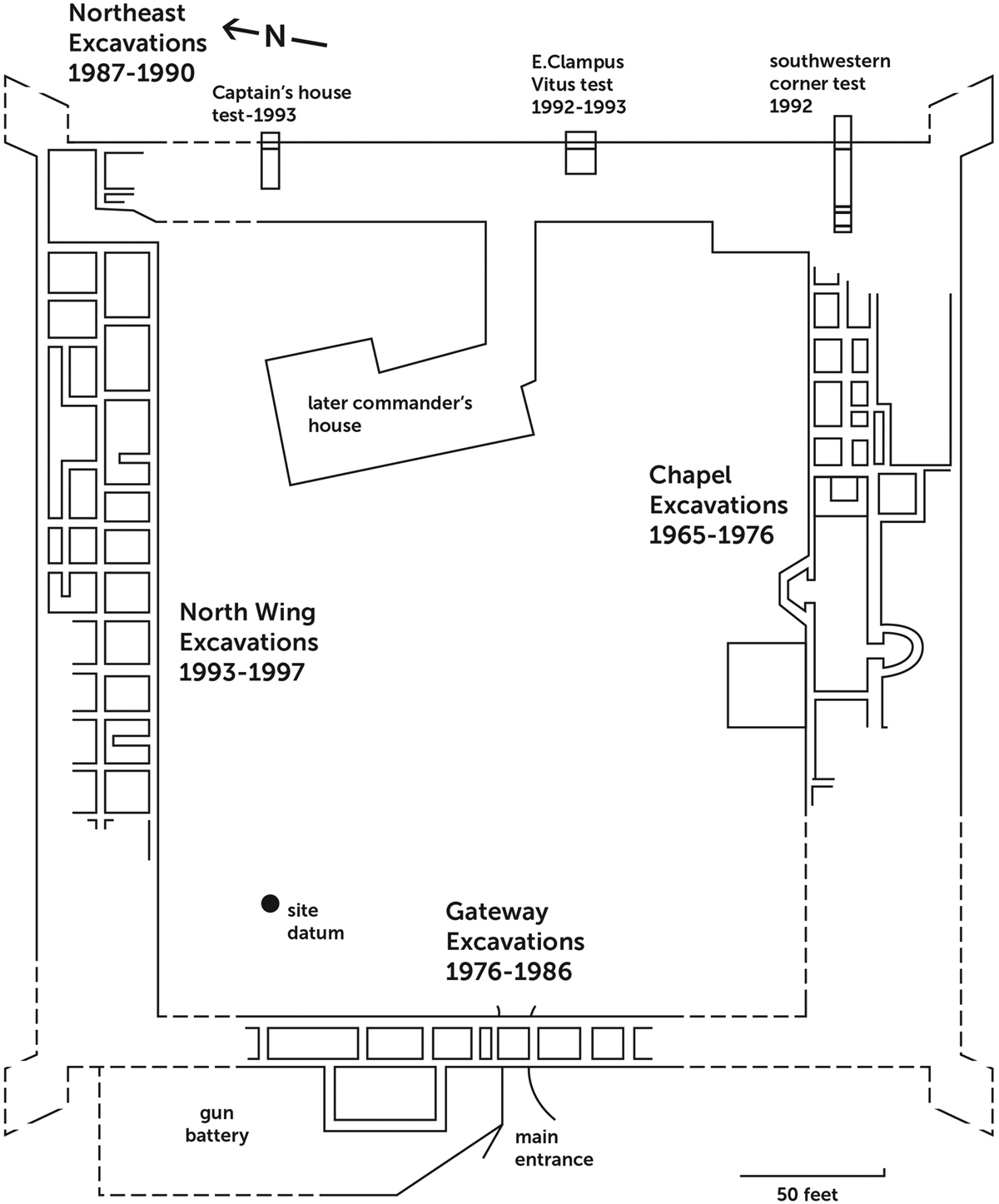
Figure 2. Primary excavation areas at San Diego (based on Williams Reference Williams1997).
Between 1927 and 1997, six different archaeological expeditions excavated the San Diego Presidio (Williams Reference Williams1997). In the years 1965–1976, the chapel and the southern fortification wall were excavated by Paul Ezell (Reference Ezell1976). Between 1976 and 1983, Diane Barbolla (Reference Barbolla1992) excavated the western fortification wall. A large midden outside the west wing of the fortification was also excavated. In the late 1980s, Brad Bartel (Reference Bartel1991) excavated the northeast corner of the presidio, and in the late 1990s, the north wing's fortification wall and residential quarter were excavated by Jack Williams (Reference Williams1997, Reference Williams2004b). The north wing served as a residential area with over 20 rooms and associated yards. The west wing consisted of two areas: a block of residential rooms within the western wall and a midden over 4 m deep that is outside the western wall. The midden very likely served as the foundation for a gun battery built between AD 1791 and 1810 (Barbolla Reference Barbolla1992; Williams Reference Williams1997, Reference Williams2004b). Most of the faunal remains we examined were recovered from the north and west wings.
Only partial results of some of the excavations described above have been published. To date, no synthesis of excavation results has been carried out, nor has a comprehensive archaeological report on the San Diego Presidio been published. Only two zooarchaeological studies from the west and north wings of the San Diego Presidio are known to us. Over 2,600 large-mammal remains from the west wing were studied by Cheever (Reference Cheever1983). According to this study, cattle constituted 72% of the identified elements, 6% were sheep, and 1.3% were deer. Over 4,000 animal bones were recovered from Bartel's (Reference Bartel1991) excavation of the presidio's northeast corner. Of these, approximately 280 bone fragments (7%) were identified to species, with 25% representing cattle, 26% fish, 24% rodents, and 12% birds (Bartel Reference Bartel1991). Bird taxa were not specified.
Materials and Methods
Retrieval of the Bone Assemblage
In the late 1990s, the City of San Diego closed the presidio excavations, and the site was backfilled. Apart from the chapel excavations (Ezell Reference Ezell1976), all presidio artifacts and ecofacts reside with the City of San Diego facility. Pending a comprehensive and costly inventory and cataloging of the presidio material culture, scholarly research remains on hold. Apart from faunal studies concurrent with past excavations (Bartel Reference Bartel1991; Cheever Reference Cheever1983), no analysis or research on material, stored at the City of San Diego facility, has been conducted since the late 1990s.
A brief window opened in 2011, and the City of San Diego allowed the authors, among other scholars, access to the presidio storage facility for a limited time. During this time, we retrieved faunal material for analysis. The total number of presidio faunal specimens has yet to be determined. The authors’ visual inspection of dozens of artifact storage boxes puts an estimate of specimens in the tens of thousands. The faunal sample bags were stored within mixed-content artifact boxes (e.g., bone, glass, ceramics). The authors searched through as many boxes as time allowed, locating and pulling sample bags containing avian remains. In addition to bird bones, many of these bags contained other classes of fauna. We studied these specimens too, and for this reason, we briefly describe the non-avian remains. In the search for chicken remains among over 400 storage boxes, we sought to include specimens from the two primary excavations: the west wing (Barbolla Reference Barbolla1992) and north wing (Williams Reference Williams1997, Reference Williams2004b). Specimens from the northeast corner of the presidio were also collected (Bartel Reference Bartel1991). As such, bird remains in our study come from all primary areas of the west wing (rooms within the compound and a deep midden outside the walls) and from 11 rooms within the north wing. Information on screening methods during excavations was only available from the west wing, where all soil was screened through 3 mm hardware mesh (Barbolla Reference Barbolla1992). No information was provided on the use of screening methods in the excavation summaries of the north wing or northeast portion of the site (Bartel Reference Bartel1991; Williams Reference Williams1997, Reference Williams2004b).
In light of the goal of this study—to provide insights into chicken husbandry at the San Diego Presidio—it should be noted that, pending stratigraphic study, dates that might delineate chronological components or phases of the site are unavailable. Therefore, our analysis may encompass the entire occupation period of the San Diego Presidio (AD 1769–1835).
Research Methods
The zooarchaeological study was carried out at the San Diego Zooarchaeology Laboratory using the osteological reference collection housed in the San Diego Natural History Museum. Anatomical skeletal atlases were also consulted (Cohen and Serjeantson Reference Cohen and Serjeantson1996; Miles et al. Reference Miles, Martin and Savage1985). Common and scientific names of animal species mentioned in the text follow Wilson and Reeder (Reference Wilson and Reeder2005) and Myers et alia (Reference Myers, Espinosa, Parr, Jones, Hammond and Dewey2018). Specimens were identified to species, skeletal element, and side (left or right) when possible. Limb bone fragments were identified as proximal end, distal end, or shaft. Specimens unidentifiable to species or skeletal element were categorized by size. Small birds include those the size of most passerines, medium birds the size of ducks and chickens, and large birds the size of geese or larger.
The assemblage was quantified using two common zooarchaeological methods: NISP (number of identified specimens) and MNI (minimum number of individuals; Reitz and Wing Reference Reitz and Wing2008:202–204). Specimens were examined with a magnifying lens (×40) or digital microscope (×60) for evidence of bone modifications resulting from fire, butchering, weathering, and animal gnawing. New breaks (fractures) incurred during archaeological excavation, transport, or storage were also recorded. Cracked bones and splinters were not counted as new breaks (Sasson Reference Sasson2010:62).
Differential representation of avian skeletal elements in archaeological sites may reflect cultural practices or natural (i.e., taphonomic) processes (Bovy Reference Bovy2012; Ericson Reference Ericson1987). We tested both notions within the historic archaeological context of the San Diego Presidio. The cultural hypothesis was assessed by examining body-part representation against the relative frequency of chicken leg and wing bones. The taphonomic hypothesis was tested by plotting the relative frequency of skeletal elements and their relative density (see discussion below). Dirrigl (Reference Dirrigl2001) scanned and measured areal bone mineral density (BMDa) and volume bone mineral density (BMDv) of the wild turkey (Meleagris gallopavo). He scanned three areas (“scan site”) for each specimen: proximal, distal, and medial (shaft). For our analysis of density-mediated attrition, we included complete long bones as well as fragments of proximal ends, distal ends, and shafts. Following Dirrigl's protocol, we assigned complete long bones to the medial category. For example, we counted a complete femur as one medial femur for the bone-density analysis (Supplemental Table 2). We used Dirrigl's data on turkey-bone density because, to date, no data on chicken-bone density are available. Density-mediated attrition is tested by comparing the relative density of scan sites (e.g., proximal femur versus distal femur). We used data on turkey-bone density under the premise that the relative density of turkey bone at the sites scanned (e.g., distal femur) correlates with that at equivalent sites in the chicken. This supposition is based on Latimer's studies, which showed that the relative weights of long bones of domestic chickens correlate with those of turkey hens (Latimer Reference Latimer1927; Latimer and Rosenbaum Reference Latimer and Rosenbaum1926).
Ageing and Sexing
The stages of limb bone fusion serves as one way to document a chicken's age at death. In the chicken, true bony epiphyses (secondary ossification centers) occur only at the proximal end of the tarsometatarsus and at the proximal and distal ends of the tibiotarsus. However, cartilaginous growth plates are present at both ends of every long bone from 16 days after hatching until growth is complete 30 weeks later (Church and Johnson Reference Church and Johnson1964; Latimer Reference Latimer1927; Serjeantson Reference Serjeantson2009:39). For assessing relative age at death, we categorized long bones as either fused (mature) or unfused (immature; Supplemental Table 3). Additionally, the outer cortical bone of hatchlings and subadult fledglings retains a visibly porous outer texture (Fothergill et al. Reference Fothergill, Best, Foster and Demarchi2017; Gál Reference Gál, Jerem, Mester and Réka2006; Serjeantson Reference Serjeantson2009:38–40; Thomas et al. Reference Thomas, Sadler and Cooper2014). We categorized these specimens as immature also.
Well-developed, fully calcified bone spurs generally occur only on the distal shaft of the tarsometatarsus of roosters and castrated capons. We examined all tarsometatarsi for the presence or absence of bone spurs. It should be noted that spurs may occasionally fail to develop in males, and they may occur as an underdeveloped, oval knob in hens by one year of age. Additionally, in productive egg-layers, normal hormonal inhibition of spur growth may be reversed during the phase of the cycle following reproduction, resulting in spur growth (Sadler Reference Sadler1991; Serjeantson Reference Serjeantson2009:47–49; West Reference West, Wilson, Payne and Grigson1982).
On average, hens are considerably smaller than roosters, although the sexes do overlap in size. We used osteometric data and statistical models to identify size variations between males and females, and possibly, different breeds. Following von den Driesch (Reference von den Driesch1976), we measured the distal breadth (Bd, Did), proximal breadth (Bp, Dip), and greatest length (GL) of chicken skeletal elements. Only fully ossified, unburnt fragments were measured (Supplemental Table 4). To assess size variation in the osteological measurements, we applied two statistical methods. In the first phase, we used the “log-ratio” method to pool measurements from different skeletal elements (Fothergill et al. Reference Fothergill, Best, Foster and Demarchi2017; Haber and Dayan Reference Haber and Dayan2004; Payne and Bull Reference Payne and Bull1988; Simpson et al. Reference Simpson, Roe and Lewontin1960:356–358). With this method, the logarithm of the ratio between the size of a zooarchaeological and a standard (modern) specimen is calculated. The standard skeleton used for this analysis was a domestic rooster from Arizona that died in 1977. Its measurements are presented in Supplemental Table 4. For example, if the Bp measurement of an archaeological femur is 15.5 mm and the standard measurement of this element is 17.2, the log-ratio between 15.5 and 17.2 is calculated as log10 (15.5/17.2) = 0.0459. Davis (Reference Davis1996) has shown that most measurements taken along the same axis—be it length, width, or depth—are quite highly correlated, but measurements taken along different axes tend to be less correlated. Following Davis, we compiled two log-ratio datasets for statistical modeling: one of the greatest length (length axis) and the other of the greatest breadth (breadth axis) of the proximal and distal portions of long bones.
In the second phase, we modeled the two datasets using “mixture analysis.” This technique can be used on samples that consist of a mixture of two or more different component populations (such as males and females), allowing individuals to be assigned to one or the other of the mixed components. In other words, in osteometric datasets, mixture analysis modeling detects and highlights the males, females, and in some cases, smaller breeds. It is based on the method of maximum likelihood and estimates the parameters for each group (mean, proportion, and standard deviation; De Cupere et al. Reference De Cupere, Van Neer, Monchot, Rijmenants, Udrescu and Waelkens2005; Fernández and Monchot Reference Fernández and Monchot2007; for further details, see Dong Reference Dong1997; Monchot and Léchelle Reference Monchot and Léchelle2002). Statistical analysis was performed in PAST (Hammer et al. Reference Hammer, Harper and Ryan2001).
Medullary bone is unique to female birds and crocodilians, and during eggshell formation, it serves as a source of stored calcium. This secondary bone is generated in varying densities within most skeletal elements and is readily visible in the open medullary cavities of a hen's long bones. Medullary bone forms during the pre-laying period of approximately 7–14 days, and it is resorbed after the hen ceases laying (Canoville et al. Reference Canoville, Schweitzer and Zanno2019; Church and Johnson Reference Church and Johnson1964; Rick Reference Rick1975; Whitehead Reference Whitehead2004). Zooarchaeological chicken limbs containing medullary bone reflect the presence of hens, most likely slaughtered at the end of their egg-laying cycle (Gál Reference Gál, Jerem, Mester and Réka2006; Kyselý Reference Kyselý2010; Rick Reference Rick1975; Van Neer et al. Reference Van Neer, Noyen, De Cupere and Beuls2002). We inspected all avian long bones in our study for medullary bone but analyzed only those from chickens. Proximal and distal ends of limbs that only retained a small fragment of the shaft were excluded from the analysis. In addition to examining substantive bone-shaft fragments, we sectioned 40 complete long bones through the middle of the shaft area and inspected them for medullary bone after they were measured, weighed, and photographed.
Results
The faunal remains analyzed in this study originated from three areas of the San Diego Presidio, with 75% of the bone assemblage recovered from the west wing, 21% from the north wing, and 4% from the northeast corner inside of the presidio walls. Although the focus of this article is chicken husbandry, data on other species represented in the bone assemblage warrant mention.
In total, 790 animal bones were studied. Of these, 469 were avian, representing 20 species—17 wild and 3 domesticated. Domestic poultry account for over half of all bird specimens, most of which are chicken (Table 1). Since the bone assemblage for this study was collated with chicken bones as the primary focus, the scant representation of goose and turkey remains should not be interpreted as a measure of those species’ economic importance (or lack thereof).
The taphonomic data, summarized in Table 1 and Supplemental Table 1, suggest good preservation of the bone assemblage. Only 0.5% of specimens were weathered, 0.75% were gnawed, 1.5% were burnt, 3% were butchered, and 9% were newly broken, indicating that degradation by taphonomic agents was rather low. In contrast, among specimens from the historic site of the San Diego County Warner's Trading Post, dated to AD 1849–1851, 75% of the bones were weathered, 21% were burnt, 10% were butchered, and 44% were newly broken (Sasson Reference Sasson, Van Wormer and Walter2011). We also assessed the depositional integrity of the presidio bone assemblage based on density-mediated attrition of the chicken bones, plotting the frequency of 176 chicken limb elements against the corresponding mean BMDa and BMDv of wild turkey (Supplemental Table 2; Dirrigl Reference Dirrigl2001). If the relative frequency of high-density body parts is greater than that of low-density body parts, one could infer that taphonomic agents modified the bone assemblage and that density-mediated attrition is responsible for the loss or destruction of lower density skeletal elements (Lyman Reference Lyman1994:252). We found no significant correlation between the two variants (r = 0.03, p = 0.87 for BMDa; r = 0.21, p = 0.30 for BMDv), suggesting that density-mediated attrition did not play a major role in structuring the relative frequency of chicken skeletal elements. Additional evidence for postdepositional integrity of the assemblage is the large proportion of complete bones. Among all mammalian and avian specimens, 23% (n = 185) were whole (complete) bones. In the study of large-mammal remains from the San Diego Presidio, Cheever (Reference Cheever1983) reported similar results: nearly 30% of the studied sample consisted of whole elements.
Adult/Juvenile Ratio
Data on the epiphyseal fusion of 106 chicken elements indicate that 23% of specimens were from birds culled at ages under four to six months. The assemblage of unfused bone includes six radii, with an MNI of three chickens killed prior to four months of age, and eight tarsometatarsi, with an MNI of five chickens killed before six months of age (Supplemental Table 3). Additionally, 12 specimens lacking fusion data exhibited a porous cortical surface. They also correspond in size to the smaller unfused specimens and appear to be from young birds likely culled well before their fourth month. Although juvenile chicken remains cannot be sexed, we believe that most if not all of the immature specimens belong to males. Very little data exists on the ratios of adult/juvenile chickens from North American sites. Simons's (Reference Simons and Felton1984) analysis of chicken remains from early twentieth-century Chinese-American communities at Ventura, California, and Lovelock, Nevada, point to equal proportions of adult and juvenile chickens. The proportion of juvenile chickens in medieval and post-medieval European sites is more comparable to that of the San Diego Presidio, with over 20% of specimens from immature chickens (Moreno-García and Detry Reference Moreno-García, Detry, Prummel, Zeiler and Brinkhuizen2010; Serjeantson Reference Serjeantson2009:81).
Female/Male Ratio
We examined the chicken assemblage for spurs. Of 21 complete or partial tarsometatarsi, 15 specimens retained the distal end where large bone spurs typically develop in roosters and capons. None showed evidence of spurs, something not uncommon in zooarchaeological assemblages (Moreno-García and Detry Reference Moreno-García, Detry, Prummel, Zeiler and Brinkhuizen2010).
In our assessment of male/female ratio in the domestic fowl assemblage, we used two osteometric datasets: greatest breadth and length (Supplemental Table 4). Mixture analysis of the greatest breadth (n = 90) detected two groups of what appear to be female and male chickens at a ratio of 12:1 (Table 3; Figure 3). Mixture analysis of the greatest length (n = 42) detected three groups: males, females, and most likely a smaller breed of chicken (Table 3; Figure 4). The female/male ratio in this dataset is 5:1. All measurable specimens, including those containing medullary bone, were included in the mixture analysis modeling. None of the specimens identified as males in the mixture analysis contained medullary bone.
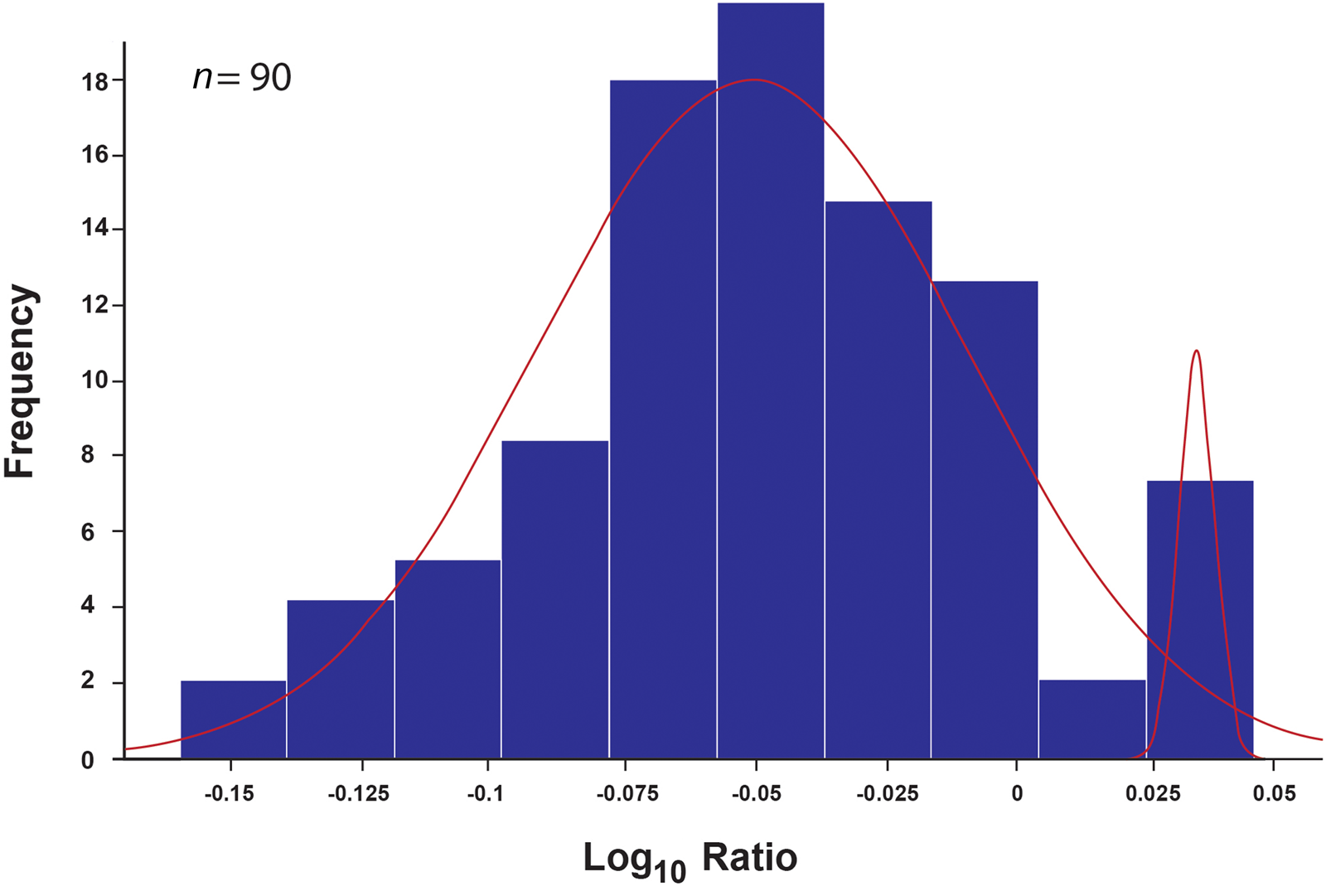
Figure 3. Distribution of the greatest breadth (Bp, Bd, Did, Dip) of long bones with corresponding results of mixture analysis (n = 90). The arches highlight the two groups detected. The larger group of smaller specimens (left) represents females, and the smaller group of larger specimens (right) represents males. The figure was produced with PAST 3.21 software.
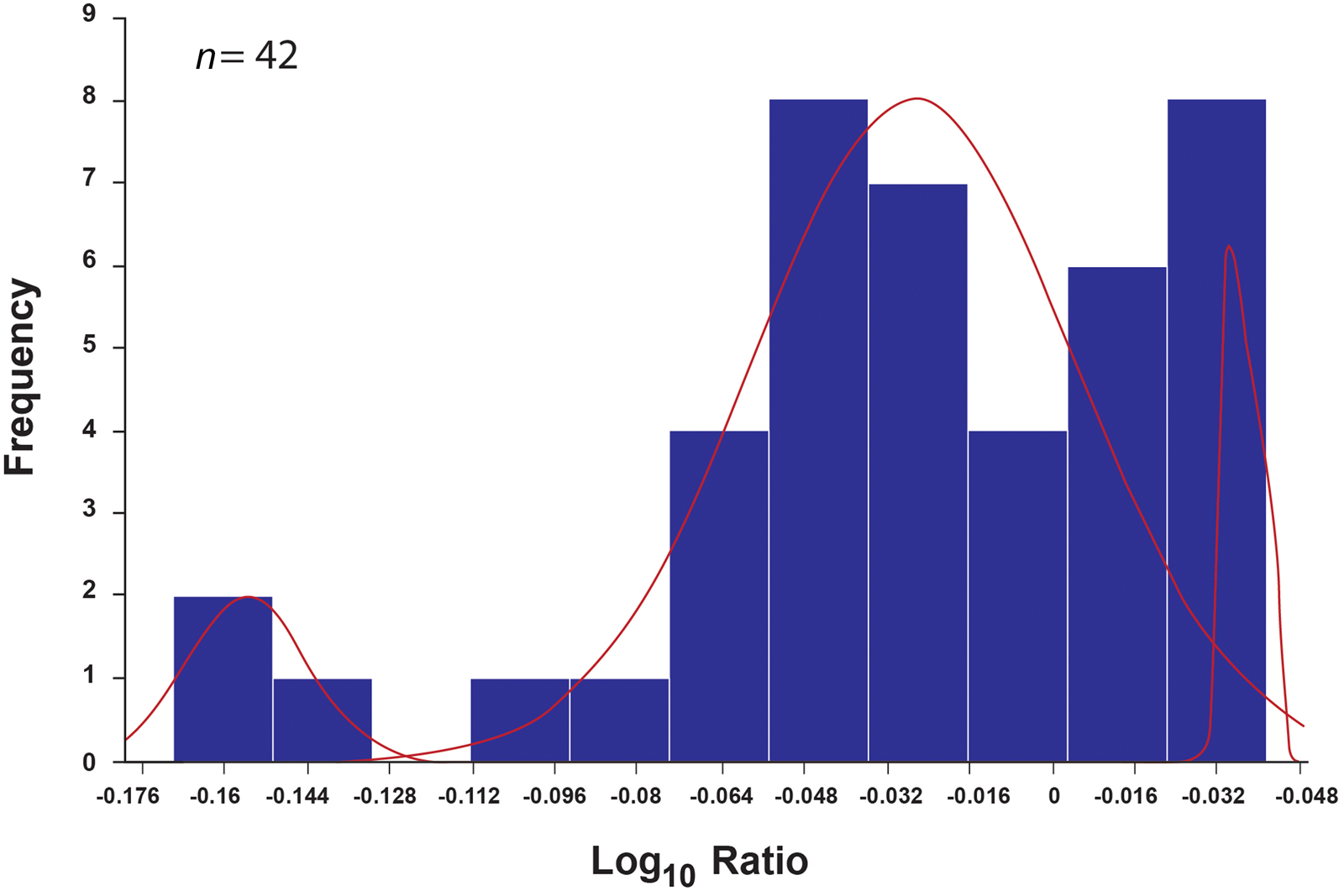
Figure 4. Distribution of the greatest length (GL) of long bones with corresponding results of mixture analysis (n = 42). The arches highlight three groups detected. The smaller group of longer bones (right) represents males, the larger group of shorter specimens (middle) represents females, and the small group of very short specimens (left) likely represents short-legged bantam. The figure was produced with PAST 3.21 software.
Table 3. Mixture Analysis Results of Chicken Long Bones.
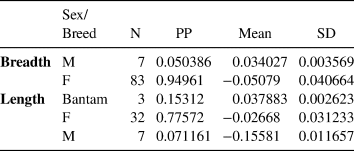
Notes: N = Number of Specimens; PP = Prior Probability; SD = Standard Deviation)
The smaller specimens, which were detected by the mixture analysis of the greatest length but not by that of the greatest breadth, suggest that the shorter specimens belong to a short-legged breed, probably the “creeper” bantam (Figure 5; Dunnington and Siegel Reference Dunnington and Siegel1991; Gordon et al. Reference Gordon, Thomas and Foster2015). This correlates with evidence of smaller chicken breeds reported from European archaeological sites from the Roman period (Benecke Reference Benecke1993; Corona Reference Corona, Prummel, Zeiler and Brinkhuizen2010; De Cupere et al. Reference De Cupere, Van Neer, Monchot, Rijmenants, Udrescu and Waelkens2005) and from Spanish Florida (Reitz et al. Reference Reitz, Scarry and Seifert1985:71).
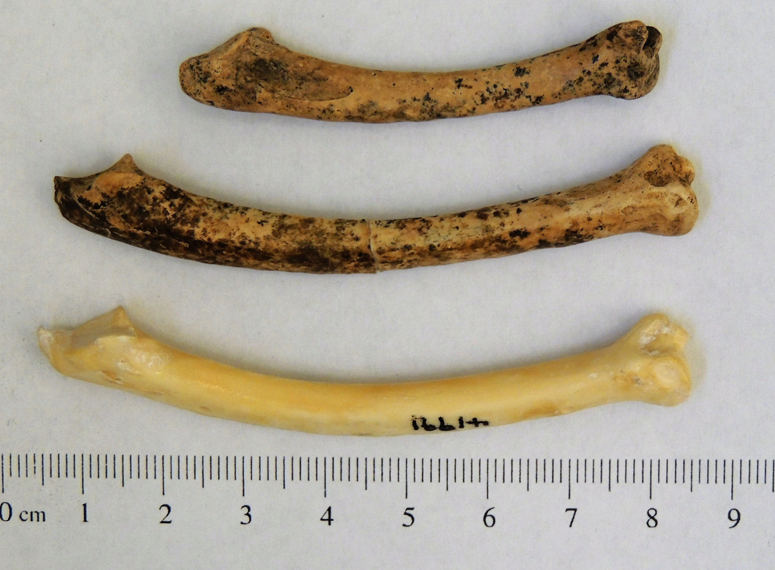
Figure 5. Chicken ulnae: (top) bantam chicken from north wing, Room10; (middle) normal-size chicken from north wing, Room 5; (bottom) modern chicken from the San Diego Natural History Museum collection. (Color online)

Figure 6. Medullary bone (marked by arrow) in chicken tibiotarsus from the Gateway area, unit QR21, level 182–198 cm. (Color online)
All avian remains, including 126 chicken specimens, were inspected for medullary bone. Of note are a barn owl (Tyto alba) tibiotarsus and redhead duck (Aythya americana) coracoid that also contained medullary bone. Among the 126 chicken specimens, 29 unfused or fragmented distal or proximal articular ends insufficient for determining the presence or absence of medullary bone were excluded from the analysis. Of the 97 remaining adult chicken specimens, 53% (n = 51) contained medullary bone (Supplemental Table 5). If we calculate the proportion of females identified from medullary bone identified through mixture analysis (see above), the percentage of egg-laying hens increases to 61% (51 out of 83; Table 3). It should be remembered that medullary bone occurs in chickens for a short time—during the egg-laying period—and it is fundamentally weaker than structural cortical bone (Whitehead Reference Whitehead2004). Therefore, archaeological preservation of medullary bone should be relatively low. The high percentage of medullary bone in the present study provides strong evidence that chickens were utilized for egg production. It also suggests that a significant number of the older hens, whose productivity was decreasing, were culled (Van Neer et al. Reference Van Neer, Noyen, De Cupere and Beuls2002).
Medullary bone in chickens has been reported from historic sites in the southeastern United States (Reitz et al. Reference Reitz, Scarry and Seifert1985:83; Rick Reference Rick1975), but no quantitative data from western North American sites are available for comparison. Most data on zooarchaeological medullary bone come from Europe and the Middle East. In Britain, Serjeantson (Reference Serjeantson2009:283–284) noted an increase in the incidence of medullary bone from Roman times through the Middle Ages, from 20% to 66%. Moreno-García and Detry (Reference Moreno-García, Detry, Prummel, Zeiler and Brinkhuizen2010) reported that 27% of the chicken-bone assemblage from a seventeenth-century monastery in Portugal contained medullary bone. The average proportions of fowl limbs with medullary bone from the Roman sites of Berenike on the Red Sea coast and Sagalassos in western Turkey were close to 66% and 54%, respectively (De Cupere et al. Reference De Cupere, Van Neer, Monchot, Rijmenants, Udrescu and Waelkens2005; Van Neer et al. Reference Van Neer, Noyen, De Cupere and Beuls2002). Biological and zooarchaeological studies indicate that skeletal elements with a rich blood supply also accumulate the greatest quantity of medullary bone (Canoville et al. Reference Canoville, Schweitzer and Zanno2019; Rick Reference Rick1975; Van Neer et al. Reference Van Neer, Noyen, De Cupere and Beuls2002; Whitehead Reference Whitehead2004). Our data from the San Diego Presidio support this notion: three elements in particular—the tibiotarsus, femur, and ulna—contained high proportions of medullary bone at 80%, 75%, and 54%, respectively.
Body-Part Representation and Butchering
Differential disposal of butchering and consumption waste is evident among the San Diego Presidio chicken remains. Head bones and phalanges account for only 9% of chicken elements, suggesting that they were not generally discarded with food waste (Supplemental Table 6; Figure 7). The phalanges include those of both the foot (NISP = 20) and the wing (NISP = 4). Seven foot phalanges (four first phalanges; three second phalanges), from the midden outside the western wall, were most likely articulated upon deposition, supporting the notion that head and toes were discarded prior to the preparation and consumption of the meat-bearing portions of the bird. Nearly even representation of other body-part categories (axial, breast, leg, and wing) points to consumption of whole chickens, rather than merely legs and wings (Supplemental Table 6; Figures 7 and 8). As with other criteria for assessing chicken husbandry, discussed above, very little comparative data exists on chicken body-part representation from North American sites. A study of chicken body-part representation from the early nineteenth-century Ontiveros Adobe in Santa Fe Springs, California, also notes the scarcity of head and foot bones, suggesting that these elements were disposed of separately from kitchen consumption refuse (Gust Reference Gust and Frierman1982).
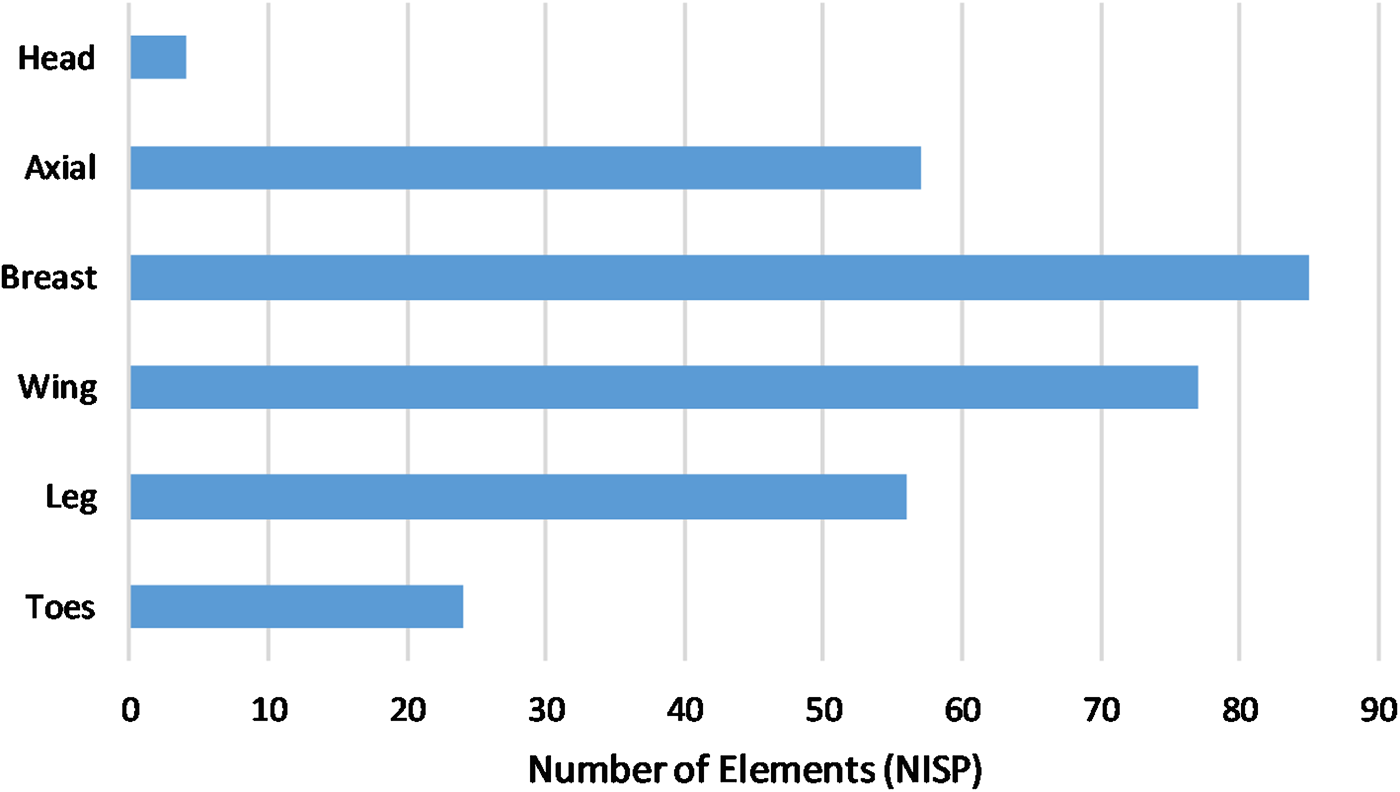
Figure 7. Chicken body-part representation.

Figure 8. Chicken axial skeleton and limb bones from the north wing, Building 2, Room 5. Elements sectioned for medullary bone inspection are marked by circles; cut marks are marked by arrows. (Color online)
All 17 recorded butchery scars (cleave and cut marks) were observed on specimens identified as consumption waste. Close to 70% of the scars were on breast bones associated with disarticulation of the furcula, coracoid, and scapula—three elements bound at the shoulder by tenacious ligaments (Figure 9).
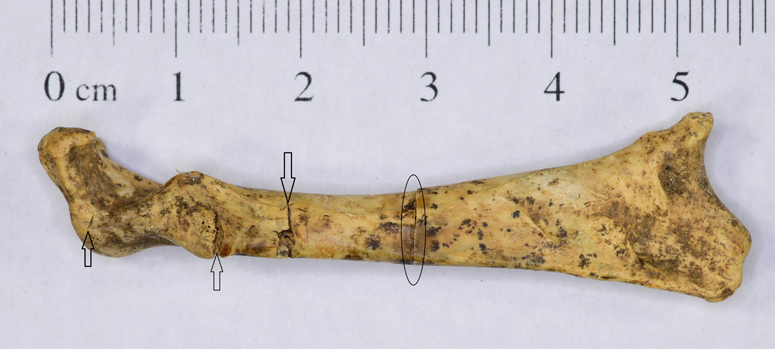
Figure 9. Cut marks (marked by arrows) on chicken coracoid from the north wing, Building 2, Room 5. Sectioning of the coracoid for medullary bone inspection is marked by an oval line. (Color online)
Other Fauna
The remains of wild birds in the bone assemblage from the San Diego Presidio include four species of raptors and eight of waterfowl (Table 1). The range of species reflects exploitation of a variety of local habitats including coastal bays, fresh- and saltwater estuaries, brush and grasslands, river bottoms, and environs ranging from coastal slopes to local mountain ranges (Unitt Reference Unitt2004).
Two species among the wild avifauna are worth comment. The American crow (Corvus brachyrhynchos) was unrecorded around San Diego from the 1870s until the 1980s, making its presence at the presidio in the late eighteenth to early nineteenth century surprising (Arter et al. Reference Arter, Sasson, Unitt, Shuford, Gill and Handel2018; Unitt Reference Unitt2004:381). The sandhill crane (Grus canadensis) no longer exists in San Diego County, but it was reported in the nineteenth century, flying over in significant numbers during spring and fall migrations. The presidio specimen represents the only physical evidence of cranes in this region (Arter et al. Reference Arter, Sasson, Unitt, Shuford, Gill and Handel2018; Unitt Reference Unitt2004:190–191).
Non-avian bone specimens (n = 321) include 16 species of wild and domesticated animals (Supplemental Table 1). Wild species include rabbit, hare, squirrel, and fish. Cattle, sheep, goat, pig, dog, and cat remains are also represented. Since our focus is on avian remains, the non-avian species are not further discussed here.
Discussion and Conclusions
California presidios were not economically self-sufficient. They were provisioned with livestock and a variety of goods from Mexico and, later, from nearby missions. Zooarchaeological analysis of chicken remains from the San Diego Presidio provides key insights into the important role of chickens both in the colonial Spanish subsistence economy and on husbandry practices within the presidios.
In traditional poultry husbandry, females are kept for reproduction and egg production, and males are culled (Blackie Reference Blackie2014; Van Neer et al. Reference Van Neer, Noyen, De Cupere and Beuls2002). Consequently, if mission provisioning included regular transfer of chickens to the presidios, a high proportion of hens would be expected at the provisioning site (missions) and a high proportion of males at the provisioned site (presidios). Zooarchaeological data from the San Diego Presidio indicate the opposite; the representation of females in the death assemblage is significantly higher than males. Moreover, various criteria examined in this study—including the male/female ratio, adult/juvenile ratio, and relative proportion of egg-laying hens at the presidio—reflect on-site, small-scale chicken husbandry focused on meat and egg production. The osteometric data indicate an approximate average ratio of 8.5 hens to 1 rooster, suggesting that most males were culled at a young age, whereas most females were kept alive. The medullary bone data suggest that when hens’ productivity declined, they were culled and eaten. Cut marks on two specimens with medullary bone support this interpretation. The age data indicate that nearly one-quarter of specimens were immature, which suggests that most of these juveniles were males. This could also explain the small percentage of adult males in the death assemblage.
Body-part representation suggests that, with the exception of the head and toes, nearly the entire chicken was consumed—consumption habits similar to those we see today. The fact that only 1.6% of the chicken bones were burnt provides additional evidence that chickens were roasted whole and that their bones were generally not exposed to fire. In addition, based on the medullary bone and the mixture analysis data, 61% of specimens were from egg-laying hens—a clear indication that chickens were raised at the San Diego Presidio for their eggs as well as for meat.
Studies of small-scale poultry production in developing countries show that all families at the village level are owners of poultry (Mack et al. Reference Mack, Hoffmann and Otte2007). Small-scale poultry production is maintained mainly by women with very low inputs (i.e., land, labor, and capital) while providing the population with dependable and readily harvested meat and eggs. This constitutes a vital source of protein for immediate home consumption as well as a valuable source of barter or income (Blackie Reference Blackie2014; Copland and Alders Reference Copland, Alders, Alders, Spradbrow and Young2009; Guèye Reference Guèye2000; Sonaiya Reference Sonaiya, Alders, Spradbrow and Young2009). Poultry husbandry in developing countries is a practical and viable option for poverty alleviation. It allows for a quick return to productivity after environmental disasters, wars or, recently, pandemics, and it can be adapted to many different agroecological zones (Copland and Alders Reference Copland, Alders, Alders, Spradbrow and Young2009; Guèye Reference Guèye2000; Mack et al. Reference Mack, Hoffmann and Otte2007; Sonaiya Reference Sonaiya, Alders, Spradbrow and Young2009).
Chickens obtain most of their diet from their immediate environment by scavenging for earthworms, insects, and household refuse. They play an important role in pest control and provide organic fertilizer (Abubakar et al. Reference Abubakar, Ambali and Tamjdo2007; Copland and Alders Reference Copland, Alders, Alders, Spradbrow and Young2009; Guèye Reference Guèye2000; Redding Reference Redding2015; Webster Reference Webster1899). Moreover, according to Redding (Reference Redding2015), in subsistence-based systems, chickens have a major advantage over almost every other domesticated animal in converting the food they ingest into protein. For example, production of chicken meat is two to three times more efficient than that of pig meat, and it is four to six times more efficient than meat production from cattle and sheep. Chickens also require less water (3,500 L) to produce 1 kg of meat than do pigs (6,000 L), cattle (43,000 L), or sheep (51,000 L). In addition, the chicken is a smaller “meat package” than most other domesticates, and a household can consume a chicken within 24 hours.
We believe that the extraordinary attributes of poultry production in developing countries are applicable to the small-scale chicken husbandry practiced by the inhabitants of the California presidios. Since the presidios were reliant on food supplied from Mexico and missions nearby, it is very likely that the Spanish government was favorable toward the presidios raising chickens. Small-scale chicken husbandry could help alleviate food insecurity experienced by the presidio families while ultimately providing a form of economic independence. It is reasonable to assume that the presidio chickens were managed by women and children as a low-investment garden crop, providing high-value protein and a possible source of barter or income. We argue that poultry husbandry at the presidios also played an important role in household disaster-mitigation strategies (e.g., drought).
Interestingly enough, it seems that chickens were rather “invisible” when it came to enumerating livestock animals in the historical documentation of early California (Archibald Reference Archibald1978:178; for similar observation from Spanish Florida, see Reitz et al. [Reference Reitz, Scarry and Seifert1985:71]). For example, a zooarchaeological study from deposits at the Mission Basilica San Diego de Alcalá, which was established contemporaneously with the San Diego Presidio, shows that chicken constituted 7% of the mission's livestock remains (Sasson Reference Sasson2014). However, the inventories of livestock at the San Diego Mission in 1773 (Palóu Reference Palóu1926:216–217), 1777 (Engelhardt Reference Engelhardt1920:91–92), and 1783 (Kenneally Reference Kenneally1965:93) enumerated cattle, sheep, goats, hogs, horses, and mules but neglected to mention chickens. Moreover, the “invisibility” of chicken is rather curious since our survey of historical documents describing early California suggests that chickens and eggs were common commodities in the presidios and missions (Bancroft Reference Bancroft1888:628; Engelhardt Reference Engelhardt1912:602; Pourade Reference Pourade1961:217; Richman Reference Richman1911:350; Webb Reference Webb1952:189). Historical accounts note that ships anchored at San Diego, and travelers passing by the presidio or mission, were provisioned with chickens (Archibald Reference Archibald1978:106; Bancroft Reference Bancroft1885:12, Reference Bancroft1888:274; Pourade Reference Pourade1961:96; Serra Reference Serra and Tibesar1955:1345). Redding (Reference Redding2015) suggests, with regard to pigs in the ancient Near East, that what the textual record reflects is what the central authority and bureaucrats valued as important. Hence, garden crops, such as chicken and pigs, managed by women, were generally not of interest to the central administration and were not central to the system-wide redistributive economy. This bureaucratic oversight does not, however, preclude chickens from playing an important role in the presidio subsistence system.
Historical accounts about price regulations in Spanish California yield insight into the value of a chicken in AD 1781. Commodities supplied to soldiers in California presidios were charged against their wages, and the Spanish government had an interest in keeping food prices low. A price-fixing list indicates that the cost of a single hen (2.5 reales) was equivalent to the price of a suckling kid goat, five rabbits, or five pounds of cheese. A single hen was valued higher than a dozen quail or a pair of pigeons (Mosk Reference Mosk1938; see also Hackel Reference Hackel1997; Hittell Reference Hittell1897:534; Sánchez Reference Sánchez1990). The fact that the price of hens was regulated may suggest that presidio families supplemented their income selling chickens.
Another historical account provides a rare opportunity to compare the results of our zooarchaeological study at the San Diego Presidio against statistical data in the historical record. Fermín de Francisco Lasuén arrived in Baja California in 1768 and ultimately founded eight missions in Alta California. In 1773, during his visit to Mission San Francisco Borja in Baja California Sur, Father Lasuén generated an inventory of the mission and noted, “Also, chicken run with two roosters and eighteen hens” (Kenneally Reference Kenneally1965:28). This rare account from San Borja indicates husbandry focused on hens with a 1:9 rooster/hen ratio, which supports our conclusion that small-scale chicken husbandry was practiced at the presidios. The similarity between the data from San Borja and our osteometric analysis is striking. Our mixture analysis yielded an average rooster/hen ratio of 1:8.5 (1:12 based on the greatest breadth, and 1:5 based on the greatest length)—nearly equal to the ratio from the historical account.
Historical records also indicate that during the early years at the San Diego Presidio, soldiers were supplied powder and shot for hunting local wild birds to supplement their meager dietary rations (Mason Reference Mason1978). This may explain the extraordinary number of wild avian species identified in our bone assemblage, including eight taxa of waterfowl (Table 1).
It is quite evident that once the Spanish colonists introduced domestic fowl to Alta California, chickens became an integral part of the subsistence economy (Archibald Reference Archibald1978:160; Bancroft Reference Bancroft1888:364). Mass production of chicken meat and eggs was feasible only with the introduction of commercial artificial incubation in the late 1800s (Christenson Reference Christenson and Hemphill1993; Lawler Reference Lawler2014:204; Simons Reference Simons, Praetzellis, Praetzellis and Brown1980, Reference Simons and Felton1984). Commercial poultry production required large capital inputs, technological investment, and much broader markets (Copland and Alders Reference Copland, Alders, Alders, Spradbrow and Young2009). Backyard poultry husbandry on the other hand, proved to be an easy, viable, and reliable source of meat, eggs, and supplemental income for the smallholder, and it is not surprising that it continued to be practiced concurrent with commercial poultry production (Sasson Reference Sasson, Van Wormer, Walter and Wade2017; Schulz et al. Reference Schulz, Quinn and Fulmer1987; Schweitzer Reference Schweitzer2010; Simons Reference Simons and Felton1984; Szuter Reference Szuter and Orser1996).
This article represents the first comprehensive study of chicken remains from a Spanish-era site in colonial North America. As discussed above, we posit that chickens were consequential to the food security of the early California settlers. Further zooarchaeological studies, however, are much needed to better document, interpret, and understand the nature and extent of the role chickens played in Spanish colonial economies.
Acknowledgments
We wish to thank the City of San Diego for allowing us access to the faunal remains from the San Diego Royal Presidio. The City of San Diego and the Rod Heller Memorial Scholarship Fund, together with Paul Chace, generously helped fund our study. We are grateful to Melinda Zeder, Naomi Sykes, Philip Unitt, and Dayle Cheever for their helpful comments on earlier drafts of this article. This article greatly benefited from insightful comments and suggestions from four anonymous reviewers and Lynn Gamble, the journal editor. San Diego Natural History Museum staff helped with the reproduction of several figures in this article. Zaira Marques translated the abstract and keywords.
Data Availability Statement
The faunal remains discussed here are presently housed at the Zooarchaeology Laboratory, San Diego Natural History Museum. They are on loan from the City of San Diego.
Supplementary Materials
For supplementary material accompanying this article, visit https://doi.org/10.1017/aaq.2020.27.
Supplemental Table 1. Non-Avian NISP, Number of Bones per Size-Group, and Modification Marks.
Supplemental Table 2. Chicken Skeletal Elements NISP and Their Bone-Density Values (Density data from Dirrigl Reference Dirrigl2001).
Supplemental Table 3. Chicken Age at Death.
Supplemental Table 4. Measurements of Chicken Long Bones.
Supplemental Table 5. Chicken Medullary Bone Data (Medullary bone detected = M+; No medullary bone = M-; Questionable medullary bone = M?).
Supplemental Table 6. Relative Frequency of Skeletal Elements and Number of Elements with Butchery Marks.













