



Introduction
The Amazon, the largest remaining tract of tropical forest on earth, is drained by a spectacular river system, which includes 20 of the 30 world’s largest rivers (Latrubesse et al. Reference Latrubesse, Stevaux and Sinha2005). The high levels of annual precipitation and the seasonal nature of rainfall, which is unevenly distributed throughout the year (dry and rainy seasons), result in large oscillations in the levels of streams and rivers, which flood extensive areas every year (Junk et al. Reference Junk, Piedade, Schöngart, Cohn-Haft, Adeney and Wittmann2011). These areas subject to flooding are known as the Amazonian floodplains, which may remain underwater from a few days to several months every year. The floodplains are covered by a complex mosaic of habitats ranging from barren beaches and sand-bar scrubs to seasonally flooded and transitional forests depending, among other things, on the amount of time they remain flooded each year (Junk et al. Reference Junk, Piedade, Schöngart, Cohn-Haft, Adeney and Wittmann2011).
The avifauna of the floodplains (known in Brazil as várzeas and igapós) is among the most poorly known of the entire basin. Although widespread and common waterbirds such as herons, cormorants, gulls, ducks, and kingfishers are ubiquitous components of Amazonian freshwater systems (Cintra Reference Cintra2015), many endemic and specialised species are restricted to floodplain forests and other riverine habitats (Rosenberg Reference Rosenberg1990). Published estimates suggest that riverine habitats contribute with up to 15% of the overall terrestrial avian diversity of Amazonia and include a number of threatened and endemic bird species (Remsen and Parker Reference Remsen and Parker1983). Despite their obvious importance for understanding Amazonian diversity patterns, distributions of flooded forest birds remain poorly described (Cohn-Haft et al. Reference Cohn-Haft, Naka, Fernandes and Albernaz2007).
Historically, most Amazonian riverine habitats have suffered relatively minor impacts from the ribeirinhos (local Amazonian human populations), whose livelihoods depend on artisanal fishing and subsistence agriculture. This apparent equilibrium between human populations and riverine biodiversity has changed dramatically in the last decades, with the decisions by Amazonian countries to unleash the hydroelectric potential of their rivers. Almost two hundred dams have already been built in the Amazon basin (RAISG 2013, Lees et al. Reference Lees, Peres, Fearnside, Schneider, Jansen and Zuanon2016), and plans include the construction of another 200 in the next decades, including dozens of large dams (> 30 MW), which are particularly damaging to the environment (Fearnside Reference Fearnside2015a). The last decade has been especially active for hydropower development in the Brazilian Amazon (Finer and Jenkins Reference Finer and Jenkins2012). Several major Amazonian rivers, such as the Madeira, Araguaia, Tocantins, and Xingú have already been dammed with either negative or catastrophic consequences on their biodiversity (Benchimol and Peres Reference Benchimol and Peres2015, Lees et al. Reference Lees, Peres, Fearnside, Schneider, Jansen and Zuanon2016, Assahira et al. Reference Assahira, Piedade, Trumbore, Wittmann, Cintra, Batista, de Resende and Schöngart2017). Despite the unprecedented rate of dam construction throughout the tropics (Laurance et al. Reference Laurance, Peletier-Jellema, Geenen, Koster, Verweij, Van Dijck, Lovejoy, Schleicher and Van Kuijk2015), most paradigms of how hydrological modifications alter regional ecology are based on temperate zones (Pringle et al. Reference Pringle, Freeman and Freeman2000). Generalisations about the impacts of dams in tropical rivers are constrained by limited data and by a lack of baseline information on the distribution and ecology of the biota before dams are actually built (Pringle et al. Reference Pringle, Freeman and Freeman2000).
Many of these Amazonian dams have been constructed despite opposition from scientists, non-governmental organizations (NGOs), local populations, and indigenous communities (Fearnside Reference Fearnside2015b, Latrubesse et al. Reference Latrubesse, Arima, Dunne, Park, Baker, d’Horta, Wight, Wittmann, Zuanon, Baker and Ribas2017). Recent joint efforts by these actors, however, have managed to halt the construction of the São Luiz do Tapajós dam, one of the most important and strategic Amazonian hydropower development projects according to the Brazilian government (Naka Reference Naka2015). Whether the Brazilian Government’s decision not to pursue this dam (for now) was due to rising costs during an unprecedented economic crisis, or due to scientific data predicting enormous impacts on biodiversity, it remains clear that good science and high-quality biodiversity data are among the best strategies in hand to raise conservation concerns facing infrastructure projects.
One especially interesting Amazonian river that has not yet been dammed is the Rio Branco, located within the northern Brazilian state of Roraima. Although it is a relatively small river by Amazonian standards, it represents the largest tributary of the Rio Negro and a river of remarkable features. Located in one of the most sparsely populated regions of Amazonia, the Rio Branco was, until recently, one of the least known rivers in the basin. In recent years, however, biologists started to pay attention to this often-neglected river, mostly due to some unique biogeographical and ecological features (Ferreira et al. Reference Ferreira, Zuanon, Forsberg, Goulding and Briglia-Ferreira2007, Naka et al. Reference Naka, Cohn-Haft, Whittaker, Barnett and Torres2007, Laranjeiras et al. Reference Laranjeiras, Naka, Bechtoldt, Costa, Andretti, Cerqueira, Torres, Rodrigues, Santos, Pacheco, Sardelli, Barnett and Cohn-Haft2014).
Biogeographically, the Rio Branco has been considered an important barrier for forest-dwelling animals, particularly birds (Naka et al. Reference Naka, Cohn-Haft, Mallet-Rodrigues, Santos and Torres2006, Naka Reference Naka2011) and primates (Boubli et al. Reference Boubli, Ribas, Alfaro, Alfaro, da Silva, Pinho and Farias2015). The distributions of at least 40 pairs of terra firme forest avian taxa are bounded by this river (Naka et al. Reference Naka, Bechtoldt, Henriques and Brumfield2012, Naka and Brumfield Reference Naka and Brumfield2018), and as such it can be considered the natural boundary of the Guianan Area of Avian Endemism (Naka Reference Naka2011). In fact, no other river of its size is known to delimit the distribution of so many avian taxa. Ecologically, two features can be highlighted: the Rio Branco represents an isolated white-water (or muddy) river in a black-water basin, and the river crosses one of Amazonia’s largest rainfall gradients (1,100–2,600 mm/yr) and a major geomorphological and climatic boundary along a savanna/forest ecotone. The upper Rio Branco is located within the savanna domain, on top of the Boa Vista sedimentary formation and the heavily weathered crystalline rocks of the Guiana Shield, in areas where it rains less than 1,700 mm/yr (Barbosa Reference Barbosa, Barbosa, Ferreira and Castellón1997, Araujo et al. Reference Araujo, Andrade Junior, Medeiros and Sampaio2001). In this region, river margins are composed of narrow gallery forests. Further south, the lower Rio Branco runs through a well-established sedimentary basin, in areas where it rains considerably more (up to 2,600 mm/yr) (Barbosa Reference Barbosa, Barbosa, Ferreira and Castellón1997). There, the river floodplains are covered by wide stretches of more typical Amazonian várzea forests.
The Bem Querer rapids, which mark the boundary between the Guiana Shield and the sedimentary basin, are in the sights of the Brazilian Government, which has already approved construction of at least one large dam on the Rio Branco. Four additional dams, on the Mucajaí and Cotingo rivers, have been among the Government’s plans for decades (Fearnside and Barbosa Reference Fearnside and Barbosa1996). The major hydroelectric dam planned for the Branco has been named after the Bem Querer rapids and is likely to dramatically change the river forever, flooding large areas above the rapids, and disrupting the natural flow of the river downstream. Government plans include an associated hydroway, a complex array of dams that would allow river navigability throughout the year, extending the effects of the dam to the entire river. Furthermore, these threats only represent direct impacts of infrastructure projects. Indirect impacts, such as population expansion due to the influx of dam workers, the construction of new roads during dam-building phase, and even the establishment of new mining companies in the area have been shown to have catalyse further habitat loss and degradation than the dam themselves (Ferreira et al. Reference Ferreira, Aragão, Barlow, Barreto, Berenguer, Bustamante, Gardner, Lees, Lima, Louzada and Pardini2014).
In this paper, we present the results of our avian surveys conducted on the Rio Branco and its major tributaries since 2001. Our goal is to provide high quality avian baseline information that will serve as a basis for future environmental studies. The data presented are the results of multi-year, multi-researcher explorations and include opportunistic observations as well as standardised censuses systematically conducted along the entire length of the river. We present a fully documented list of the avifauna (including available specimens, recordings, and photographs), and describe habitat use and general avian distribution patterns along the river. We show that many of the species detected on the Rio Branco are not widely distributed along the river and that a significant number of species are specialists restricted to single habitats. We demonstrate that the Rio Branco presents two distinct avifaunas along its course, one mostly restricted to the gallery forests on the upper section of the river, and the other, mostly restricted to the varzea forests downriver of the Bem Querer rapids. We also highlight the presence of 25 bird species of conservation concern (endemic and threatened) that occur along the riverine habitats of the Rio Branco. Finally, we use a species indicator analysis and our own experience of the river to provide a list of 50 species that we believe should be monitored in future environmental studies. Some of these species are threatened, while others are habitat specialists, all being common enough to represent good target species. We believe these species may be particularly threatened by the construction of a hydroelectric dam and may suffer changes in abundance as well as drastic range reduction. Our results suggest that a number of species may be directly impacted by this particular dam and its associated hydroway, with unpredictable consequences to this evolutionary and ecological hotspot, and the overall species diversity of the river.
Methods
Study area
The Rio Branco basin (∼ 192,000 km2) is located entirely within the Brazilian state of Roraima. The name Rio Branco is applied to that stretch of the main river that flows directly south from the confluence of the Tacutu and Uraricoera rivers in the Roraima-Rupunnuni savannas to the Rio Negro (Figure 1). Although the Branco is the main tributary of the Rio Negro, it is only ∼ 550 km long and it represents a medium-sized river by Amazonian standards, ranking 12th in discharge volume (Ferreira et al. Reference Ferreira, Zuanon, Forsberg, Goulding and Briglia-Ferreira2007) and 18th in length (Goulding et al. Reference Goulding, Barthem and Ferreira2003). As its name suggests, the Rio Branco (“white river” in Portuguese) is one of the few muddy, sediment-rich rivers in the Guiana Shield, otherwise known for its many black-water, nutrient-poor rivers (Goulding et al. Reference Goulding, Barthem and Ferreira2003). It has also been classified as a clear-water river (Junk et al. Reference Junk, Wittmann, Schöngart, Maria and Piedade2015), especially during the dry season (Cremon et al. Reference Cremon, de Fátima Rossetti, de Oliveira Sawakuchi and Cohen2016). However, considering that sediment deposition on the floodplains is determined during the high-water (rainy) season, the Branco behaves ecologically as a white-water river. This is evidenced by the presence of numerous white-water bird (Naka et al. Reference Naka, Cohn-Haft, Whittaker, Barnett and Torres2007, Laranjeiras et al. Reference Laranjeiras, Naka and Cohn-Haft2019) and plant species, such as the kapok Ceiba pentandra, munguba Pseudobombax munguba, and tachi-da-várzea Triplaris surinamensis.
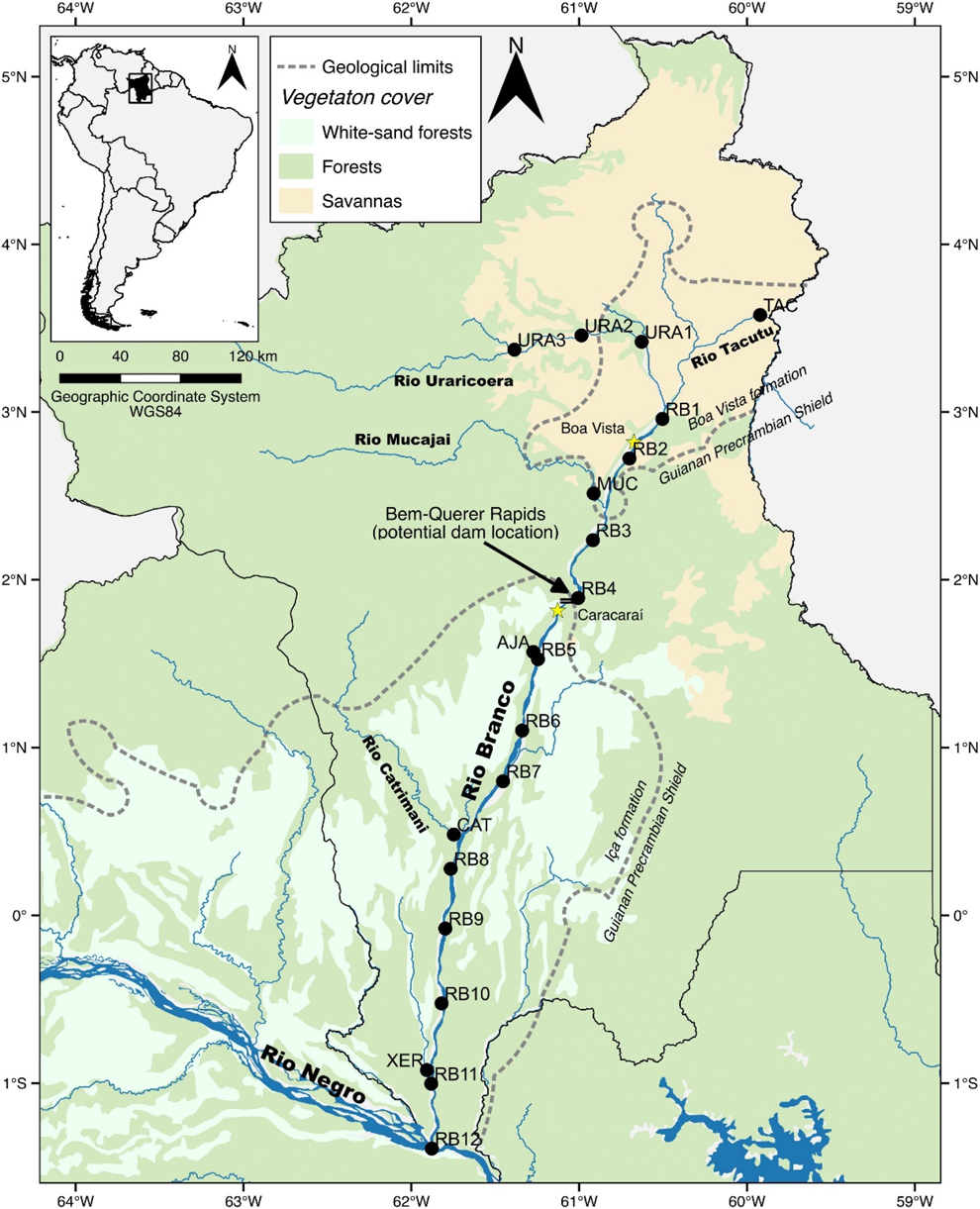
Figure 1. Map of the Rio Branco and its major tributaries surveyed on this study. The black outline indicates the boundaries of the Brazilian state of Roraima. Dotted line indicates major geological limits. Black dots refer to localities where the avifauna was systematically surveyed using point counts, including 12 localities on the Rio Branco, three on the Rio Uraricoera, and single localities on the Tacutu, Mucajai, Ajarani, Catrimani, and Xeruini rivers. The arrow points the projected localization of the Bem Querer Dam, just above the city of Caracaraí.
The Rio Branco can be divided into two main geographical and ecological regions, upper and lower, with the Bem Querer rapids (Figure S1A in the online supplementary material), close to the city of Caracaraí (1°50’N, 61°06’W), as a natural dividing point. The rapids also represent the boundary between the northern crystalline geological formation of the Guiana Shield and the sedimentary formation of the Rio Branco basin (Empresa de Pesquisa Energética 2011). North of Caracaraí, rainfall decreases sharply, with areas where it rains as little as 1,100 mm/yr, whereas on the lower Rio Branco rainfall may exceed 2,600 mm/yr (Barbosa Reference Barbosa, Barbosa, Ferreira and Castellón1997).
Habitats
We follow Remsen and Parker’s (Reference Remsen and Parker1983) description of riverine habitats with some minor modifications, appropriate for our study region. The habitats used by birds mentioned in this study include: i) water (lakes and rivers) (Figure S1B), ii) beaches and sandbars (Figure S1B), iii) sandbar scrub (Figure S1C), iv) river edge forest (or Cecropia-dominated forest) (Figure S1D), v) várzea forest (Figure S2A), vi) transitional forest (Figure S2B), vii) black-water flooded forest (Figure S2C), and viii) gallery forest (Figure S2D). A more complete description and images of these habitats are available in the online Supplementary Material.
The terms várzea and igapó are often used interchangeably by local inhabitants to describe seasonally flooded forests. We follow Prance (Reference Prance1979) who used these terms to distinguish forests flooded by rivers of muddy waters (várzea) or flooded by black-water rivers (igapó). The margins of the lower Rio Branco are occupied by extensive areas of várzeas, whereas some of its tributaries (i.e. Agua Boa do Univini) are black-water rivers whose margins are covered by igapó.
We classified the preferred habitat (or habitats) for each species based on our systematic surveys, bird specimens, audio recordings, and opportunistic observations. Given the high habitat heterogeneity present along the Rio Branco and its tributaries, defining habitats was not always straightforward. In those cases, we opted to include the number of habitats used by each species, or at least where it has been recorded during our sampling. The order of habitats presented in Appendix S1 represents our interpretation of species’ preferences.
Fieldwork and data collection
We first visited the Rio Branco in 2001, when LNN and MFT explored the island of São José, on the upper portion of the river, just above the city of Boa Vista. Since then, we conducted several expeditions in different parts of the river, and on different tributaries, totalling an estimated number of 143 days in the field (Table S1). Among the tributaries where we conducted fieldwork are the Uraricoera, Tacutú, Anauá, Mucajaí, Ajaraní, Água Boa do Univiní, and Catrimani rivers (Figure 1). Until 2012, our sampling was mostly opportunistic, based on day lists, selective collecting and general audio recordings. Our visits were mostly made using riverboats for long-distance travel and speedboats to reach the shores and smaller tributaries. Our observations often started before dawn, and continued until late morning, and continued from the early afternoon until sunset. We documented our observations by means of audio recordings, photographs, and specimen collections (Appendices S1 and S2). Audio recordings are available through xeno-canto (Xeno-canto.org), the Macaulay library of Natural Sounds (Macaulaylibrary.org) or our own personal collections.
In 2012, we began systematic and standardized avian censuses (point-counts) along the entire length of the river. To do so, we established sampling localities every 50 km along the Rio Branco and on some of its tributaries (Figure 1, Table S2). At each locality along the Rio Branco, we made 30 point-counts, including 10 on each side of the river and 10 located on river islands in the middle. Point counts were systematically established every 500 m, covering ∼5 km at each locality. In 2013, we sampled three Rio Branco tributaries: the Ajaraní, Catrimani, and Água Boa do Univiní rivers, which were sampled using a similar scheme (Table S1). Because no islands are present on smaller rivers, we only conducted 20 point counts on those tributaries (10 on each bank). In 2013, we resampled some of the points at four localities (RB5 to RB8, see Figure 1). In 2014 and 2015, we sampled two and one localities separated by 50 km along the Uraricoeara river (Figure 1) with a similar sampling protocol (30 point counts per locality, including 10 on river islands). All point counts lasted 15 minutes and were conducted from the shore, often penetrating a few meters into the forest. Sometimes censuses were conducted directly from the boat, particularly during the dry season when land was hard to reach. Point counts started around 05h30–05h45, before sunrise, and usually finished before 10h00. All counts were conducted by at least one of the most-experienced observers (LNN, TOL, MCH) joined by the other authors.
Avian distribution patterns
Preliminarily, we had observed that many bird species do not occur throughout the entire length of the river. In order to describe general distribution patterns within the Rio Branco basin, we divided the river into upper and lower, which differ in habitat structure and geomorphology (see Study area). Localities north of the Bem Querer rapids (Rio Branco 1-3, Uraricoera 1-3, Tacutu, and Mucajaí) were considered as part of the upper Rio Branco, and localities to the south of the rapids (Rio Branco 4-12, Ajarani, Catrimani, and Xeruini), were considered as part of the lower Rio Branco.
Based on our observations (including point-counts and general surveys) we classified all bird species as: i) widespread (species that were encountered indiscriminately on both the upper and lower Rio Branco); ii) restricted to the upper Rio Branco (species commonly found on the upper portion of the river and its savanna tributaries, and generally absent on the lower portion of it); iii) restricted to the lower Rio Branco (species commonly found on the lower portion of the Rio Branco, and generally absent on the upper portion of the river); and iv) inadequate data, species for which we do not yet have enough data to fully understand their distribution patterns on the Rio Branco basin.
Data analysis
To evaluate differences in bird species composition between the upper and lower Rio Branco, we visualized between-site composition variation by reducing data dimensionality through non-metric multi-dimensional scaling (two axes NMDS ordination), using the Bray-Curtis dissimilarity index (for abundance data). We used the systematic survey data (16 localities), including eight localities from the upper and eight from the lower Rio Branco. To assess statistical significance of observed differences in assemblage composition, we ran a Permutational Analysis of Variance (PERMANOVA) (Anderson Reference Anderson2001, McArdle and Anderson Reference McArdle and Anderson2001). This analysis uses pseudo-F values to compare within and among-group similarity, assessing significance by permutation.
Following our observations that part of the avifauna was restricted to either the lower or the upper portion of the river, we conducted an Indicator Species Analysis (ISA) (Dufrêne and Legendre Reference Dufrêne and Legendre1997) to objectively identify those species that presented higher fidelity to one region than the other. This analysis generates values between 0 and 1, where 1 indicates complete fidelity to one of the groups and presence in every site in that given group. Statistical significance is obtained by conducting 9999 permutations. We ran the ISA using the multipart function of the indicspecies package in R (De Cáceres and Legendre Reference De Cáceres and Legendre2009). We plotted a histogram of frequencies with the species indicated by the ISA as having significant associations with either the upper or lower Rio Branco using the poncho function in R (Dambros Reference Dambros2014). This function arranges species according to environmental gradients, showing species turnover across the landscape. For the ISA, we included only those species recorded during our standardised censuses (point-counts) along the river. In total, we used data from 15 localities ( Table S2), including 332 bird species. The ISA was conducted using two groups, “upper” and “lower” Rio Branco, divided by the Bem Querer rapids ( Figure S1A). Within the upper Rio Branco we included six localities (RB1-3, Ura 1 and 2, and Tac 1) and nine within the Lower Rio Branco (RB 4 to RB12) (Figure 1, Table S2).
Target species for monitoring
A key aspect of environmental assessment studies is to provide specific data for threatened or indicator species, those that are most likely to be affected by the development project under consideration, and those that can be used as proxies for a larger group of species. We selected a number of bird species as appropriate candidates for monitoring, considering their i) threatened status, ii) significant associations with particular regions of the river, iii) habitat specialisation, and iv) frequency of records. Thus, some threatened species were excluded from this list due to their rarity and consequent unlikelihood of providing reliable results in standardized censuses or target searches.
Results
Avian surveys
We catalogued a total of 439 bird species, in 64 different avian families, in the riverine habitats along the entire length of the Rio Branco and its major tributaries (Appendix S1). Most of these species were found along the Rio Branco itself, but a handful of them were found exclusively on a tributary, such as the Uraricoera (Gray-headed Kite Leptodon cayanensis, Tiny Hawk Accipiter superciliosus, Painted Parakeet Pyrrhura picta, White-bearded Manakin Manacus manacus, Pied Water-Tyrant Fluvicola pica, Slender-footed Tyrannulet Zimmerius gracilipes), Anauá (Guianan Streaked-Antwren Myrmotherula surinamensis), Xeruini (Band-tailed Nighthawk Nyctiprogne leucopyga latifascia, Cherrie’s Antwren Myrmotherula cherriei, and Dusky-chested Flycatcher Myiozetetes luteiventris), or the Catrimani river (Bronzy Jacamar Galbula leucogastra, Ivory-billed Aracari Pteroglossus flavirostris). It is likely that some of these species may be found on the Branco itself in future surveys, although half of those species are black-water river specialists, that occur exclusively in white-sand forests, a habitat found along some minor tributaries, but not on the Branco.
A high proportion (87%; 385 spp.) of the bird species listed here were documented by physical evidence, including voucher specimens (258 spp.), audio recordings (190 spp.), and/or photographs (303 spp.) (Appendix S1). The remaining species were listed based on sight and/or auditory records, and most had been documented previously in the state of Roraima and their presence in the study area is not controversial. We have included all modern voucher specimens available from the Rio Branco and the Uraricoera, totalling 1,061 specimens held at three Brazilian institutions, including the Bird Collection of the Universidade Federal de Pernambuco -UFPE (701 specimens), the Museu Paraense Emílio Goeldi – MPEG (187 specimens), and the Instituto Nacional de Pesquisas da Amazonia – INPA (172 specimens) (Appendix S2).
At least 75% of the species listed were recorded in three or more of the 20 localities that we surveyed in detail along the Branco, Uraricoera, and other minor tributaries. Nonetheless, 89 species were found only at single localities (Appendix S1). A third of those (28 spp.) are typical terra firme forest birds that were detected in transitional forests along the lower Rio Branco.
Most of the species are likely residents in the area, although local movements within the flooded forests have not been studied in detail along the Rio Branco or any other Amazonian river. We detected the presence of 21 Nearctic migrants (Appendix S1), but only three likely southern migrants (Large Elaenia Elaenia spectabilis, White-throated Kingbird Tyrannus albogularis and Bran-coloured Flycatcher Myiophobus fasciatus). The relatively low number of southern visitors may be related to a temporal bias in our sampling (mostly concentrated in the low-water period between September and October). It is possible that some populations of Fork-tailed Flycatcher Tyrannus savanna, Tropical Kingbird Tyrannus melancholicus, or Brown-chested Martin Phaeoprogne tapera may represent visitors from southern South America, or even have both resident and migratory populations.
Avian distribution patterns
Using our entire distributional dataset (439 species), we found that one-third of all bird species recorded along the Rio Branco and its main tributaries (32%, 144 spp.) are widely distributed along the entire length of the river. On the other hand, nearly half of the species seem to be restricted to particular regions along the river. About 10% (45 spp.) seem to be restricted to the upper Rio Branco, the Uraricoera and Tacutú rivers, and associated savannas; whereas ∼ 35% are associated with the lower portion of the Rio Branco, living either on flooded forests (25%, 115 spp.) or in patches of transitional/terra firme forests (10%, 44 spp.). A handful of species (7) are restricted to igapós or forests flooded by black-water rivers. The distribution of ∼ 20% of the avifauna (91 spp.) remains poorly understood, and these are listed as having inadequate distribution data (Appendix S1).
The two-dimensional NMDS ordination (stress = 0.08) revealed a geographic separation in bird species composition between sites located on the upper Rio Branco (and its two northern tributaries the Uraricoera and Tacutu rivers) and those located on the lower Rio Branco (Fig. 2). Differences were confirmed by permutation tests (Permanova, pseudo F = 5.17, P = 0.0009, r 2 = 0.27).
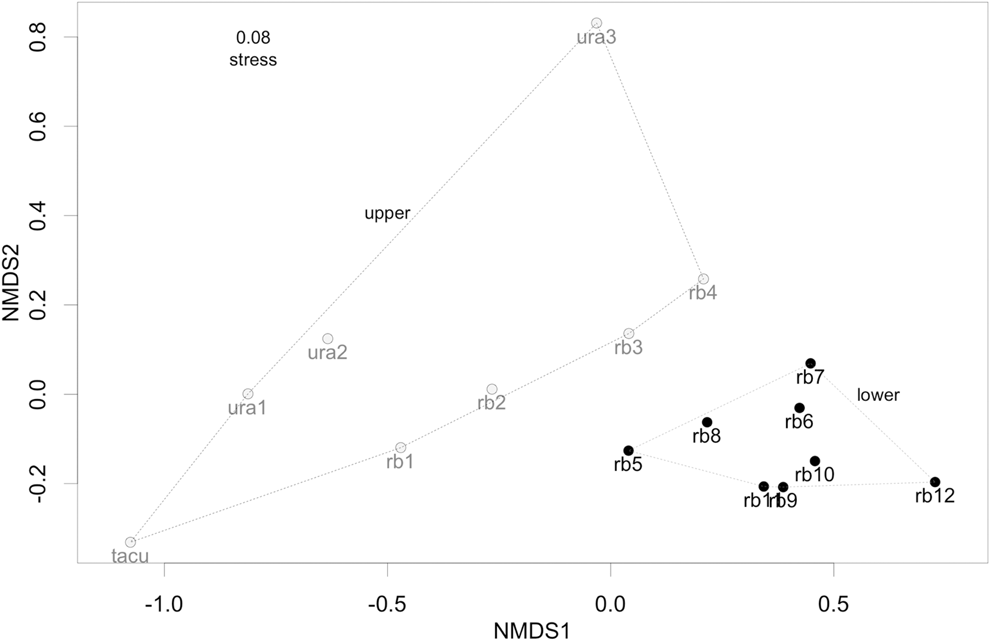
Figure 2. Non-metric multidimensional scaling (NMDS) ordination of 16 localities located along the Rio Branco (rb 1-12), and its two tributaries, the Uraricoera (ura 1-3) and Tacutú (tacu) rivers, according to their dissimilarity in bird species composition using the Bray-Curtis index for quantitative data. Light grey dots represent localities from the upper Rio Branco in the savanna domain, whereas black dots represent localities from the lower Rio Branco.
Using quantitative data from our point counts (332 species) we found significant associations with either the upper or the lower Rio Branco in 70 species (∼ 20% of the sample). The Indicator Species Analysis revealed that 40 species were associated with the gallery forests of the upper Rio Branco, and 30 species with the lower portion of the river (Table 1, Figure 3).
Table 1. Avian indicator species in the Rio Branco, based on abundance data of 332 species in 15 sampled localities (See Table S2). We found 70 species with significant associations with either the upper or the lower Rio Branco. For each species, we indicate the correlation value and statistical significance of association (P-value). Species are ordered by level of association, from highest to lowest. Bold species are those included under IUCN (2018) or Brazilian (Instituto Chico Mendes de Conservação da Biodiversidade 2018) threat categories.

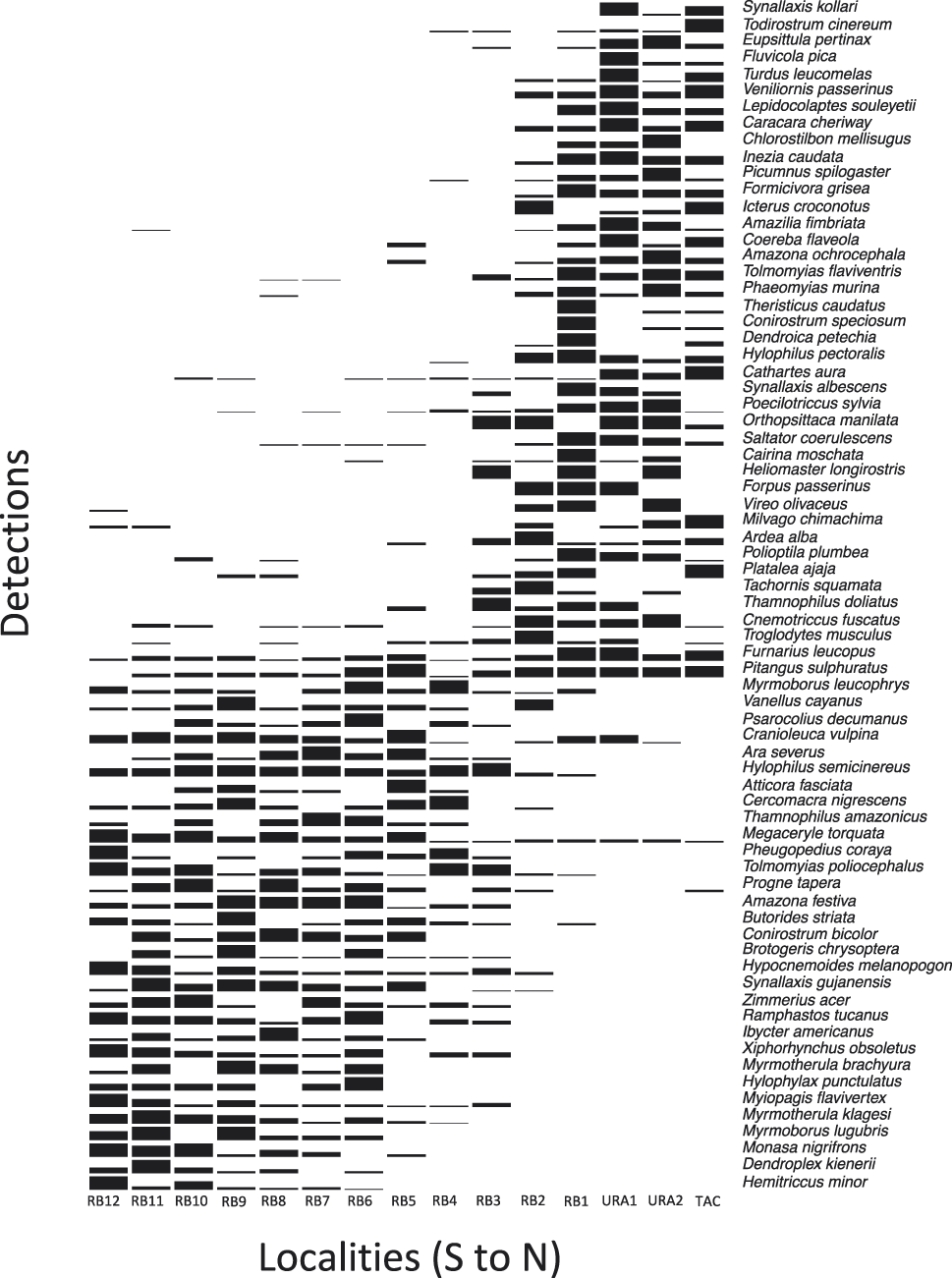
Figure 3. Frequency of bird species significantly associated with the lower or upper section of the Rio Branco according to the Indicator Species Analysis. Localities are organized from the lower Rio Branco (RB12) to the upper Rio Branco (RB1) and its tributaries, the Uraricoera (URA) and Tacutu (TAC) rivers. Bar height represents the number of point counts where a given species was recorded during our systematic surveys. Species with presence on the upper right part of the graph represent species restricted to the gallery forests of the upper Rio Branco, including the Uraricoera and Tacutu rivers; species located at the bottom left of the graph represent those species restricted to the lower Rio Branco Varzea forests.
Habitat association and specificity
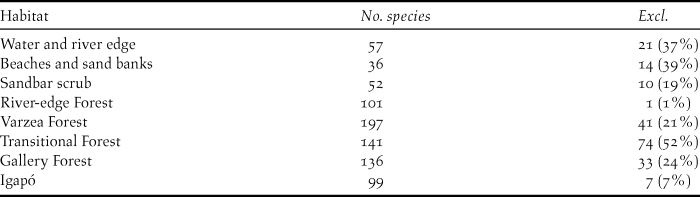
Nearly 46% of the bird species (202) recorded along the Rio Branco and its main tributaries were restricted to single habitats, suggesting a high degree of habitat specialisation within the basin. Transitional forests presented the highest number (52%) of exclusive species within the riverine forests sampled, most of which are common terra-firme forest species. Aquatic environments and beaches and sand banks presented high numbers of exclusive species, reaching 37% and 39% of the species recorded in those habitats, respectively. Gallery and várzea forests also presented high numbers of exclusive species (24% and 21% respectively) (Table 2).
Table 2. Number of bird species observed by habitat along the Rio Branco and its tributaries, including the number of exclusive species associated to each habitat. Habitat data were retrieved from Appendix S1 and is not the result of a fair comparative sampling among habitats, simply representing the results from our inventories along the river.
Threatened species and targets for future monitoring
We recorded 23 bird species that are currently listed under some IUCN (2018) threat category and two that are considered ‘Vulnerable’ under Brazilian law (Instituto Chico Mendes de Conservação da Biodiversidade 2018), totalling 25 species of conservation concern (Appendix S1, Figure 4). Two of these are endemic to the Rio Branco basin and are ‘Critically Endangered’ (the Hoary-throated Spinetal Synallaxis kollari and the Rio Branco Antbird Cercomacra carbonaria); seven are ‘Vulnerable’, and 14 are ‘Near Threatened’ (Appendix S1, Figure 4). The two species considered ‘Vulnerable’ under Brazilian law but not by IUCN are the Lesser Wagtail-Tyrant Stigmatura napensis and the Black-striped Sparrow Arremonops conirostris. Eight of the species of conservation concern also presented significant associations with particular portions of the river (Table 1); two are significantly associated with the upper Rio Branco (White-bellied Piculet Picumnus spilogaster and Hoary-throated Spinetail Synallaxis kollari) and six with the lower Rio Branco (Festive Parrot Amazona festiva, Red-billed Toucan Ramphastos tucanus, Zimmer’s Woodcreeper Dendroplex kienerii, Klages’s Antwren Myrmotherula klagesi, Ash-breasted Antbird Myrmoborus lugubris, and Bicolor Conebill Conirostrum bicolor) (Figure 4).
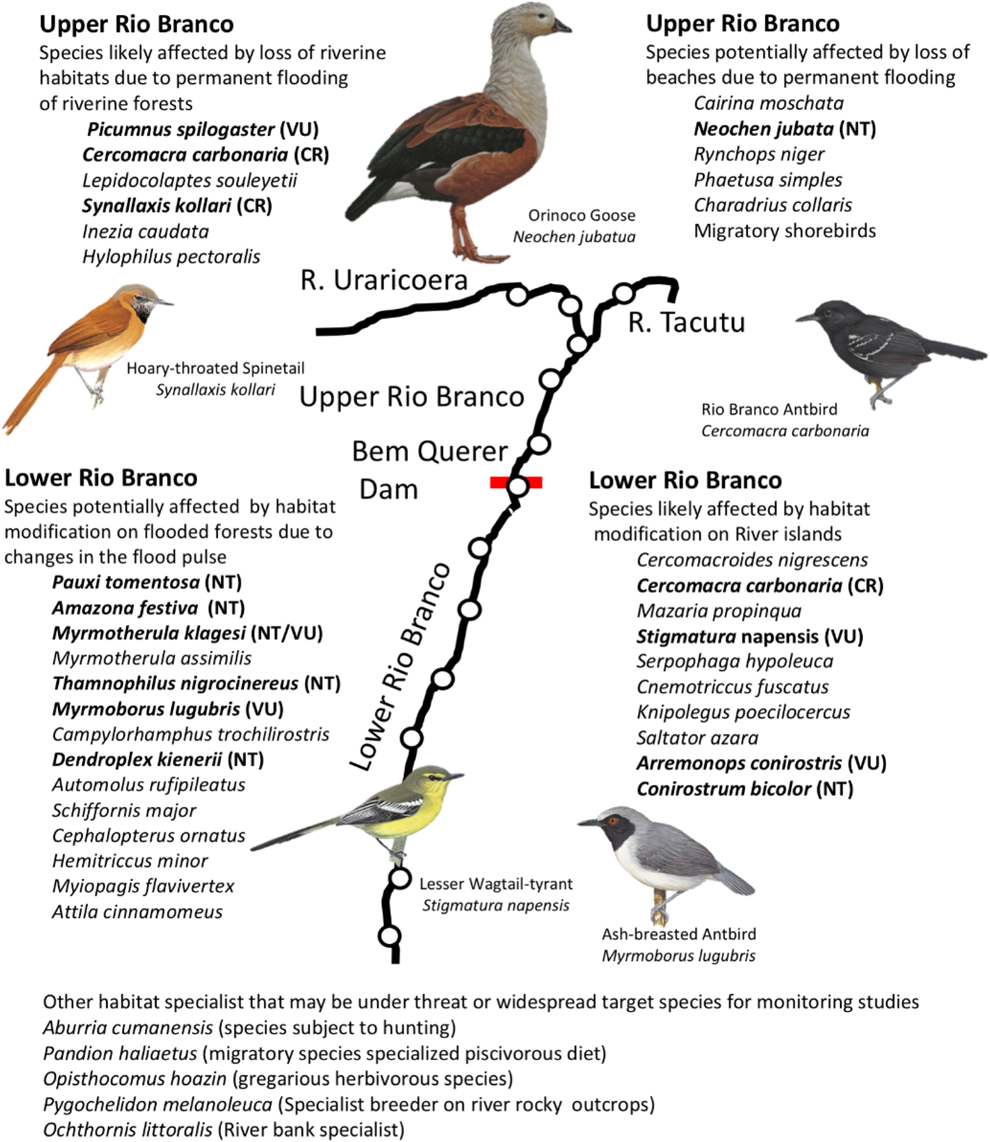
Figure 4. Main threats to the avifauna due to the potential construction of a hydroelectric dam and a hydroway on the Rio Branco. The black line represents the Rio Branco and the white dots represent our study areas along the river. The red line represents the location of the Bem Querer rapids, natural boundary between the upper and lower sections of the Rio Branco, and location of the potential dam. In bold, species under any threat category. Bird illustrations were obtained from the Handbook of the Birds of the World (retrieved from http://www.hbw.com on 20 March 2018) with the permission of Lynx Edicions.
We selected a group of 50 target species that we suggest be considered in future monitoring studies (Appendix S1, Figure 4). These species include 13 species of conservation concern (two ‘Critically Endangered’, four ‘Vulnerable’, and seven ‘Near Threatened’), six northern migrants, and 37 habitat specialists. From the 50 species, 18 are widespread along the river, 21 are restricted to the lower Rio Branco, and seven are restricted to the upper Rio Branco.
Table 3. List of 50 target species suggested for future monitoring studies on the Rio Branco. This list includes species of conservation concern, threatened species, habitat specialists, migratory, and geographically restricted species, whose ranges may expand or retract following changes in their natural habitats.

M Migratory species, CR Critically Endangered, VU Vulnerable, NT Near-threatened.
Discussion
This study represents one of the first attempts to survey the avifauna of an entire river system in the Amazon basin. Three key aspects of this study show the importance of this kind of approach, particularly for rivers where major infrastructure projects may be established, such as hydroelectric dams and hydroways, which may potentially affect them permanently.
First, we have shown that the avifauna found along the Rio Branco and its tributaries represents a very diverse set of species, many of which are currently threatened and are restricted to riverine habitats. We detected around 60% of the bird species ever recorded in the state of Roraima (Naka et al. Reference Naka, Cohn-Haft, Mallet-Rodrigues, Santos and Torres2006), surveying a rather limited number of riverine habitats. It is clear that the river system studied represents a hotspot for avian diversity and conservation with the presence of 25 threatened species, two of which are ‘Critically Endangered’ at the global level (Synallaxis kollari and Cercomacra carbonaria). Nearly 35% of the bird species recorded during our surveys were exclusive to single habitats within the Rio Branco basin, showing a high level of habitat specificity. It has long been known that riverine birds are tightly associated with specific stages of forest succession along the flooding gradient (Remsen and Parker Reference Remsen and Parker1983, Rosenberg Reference Rosenberg1990, Robinson and Terborgh Reference Robinson and Terborgh1997). Specialised birds are likely to be most affected by habitat disruptions, leading to local extinctions if their specific habitats are severely affected, as is the case with dams and hydroways.
Second, this study supports the observations raised a decade ago (Naka et al. Reference Naka, Cohn-Haft, Whittaker, Barnett and Torres2007), suggesting that two distinctive avifaunas inhabit the Rio Branco. One avifauna associated with the savannas and gallery forests of the upper portion of the river, and the other associated with its lower portion. It is particularly important to note that the transitional zone between the upper and lower portions of the river is located at the place where the Brazilian Government plans to build a large dam, named after the Bem Querer rapids, possibly wiping out this entire ecotone region. The pattern observed along the Rio Branco may not be unique in the Amazon, but this represents the first well-described case. Patterns of geographic variation along the flooded forest avifaunas in the Amazon itself have been described by Cohn-Haft et al. (Reference Cohn-Haft, Naka, Fernandes and Albernaz2007), suggesting that geographic variation in the avifaunas along Amazonian rivers may be more common than previously thought. Unfortunately, we lack comparative data from other major Amazonian rivers to compare our results with.
Third, we have shown that only by studying the entire river, which is rarely or never done in environmental impact studies, were we able to fully understand the true threat of disrupting the normal flow of a river such as the Rio Branco. What we have shown here is that it is necessary to describe distribution patterns in an entire river basin, to assess potential threats. Basin-wide approaches have been suggested elsewhere (Lees et al Reference Lees, Peres, Fearnside, Schneider, Jansen and Zuanon2016, Latrubesse et al. Reference Latrubesse, Arima, Dunne, Park, Baker, d’Horta, Wight, Wittmann, Zuanon, Baker and Ribas2017), and it seems the only way to fully understand current threats and make sound and science-based decisions in terms of human interference in Amazonian rivers (Laurance et al. Reference Laurance, Peletier-Jellema, Geenen, Koster, Verweij, Van Dijck, Lovejoy, Schleicher and Van Kuijk2015). This means that by investigating one, two, or even a few localities to assess patterns of avian diversity, we would have failed to understand the real threats to the avifauna following a major infrastructure project.
We argue that before conducting any major infrastructure construction with permanent consequences on biodiversity, it is key to assess the degree of damage that the river and its associated biota may suffer, and such assessment would only be possible after surveying the entire length of a river, including its main tributaries, or an entire river basin (Latrubesse et al. Reference Latrubesse, Arima, Dunne, Park, Baker, d’Horta, Wight, Wittmann, Zuanon, Baker and Ribas2017). Unfortunately, environmental assessment studies are often conducted in restricted areas around projected dams, and most often only above them, where direct impacts such as flooding may occur (Fearnside Reference Fearnside2014, Reference Fearnside2015b). Up to date, no Amazonian river has a geographically complete survey of its biodiversity along its entire course. And this remains true, even on those rivers that have already been dammed, and have received considerable funds for research from electricity companies and infrastructure consortia. We argue that without high-quality baseline data from all biological groups that could be potentially affected by the changes that a hydroelectric dam will bring to an entire ecosystem, there is little hope of understanding the real consequences of such environmental modifications. This kind of study ought to be conducted in free-flowing rivers throughout the basin, and these data could then be used to make decisions on which basins (if any) may be used for energy development, and which should be spared and prioritized for conservation reasons.
Avian conservation on the Rio Branco
The potential construction of a hydroelectric dam and a hydroway along the Rio Branco is a cause for serious concern. The social and biological impacts of transforming free-flowing rivers into chains of reservoirs are yet to be fully understood (Finer and Jenkins Reference Finer and Jenkins2012, Forsberg et al. Reference Forsberg, Melack, Dunne, Barthem, Goulding, Paiva, Sorribas, Silva and Weisser2017, Latrubesse et al. Reference Latrubesse, Arima, Dunne, Park, Baker, d’Horta, Wight, Wittmann, Zuanon, Baker and Ribas2017). We know, however, that major impacts are associated with both the flooding above the dams, and the disruption of the river’s natural seasonal fluctuations downstream, including permanent inundation or irregular unpredictable floods (Sá-Oliveira et al. Reference Sá-Oliveira, Hawes, Isaac-Nahum and Peres2015, Lees et al. Reference Lees, Peres, Fearnside, Schneider, Jansen and Zuanon2016, Assahira et al. Reference Assahira, Piedade, Trumbore, Wittmann, Cintra, Batista, de Resende and Schöngart2017). Although floodplain organisms are adapted to flooding lasting several months each year, they are not adapted to being permanently flooded nor to the disruption of seasonal flooding (Fearnside Reference Fearnside2014). Flood disruption may be particularly damaging for those species adapted to seasonal flooding, causing permanent changes in the structure of the forest and plant species composition. Other indirect impacts begin to be witnessed during dam construction (Alho Reference Alho2011) and include human migration to the dam area, indirect habitat loss, increasing land use changes, and hunting (Lees et al. Reference Lees, Peres, Fearnside, Schneider, Jansen and Zuanon2016). These kinds of impacts (direct and indirect) have non-random effects on biodiversity, potentially filtering out organisms bearing particular traits and allowing the establishment, persistence, and spread of others, precipitating shifts in biological communities, including local extinctions (Díaz et al. Reference Díaz, Lavorel, Chapin, Tecco, Gurvich and Grigulis2007).
We have found that the avifauna of the Rio Branco is rich and diverse, and many stretches of the river are among the most well-preserved in the entire Amazon (Goulding et al. Reference Goulding, Barthem and Ferreira2003). The mere idea of a major dam in one of the least populated and best-preserved rivers of the Amazon basin is of major concern. As we have shown, the river is home to at least 25 bird species of conservation concern, including two ‘Critically Endangered’ and endemic species, which if extirpated from this river, will become globally extinct. Also, eight of the species currently included under any IUCN (2018) threat category also are significantly associated with either the upper or the lower stretches of the Rio Branco and its main tributary, the Uraricoera river (Figure 4). These species are particularly prone to local and even global extinction and should be closely monitored if a dam is eventually built on this river.
The avifauna of the Rio Branco could be impacted by the Bem Querer dam in at least four different ways (Figure 4). For those species restricted to the upper stretch of the river, habitat loss due to permanent flooding above the dam will be inevitable, and the higher the final level of the waters (currently set at 62.5 m, Empresa de Pesquisa Energética 2011), the more habitat will be lost. The ‘Critically Endangered’ Synallaxis kollari is restricted to the upper Rio Branco, and the two rivers that form the Branco, the Uraricoera and Tacutú rivers. Loosing this species’ stronghold along these rivers will severely jeopardise its future. On the upper Rio Branco, another main cause of concern will be the permanent disappearance of sandy beaches, which are the habitat for at least one ‘Near Threatened’ species, the Orinoco Goose Neochen jubatus, and many other aquatic birds. For those species linked to the riverine habitats of the lower Rio Branco, their natural habitats can be affected by the disruption of the flooding pulse of the river, especially if an associated hydroway is also built along this river. The flooding pulse disruption will likely be particularly damaging to river islands and all bird species associated to the several unique habitats found on these islands, such as beaches, sandbar scrubs, and riverine forests. Among the species exclusive to these habitats are several threatened species, such as the Amazonian subspecies of Stigmatura napensis, Arremonops conirostris, and Conirostrum bicolor (Figure 4). Some of these species have been included as ‘Vulnerable’ under the Brazilian law (MMA no. 444/2014), because the construction of dams in the Amazon has already disrupted their habitats elsewhere (Cochrane et al. Reference Cochrane, Matricardi, Numata and Lefebvre2017, Instituto Chico Mendes de Conservação da Biodiversidade 2018). Also, species associated with tall flooded forests may suffer from the disruption of the flood pulse due to the construction of a dam (Resende et al. Reference Resende, Schöngart, Streher, Ferreira-Ferreira, Piedade and Silva2019, Sousa Lobo et al. Reference Sousa Lobo, Wittmann and Piedade2019). Seasonally flooding forests that flood permanently will die, whereas those that do not flood regularly are likely to present major plant species composition changes, with changes in species dominance and flooded forest specialist trees being outcompeted. These changes could directly affect the avifauna, particularly those species tightly associated with tall várzea forests. Among these species are some species of conservation concern, such as the Crested Curassaw Pauxi tomentosa, Myrmotherula klagesi, Blackish-gray Antshrike Thamnophilus nigrocinereus, Dendroplex kienerii (all ‘Near Threatened’), and Myrmoborus lugubris (‘Vulnerable’) (Figure 4).
Therefore, it is clear that the avifauna will be affected at different levels, and in different ways along the Rio Branco. We recommend that future Ecological Assessment Studies (known in Brazil as Estudos de Impacto Ambiental or EIA) are conducted above and below the planned dammed area, to fully evaluate the risks to biodiversity. Given the linear distribution of riverine bird species (whose habitats follow the course of rivers), the degree of habitat loss in relation to species distribution can, and should be, measured. Studies from other biotic groups, such as plants, fish, mammals, reptiles, and amphibians, may shed light into the nature of spatial patterns of biodiversity distribution along the river. We suspect that these groups will show distribution patterns very similar to those found in birds, and that the risks and threats will be comparable. From a conservation standpoint, if all five projected dams for the Brazilian state of Roraima are actually built, these will directly impact two National Parks, two Ecological Stations, two National Forests, and at least two large Indian Territories. Furthermore, the long-term impacts of reduced sediment load and irregular flooding and deposition on rivers further downstream has not even begun to be investigated. In particular, the Anavilhanas Archipelago (the world’s second-largest fluvial island complex and the main feature of a Brazilian National Park) in the Rio Negro, below the mouth of the Branco, owes its existence to the deposition of sediments from the Branco and may have its long-term integrity threatened by altering flooding regimes and sediment retention in the Rio Branco. In sum, we hope that this study will call attention to the evolutionary and ecological world hotspot that is the Rio Branco, trigger interest among other groups to understand the nature of the ecotone zone and the patterns of species replacements along it, and make clear the serious impacts likely caused by damming and alteration of this important and unique Amazonian river.
Supplementary Material
To view supplementary material for this article, please visit https://doi.org/10.1017/S0959270919000133
Acknowledgements
We are grateful to a number of people involved in our many expeditions, including many field assistants, cooks, and boatmen. Among these, we are particularly indebted to Agnaldo de Souza, Maxwell Santos, Hamilton de Melo, and Sebastián Salvino. Several people from the Chico Mendes Institute of Biodiversity Conservation (ICMBio) were key to the success of many of our trips. These include Beatriz Ribeiro, Antonio Lisboa, Romerio Briglia, Antônio Galdino, Renata Bocorny, Erica Tieko Fijisaki, and Inara Santos. We are also grateful to Hevana Lima, Flor Maria Las-Casas Guedes, and Alexandre Curcino for joining us in some of the point counts on the Uraricoera and Mucajaí Rivers. We are also very grateful to the curators of the institutions that hold specimens from the Rio Branco and share their data with us, particularly Alexandre Aleixo, from Museu Paraense Emilio Goeldi. We would finally like to remember our beloved friend Juan Mazar Barnett who participated in one of our first expeditions to the Rio Branco in 2001. This work was supported by Fundação Boticário de Proteção à Natureza (L.N.N. grant number 934-2012.1), the Brazilian Research Council, CNPq (L.N.N. grant numbers 484219/2011-1 and 480496/2012-9), the Fundação Estadual de Meio Ambiente e dos Recursos Hídricos, FEMARH (L.N.N.), the Chico Mendes Institute of Biodiversity Conservation (ICMBio), through the Diretoria de Pesquisa, Avaliação e Monitoramento da Biodiversidade (T.O.L. grant number MPC-014.007), and Rufford Foundation (TOL grant # 19908-1). A. C. P. (2011-2013) was granted a MSc fellowship by the Coordination for the Improvement of Higher Education Personnel (CAPES), whereas T. O. L. and G. R. L. are currently funded by PhD fellowship by the same institution.








