


Introduction
The Little Owl Athene noctua is a farmland bird species that has experienced one of the steepest population declines across its entire European range during the last 60 years, when its distribution became highly fragmented and several populations became locally extinct (Cramp Reference Cramp1985, Tucker and Heath Reference Tucker and Heath1994, van Nieuwenhuyse et al. Reference van Nieuwenhuyse, Génot and Johnson2008). In the Czech Republic, the Little Owl is an endangered species that suffered a population decrease of 59% between 1985 and 2003 (Št'astný et al. Reference Št'astný, Bejček and Hudec2006) and this negative trend still continues (Šálek and Schröpfer 2009). Due to the severe population loss of Little Owls, their protection should be treated as a high conservation priority.
The reasons for the marked decline are discussed in the literature and connected with several factors that include habitat destruction due to agricultural intensification (Cramp Reference Cramp1985, Schönn et al. Reference Schönn, Scherzinger, Exo and Ille1991, Šálek and Berec Reference Šálek and Berec2001), harsh winters (Cramp Reference Cramp1985, Schönn et al. Reference Schönn, Scherzinger, Exo and Ille1991, Bauer and Berthold Reference Bauer and Berthold1996), predation by stone martens Martes foina (Luder and Stange Reference Luder and Stange2001, van Nieuwenhuyse et al. Reference van Nieuwenhuyse, Génot and Johnson2008), disappearance of suitable nesting places (Exo Reference Exo1983), road mortality (Exo and Hennes Reference Exo and Hennes1980, Hernandez Reference Hernandez1988, Génot Reference Génot1995), deaths in water reservoirs, air shafts and chimneys (Génot Reference Génot1995, Bauer and Berthold Reference Bauer and Berthold1996), contamination by biocides (van den Bring et al. Reference van den Bring, Groen, De Jonge and Boseld2003, Zaccaroni et al. Reference Zaccaroni, Amorena, Naso, Castellani, Lucisano and Stracciari2003) and direct human interference (Schönn et al. Reference Schönn, Scherzinger, Exo and Ille1991). Analysis of mortality of adult Little Owls shows two distinct peaks: a winter peak associated with severe winters that had long-standing snow cover and a summer peak connected with exhaustion of birds after the breeding season (Exo Reference Exo1988, Šálek Reference Šálek2004). During the breeding season, birds expend a high amount of energy in nourishing themselves and their offspring (energetic stress) and consequently reach the lowest body mass levels in comparison with other periods of their life cycle (Exo Reference Exo1988, Gassmann and Bäumer Reference Gassmann and Bäumer1993). Similarly, body mass of females before the incubation period is closely correlated with total breeding success (Gassmann and Bäumer Reference Gassmann and Bäumer1993).
Home range size of long-lived sedentary birds should encompass enough feeding sites and adequate resources during the whole year to assure their survival (Newton Reference Newton1979). Animals that occupy territories with a high availability of preferred foraging habitats have smaller home ranges which should affect reproductive and survival rates (Newton Reference Newton1979). Some researchers indicate that grassland habitats such as pastures and mown hay fields are particularly important for Little Owls because they offer a continuous food supply, especially insects, earthworms and rodents, almost all year round (Finck Reference Finck1990, Schmid Reference Schmid2003, Thorup et al. Reference Thorup, Sunde, Jacobsen and Rahbek2010). Orf (Reference Orf2001) recorded that the Little Owl`s home range reached 20.3 ha in grassland habitats and 74.8 ha in arable land. Furthermore, meadows and pastures were over-represented in home ranges of radio-tracked Little Owls in north-east France (Génot and Wilhelm Reference Génot and Wilhelm1993).
In the Czech Republic, the Little Owl occurs in localities that have a significantly larger proportion of grasslands, and are situated at lower altitudes, in an agricultural landscape (Šálek and Schröpfer Reference Šálek and Schröpfer2008). The results indicate that the availability of short-sward grassland patches in Little Owl territories could be a limiting factor to their persistence in Central European farmlands, but information about detailed habitat utilisation during the breeding season is still missing. The goals of this study were to gain knowledge of spatial ecology and habitat selection of radio-tracked Little Owls during the breeding season and to determine the major management actions that would assure its successful conservation.
Methods
Study area
This study was carried out in five villages occupied by the Little Owl in western Bohemia, south-west of the city of Plzeň, Czech Republic, within a 50 km2 area of agricultural landscape (49.40°N, 13.9°E; 350–486 m asl). These sites were chosen based on previous research on the distribution of Little Owls, with a minimal distance between adjacent sites of 2.3 km (average = 3.9 km). In this study area, Little Owls nest in artificial cavities, especially brick pigeon-lofts in old buildings, and its population density reaches 2.9 pairs/10 km2 (Šálek Reference Šálek2004). The study area typically consists of intensely managed fields (45%), forests (32%) and grassland patches (10%). The grasslands are dominated by species of Festuca, Phleum Trisetum, Alopecurus and Lolium, among many others.
Radio-tracking survey and determination of vegetation characteristics
Data on foraging habitat preferences were gathered by radio-telemetry, carried out during the breeding season in 2002. Adult Little Owls were caught in a mist-net using a dummy combined with a territorial call played from a tape-recorder near the nests before the monitoring period (at the beginning of March). Birds were equipped with “back-pack” transmitters (Biotrack TW 4, 3.5 g; battery lifespan of up to 8.5 months). A three-piece Yaggi aerial and an ICOM R-10 wideband receiver were used to locate the individuals. Accurate geographical locations of radio-tracked Little Owls were assessed by triangulation from at least two locations every 15 minutes and were drawn on to 1:5,000 maps. Telemetry fieldwork was carried out during favourable meteorological conditions at intervals from one hour after dusk to one hour after midnight, which coincides with the period of highest foraging activity in Little Owls (Exo Reference Exo1989, Fajardo et al. Reference Fajardo, Pividal, Tringo and Jiménez1998).
In April, five adult male Little Owls were radio-tagged near their breeding sites. Additional tagging was also conducted in May when we equipped one female and two full-grown individuals with radio transmitters, however both young were predated by stone marten Martes foina during the first week and these data were not included in further analysis. To assess habitat preferences, we randomly selected the same number of control locations within Little Owl home ranges. Spatial ecology of Little Owls was recorded during three phases of the breeding period (see also Finck Reference Finck1990, Grzywaczewski Reference Grzywaczewski2009) including: incubation (15 April–17 May), nesting (18 May–28 June) and fledging period (29 June–22 August), which coincide with breeding timing of owls in our study area (J. Hruška unpubl. data).
For each habitat type surrounding Little Owl territory, we assessed the two main vegetation characteristics for each 14-day period: (1) Average vegetation height (AVH) – average height of vegetation measured by tape measure (in centimetres) and (2) Vegetation cover(VC) - estimated as percentage of ground that was covered with vegetation.
Analysis of radio telemetry data and habitat selection
Complete home ranges were defined as 100% minimum convex polygons (MCP). For more precise home range assessment and variation during the breeding season, the kernel density estimator was used (Worton Reference Worton1989) and 95% and 50% kernels were calculated. All home range analysis was performed with the ABODE Kernel Home Range extension for ArcGIS 9.3 (Laver Reference Laver2005). Criteria for estimating home-range size were as follows: kernel - fixed bivariate; bandwidth selection: least-square cross-validation method; grid resolution – resolution 100 (see Laver Reference Laver2005 for more details). Flying distances of Little Owls from nesting/roosting sites to foraging areas were determined by measuring distances in Geoportal COSMC (http://www.geoportal.cz).
For the purposes of habitat selection of the Little Owl, we divided our study area into nine main groups on the basis of ortho-photo-maps using ArcGIS software: fields, forests, tall vegetation, buildings, roads, meadows, pastures, lawns and gardens. Compositional analysis (Aebischer et al. Reference Aebischer, Robertson and Kenward1993) was used to test Little Owl habitat selection at two scale levels: (1) habitat composition within home range was compared to habitat availability in the total study area, and (2) the proportion of habitat used based on number of radio locations compared to habitat availability within the home range. Because compositional analysis is sensitive to the number of habitats (Aebischer et al. Reference Aebischer, Robertson and Kenward1993), we joined all grassland habitats (meadows, pastures/lawns, gardens) into one group and analysed these as one habitat type.
Statistical analyses
Generalised linear mixed effect models (GLMM; lme4 package for the R statistical package) were used to evaluate the effect of progress of the breeding season (particular months, see above) on the size of the home range of the Little Owls. A logarithmic transformation of the entry dataset was used. The GLMM statistic was also applied to data of habitat characteristics preferences of Little Owls. The suitability of each model was assessed on the basis of the minimal AIC criteria, where the best model has the smallest AIC. The individual animal was treated as a factor with random effect to avoid pseudo-replications. All tests were performed in R statistical package (R Development Core Team 2009). The R statistical package was used to compute compositional analysis of habitat selection (see above). We used a randomisation test with 500 repetitions. Habitat that was not found within the particular home range (zero values in entry data matrix) was replaced by 0.01 (see Aebischer et al. Reference Aebischer, Robertson and Kenward1993).
Results
Home range size
In total, we recorded 667 locations from five males and one female radio-tracked Little Owls which were used to determine the home range size and detail habitat use during the breeding season. During the five months of research we lost radio contact with three male Little Owls in the third (M1, M3) and fourth (M4) month of the radio-tracking. The mean home range size differed according to the method used (Table 1). The MCP home ranges were larger than those determined by the kernel method (n = 6). Both the 95% and 50% home ranges varied significantly with the progress of the breeding season (Table 2). The smallest home ranges were recorded during April–June (incubation and nesting period), which expanded during July–August (fledging season; for time division of the breeding season see Methods; Figure 1).
Table 1. Total home range size (ha) of individual radio-tracked Little Owls during breeding season in western Bohemia, Czech Republic (MCP, Kernel UD method, CI – 95% confidence intervals).
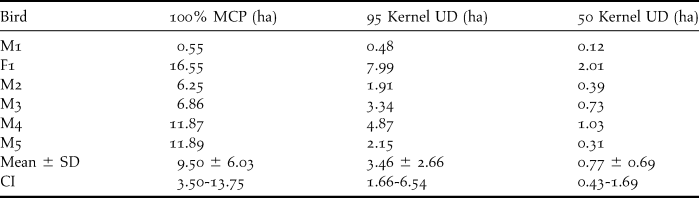
Table 2. Variability of home range (HR) size during the course of the whole breeding period (Generalised linear mixed effect models with Gaussian distribution, log-transformed HR size).

The same pattern was obvious when flying distances from nest/roosting sites were compared during the breeding season (Figure 2). In April, the mean distance from roosts was 64.8 m (n = 84); this increased uniformly during the season up to 211.5 m in August (n = 101). Mean flying distance to the foraging area during the whole breeding season was 106.6 m (n = 667).

Figure 1. 95% and 50% kernel home ranges (ha) of radio-tracked Little Owls during the breeding season in western Bohemia, Czech Republic (means ± SD presented; numbers in parentheses indicate the number of owls monitored each month).

Figure 2. Changes in flight distance to foraging grounds (habitats) from roosting/nest sites of radio-tagged Little Owls during the breeding season (n = 667).
Selection of home range
Habitat composition of home ranges was assessed for 95% kernels. Little Owls slightly preferred certain habitats over others when they established their home ranges within the study area (compositional analysis 1: Wilk’s λ = 0.004, P = 0.06). The two most preferred habitat types were those associated with human settlement, such as buildings and roads. Forest habitat was avoided.
Selection of habitats within the home range
Utilisation of habitats within the home-ranges differed only marginally significantly from habitat availability (compositional analysis 2: Wilk’s λ = 0.001, P = 0.068, Figure 3A). Within the home ranges, grassland habitats and roads were two the most preferred habitat types and buildings the most avoided (Figure 3B).

Figure 3. Habitat characteristics of home ranges of radio-tracked Little Owls and their hunting preferences: (A) habitat use (black bars) compared to availability of habitats (grey bars) within Little Owl home-ranges and (B) habitat selection on the basis of the same data (indexed as log2[use/availability]). Means ± SD are presented.
Variation of habitat use during the breeding season
Utilisation and availability of different habitat types in home ranges changed markedly during the breeding season (Figure 4). In April, Little Owls predominantly hunted in gardens, whereas from May to July they preferred short-sward pastures or lawns. Total utilisation of grassland habitats (pastures/lawns, meadows and gardens) was highest during the nesting and fledging periods (June) and reached up to 90% of all locations, while later in the fledging period (August) these areas were represented less and decreased to 51%.

Figure 4. Seasonal differences in habitat use of radio-tagged Little Owls during the breeding season in western Bohemia, Czech Republic. Means ± SD are presented.
Factors determining habitat use
Two factors, mean vegetation height (AVH) and vegetation cover (VC) were tested for their influence on the selection of foraging habitats of Little Owls. Both vegetation characteristics showed a significant relationship with Little Owl hunting preferences (Table 3). Average vegetation height of Little Owl foraging sites during the breeding season was 4.7 cm (SD = 5.9, n = 667) and total vegetation cover was 51.0% (SD = 34.6, n = 667), in contrast with 31.6 cm and 82.8% respectively in control locations. This pattern held consistently during the whole breeding season (Table 3). Little Owls predominantly hunted from elevated perches (87%), such as trees, poles and buildings that were on average 4.5 m above ground. In August they shifted their foraging activity to cereal fields after stubble ploughing and hunted on the ground (13%).
Table 3. Factors affecting habitat use (on the basis of individual radio-fixes) of Little Owls during breeding season in western Bohemia (results of generalized linear mixed model, AVH = average vegetation height and VC = vegetation cover).

Discussion
Spatial ecology and habitat requirements are crucial aspects of the biology of a species, and as such they may contribute greatly to conservation strategies and management (see e.g. Newton Reference Newton1979). The Little Owl is mainly a sedentary, ground-hunting predator with high site fidelity to year-round territories (Exo Reference Exo, Galbraith, Taylor and Percival1992, van Nieuwenhuyse et al. Reference van Nieuwenhuyse, Génot and Johnson2008). Our study focused on habitat use of Little Owls during the critical period, the breeding season, when birds reach their lowest body mass level during the year (Exo Reference Exo1988, Gassmann and Bäumer Reference Gassmann and Bäumer1993). Home range size during the breeding season is influenced by many factors such as habitat structure and distribution of available food resources (McNab Reference McNab1963, Jenkins Reference Jenkins1981, Mace and Harvey Reference Mace and Harvey1983), which have to provide sufficient energy supplies for owls and their offspring. The mean home range size of 9.0 ha (MCP method) shown in our results is considerably smaller than those reported for Little Owls in other studies across Europe (range 14.5–41 ha; Exo Reference Exo1991, Génot and Wilhelm Reference Génot and Wilhelm1993, Finck Reference Finck1990, Sunde et al. Reference Sunde, Thorup, Jacobsen, Holsegård-Rasmussen, Ottessen, Svenné and Rahbek2009). Our study, however, focused on the breeding season where home ranges reach the smallest size during the whole year (Exo Reference Exo1987, Génot and Wilhelm Reference Génot and Wilhelm1993). During the breeding season, Grzywaczewski (Reference Grzywaczewski2009) reported mean home range size of 19.9 ha in an agricultural landscape and further noted that areas with more grassland habitats contained significantly smaller home ranges (see also Orf Reference Orf2001). High proportion/availability of grassland habitats in Little Owl home range should explain the smaller sizes in our study area. Similarly Exo (Reference Exo1991) and Génot and Wilhelm (Reference Génot and Wilhelm1993) documented that different types of grasslands were proportionally over-represented in Little Owl home ranges.
Our results show a gradual increase in home range size and flying distance to foraging sites during the breeding season. During the incubation period (April–May) the majority of foraging grounds have lower average vegetation height, which is the primary determining factor in habitat choice by Little Owls (see below and also in Šálek et al. Reference Šálek, Riegert and Křivan2010). The availability of short-sward vegetation patches in the vicinity of breeding sites rapidly decreased during the vegetative season and birds were thus forced to fly longer distances. Flying distances during the nesting period are, however, constrained due to the presence of offspring at the nest site. During the nesting and early fledging periods Little Owls significantly preferred mown hayfields, lawns and grazed pastures. Pastures are suitable for hunting as they contain short-sward vegetation throughout the breeding season (see also Finck Reference Finck1990, Exo Reference Exo1991). The rapid increase in home range size during the fledging season (July–August) could be caused by harvesting of crop fields and subsequent stubble ploughing which offer ideal feeding conditions: high abundance of ground-dwelling beetles, the primary prey in our study area (Šálek et al. Reference Šálek, Riegert and Křivan2010), and bare ground (free access to hunting). The offspring, after leaving the nest site, remained in close vicinity of the foraging parents (Šálek pers. obs.) minimising their flight back to the nest. Finally, during the fledging season, male Little Owls reached their lowest level of aggressiveness and defended the smallest territory at any time during the year (Finck Reference Finck1990).
Compositional analysis showed indicative preferences of Little Owls for buildings, roads and grasslands within their home ranges and an avoidance of forest and tall vegetation. The presence of buildings in Little Owl home ranges is connected with the availability of breeding sites. The majority of current Little Owl nest sites were found in man-made structures, especially agricultural objects and residential buildings (Šálek and Schröpfer Reference Šálek and Schröpfer2008) and thus its recent distribution is concentrated around human settlements. Within home ranges, grassland patches are the most preferred habitat type, and together with roads and fields, they were visited significantly more than remaining habitat types. The importance of grassland was found in many previous studies in the Central European agricultural landscape (Loske Reference Loske1986, Dalbeck et al. Reference Dalbeck, Bergerhausen and Hachtel1999, Šálek and Berec Reference Šálek and Berec2001). Similarly, our previous research has shown that localities occupied by Little Owls had a higher proportion of grassland than unoccupied localities (Šálek and Schröpfer Reference Šálek and Schröpfer2008). The higher proportion of roads in the home range could be an important attribute of its territories in residential areas. Roads may be important alternative foraging grounds in periods when vegetation is higher in other habitat types (our results); however their utilisation may be associated with the risk of vehicle collisions (Exo and Hennes Reference Exo and Hennes1980, Hernandez Reference Hernandez1988, Génot Reference Génot1995).
Average vegetation height and cover were the major factors determining the habitat choice of Little Owls during the whole breeding season. In contrast to control locations, Little Owls significantly preferred sparse and short sward vegetation patches within their home ranges which provided access to ground-dwelling prey. In addition, Grzywaczewski (Reference Grzywaczewski2009) found that Little Owls spent 80–95% of their time in short-sward vegetation patches. Although the importance of these structures for Little Owl foraging success is also cited in other studies (Exo Reference Exo1991, Eick Reference Eick2003, Schönn et al. Reference Schönn, Scherzinger, Exo and Ille1991, Fajardo et al. Reference Fajardo, Pividal, Tringo and Jiménez1998) as well as for many other farmland birds (Atkinson et al. Reference Atkinson, Buckingham and Morris2004, Devereux et al. Reference Devereux, McCeever, Benton and Whittingham2004), tall and dense vegetation patches may host a larger diversity and number of invertebrates and vertebrates (Dalbeck et al. Reference Dalbeck, Bergerhausen and Hachtel1999) and thus create “prey refuges” for the surrounding short-sward patches.
Management and conservation implications
Based on recorded data on the spatial ecology of the Little Owl during the breeding season it is obvious that active management has to be directed at the close vicinity of breeding sites. Short-sward grassland habitats, especially pastures with low and sparse vegetation cover, are the preferred habitat types of the Little Owl in their central European farmland territories (see also Exo Reference Exo1988, Finck Reference Finck1990, Šálek and Schröpfer Reference Šálek and Schröpfer2008). If suitable patches with scattered and low sward vegetation are not present, owls are forced to forage in suboptimal or more distant sites, which can result in lower fitness of the adult birds and consequently higher mortality (Exo Reference Exo1988, Thorup et al. Reference Thorup, Sunde, Jacobsen and Rahbek2010). High-quality home ranges of Little Owls consist of a high proportion of small grassland patches with regular grazing and mowing. Although the density of preferable prey (especially ground-dwelling insect) during the breeding season should be higher in patches with permanently tall vegetation, short-sward grassland habitats increase prey accessibility (Romanowski and Žmihorski Reference Romanowski and Żmihorski2008, Šálek et al. Reference Šálek, Riegert and Křivan2010), and its management should be highlighted for conservation of endangered Little Owls. Food limitation during breeding season is thought to be main factor explaining population decline of the Little Owl in Denmark (Thorup et al. Reference Thorup, Sunde, Jacobsen and Rahbek2010). Moreover, emphasis should be placed on the architecture and habitat structures of landscape elements within the Little Owl territory. The Little Owl prefers localities with higher number of small grassland plots over those with few large grasslands (Dalbeck et al. Reference Dalbeck, Bergerhausen and Hachtel1999). Furthermore, linear structures, poles and trees are important as elevated hunting perches (Loske Reference Loske1986, Dalbeck et al. Reference Dalbeck, Bergerhausen and Hachtel1999, Šálek Reference Šálek2004, this study). A fine-scale mosaic of different grassland patches and diverse spatio-temporal dynamics of mowing and grazing could thus provide suitable landscape characteristics and management actions for current populations. On the contrary, recent, most widespread agri-environmental management practices applied to grasslands are characterised by two cuts per year which lead to the homogenisation of grassland areas. Finally, due to the Little Owl’s sedentary lifestyle (short distance dispersion of offspring, and high fidelity of adult individuals) future conservation planning and actions should be focused on occupied areas and their surroundings in order to support existing populations.
Acknowledgements
We would like to thank J. Procházka, J. Hruška, L. Schröpfer, and J. Wertigová for helping with field work. We are also grateful to J. Riegert for helping with statistics, and to R. Klee and M. Kipson for correcting the English manuscript. This study was supported by a grant MSMT6007665801 of the Czech Ministry of Education, research arm of the Research Centre AV0Z60870520 and Grant Agency of the University of South Bohemia 04-136/2010/P.







