


Introduction
Designating effective management areas for threatened species requires knowledge of the factors influencing animals’ spatial ecology (e.g. Chalfoun and Martin Reference Chalfoun and Martin2007, Xu et al. Reference Xu, Zhang, Sun, Zheng, Wang and Zhang2009, Gerber et al. Reference Gerber, Arrigo-Nelson, Karpanty, Kotschwar and Wright2012). Animals that routinely move outside managed areas may be at higher risk than animals whose space use is contained within a protected area (Sutherland Reference Sutherland1998, Woodroffe and Ginsberg Reference Woodroffe and Ginsberg1998, Afonso et al. Reference Afonso, Fontes and Santos2011). While the size of management areas is limited by financial and logistical concerns, wildlife managers should balance feasibility with knowledge of species-specific spatial ecology to implement conservation programmes at an optimal spatial scale.
Although measurement of home range area is important for understanding species’ habitat needs, the home range concept has suffered recent criticism due to the assumption that space use is static (Börger et al. Reference Börger, Dalziel and Fryxell2008, Powell and Mitchell Reference Powell and Mitchell2012). For this reason, some authors question the use of the home range asymptote as a neutral model for home range analysis (Gautestad and Mysterud Reference Gautestad and Mysterud1995) and instead predict more dynamic space use resulting from the complex interplay between individual characteristics and environmental conditions (Wang and Grimm Reference Wang and Grimm2007, Tao et al. Reference Tao, Börger and Hastings2016). While the restriction of activities to a home range can facilitate the exploitation of resources, such as food and nesting areas, nomadic movement patterns may be advantageous when resources are widespread or highly unpredictable (Börger et al. Reference Börger, Dalziel and Fryxell2008). For many species, this may result in variable space use among life stages and across the annual cycle as the costs and benefits of maintaining a home range shift. Relationships between spatial behaviour and habitat variables, breeding status, conspecific density, age, sex, and social status (Sanderson Reference Sanderson1966, Maher and Lott Reference Maher and Lott2000, McLoughlin and Ferguson Reference McLoughlin and Ferguson2000, Jetz et al. Reference Jetz, Carbone, Fulford and Brown2004, Kjellander et al. Reference Kjellander, Hewison, Liberg, Angibault, Bideau and Cargnelutti2004, Mitchell and Powell Reference Mitchell and Powell2004, López-Bao et al. Reference López-Bao, Rodríguez, Delibes, Fedriani, Calzada and Ferreras2014) highlight the complexity of intraspecific variation in home range and movements.
Careful examination of intraspecific variation in space use over time is lacking for many endangered species (Rechetelo et al. Reference Rechetelo, Grice, Reside, Hardesty and Moloney2016). Understanding how spatial behaviour reflects life history is imperative, especially for animals that face high mortality at specific life stages, such as the post-fledging period of altricial birds (Cox et al. Reference Cox, Thompson, Cox and Faaborg2014, Naef-Daenzer and Grüebler Reference Naef-Daenzer and Grüebler2016). Following natal dispersal, space use becomes more extensive and less predictable for many avian species (Mitchell et al. Reference Mitchell, Taylor and Warkentin2010, Penteriani et al. Reference Penteriani, Ferrer and Delgado2011, Small et al. Reference Small, Blank and Lohr2015). In some cases, this coincides with increased mortality risk (Davies and Restani Reference Davies and Restani2006, Weins et al. Reference Weins, Reynolds and Noon2006, Sim et al. Reference Sim, Ludwig, Grant, Loughrey, Rebecca and Reid2013), suggesting that managers should account for the habitat needs and vulnerabilities of dispersing sub-adults. Particularly for species with prolonged juvenile periods (e.g. corvids; Webb et al. Reference Webb, Boarman and Rotenberry2009) or high adult mortality rates, the stability of sub-adult populations is important to consider given the direct impact on breeding populations (Penteriani et al. Reference Penteriani, Ferrer and Delgado2011).
The Mariana Crow Corvus kubaryi is a critically endangered forest bird that faces a high risk of extinction due to elevated adult and juvenile mortality (Ha et al. Reference Ha, Butler and Ha2010). While habitat occupancy of Mariana Crows has been broadly delineated (Morton et al. Reference Morton, Plentovich and Sharp1999, Zarones et al. Reference Zarones, Sussman, Morton, Plentovich, Faegre, Aguon and Ha2015, Faegre et al. Reference Faegre, Ha, Hubl, Ware and Wiitala2016, Kroner and Ha Reference Kroner and Ha2017), little is known about the home range and movement patterns of the species, and intraspecific differences in home range have never been studied. After natal dispersal, and prior to recruiting into the breeding population, sub-adults (average 8–24 months post-fledging) are rarely seen and their behaviour has never been described. Since mitigation of Mariana Crow pre-breeding mortality risk is a conservation priority (Ha et al. Reference Ha, Butler and Ha2010), knowledge of juvenile space use may influence population management.
Evidence from a radio-telemetry study suggests that predation by feral cats Felis catus is one cause of fledgling and sub-adult Mariana Crow mortality (Table 1; S. Faegre and R. Ha unpubl. data). A poorly understood inflammatory disease of unknown cause is also responsible for a large number of deaths of immature birds (T. Work unpubl. data). Habitat management practices for the Mariana Crow population have been focused on removing feral cats within ∼ 350 m of active nests. However, a better understanding of Mariana Crow home ranges and movement patterns during the non-nesting period will help determine if this management strategy is sufficient or if there are larger habitat requirements critical to family groups, and to the survival of young non-breeders. In this study, we: 1) compare home range size and daily movements between radio-tagged fledglings and sub-adults to identify different susceptibilities and habitat requirements between age classes, 2) evaluate the stability of Mariana Crow home ranges and 3) provide an assessment of current Mariana Crow management strategies in light of our results.
Table 1. Description of the 22 Mariana Crows radio-tagged to study spatial behavior.

1 Status at last location: 1 = survived duration of tracking (i.e. tag’s battery life), 2 = Suspected feral cat predation, 3= inflammatory disease, 4 = Cause of death unknown, 5 = Taken into captivity due to injury
2 Tracked opportunistically due to having radio-tagged parents
3 Excluded from fledgling period analysis due to having a tagged sibling
4 Captured as a sub-adult of unknown age (not included in models of home range size, overlap, or daily movement)
Methods
Study system
The single remaining population of Mariana Crows is confined to the island of Rota, Commonwealth of the Northern Mariana Islands, and consists of fewer than 200 individuals (Zarones et al. Reference Zarones, Sussman, Morton, Plentovich, Faegre, Aguon and Ha2015, Kroner and Ha Reference Kroner and Ha2017). Mariana Crows were extirpated from the island of Guam in the 1990s due to predation by the brown tree snake Boiga irregularis (Savidge Reference Savidge1987, Wiles et al. Reference Wiles, Bart, Beck and Aguon2003), an introduced species that does not occur on Rota. Mariana Crows breed as pairs that defend territories around active nests and exclude all conspecifics, including offspring from prior years.
Rota is the second most southerly island after Guam in the Mariana Islands, Western Micronesia (14°09’N, 145°12’E). The 85-km2 island is volcanic in origin with uplifted limestone terraces. The climate is tropical, with high humidity. Wet and dry seasons are typically July–November (wet) and January–May (dry) with rainfall measuring from a minimum of 93.7 mm/month (3.69 inches) in March to a maximum of 339.6 mm/month (13.37 inches) in September (Lander and Guard Reference Lander and Guard2003). Rota is located within the Western Pacific typhoon belt and experiences typhoons periodically.
Approximately 60% of Rota is forested and Mariana Crows are distributed widely throughout secondary and mature limestone and ravine forests (Plentovich et al. Reference Plentovich, Morton, Bart, Camp, Lusk, Johnson and Vanderwerf2005). In 2016, an estimated 4,377 ha of Mariana Crow habitat remained on Rota (based on the exclusion of small, isolated forest fragments from 4,477 ha of forest cover identified from forest service landcover maps; F. Amidon unpubl. data).
Sexing
Crows are not sexually dimorphic and thus were sexed on the basis of their DNA. DNA was extracted from approximately 150 μl of blood taken from the wing vein of each bird. Sex was determined using a PCR-based assay of the absence (male: ZZ) or presence (female: ZW) of the W chromosome (Griffiths et al. Reference Griffiths, Daan and Dijkstra1996).
Radio-tracking
Between March 2010 and January 2017, Mariana Crows were radio-tagged and tracked using Holohil RI-2CT VHF transmitters. Transmitter/harness packages were 3–4% of crow body weight and were fitted to individuals using a backpack with a weak link system incorporated, similar to the design described by Karl and Clout (Reference Karl and Clout1987). The average battery lifetime was 12 months.
Nutritional independence of young from the parents, at an average age of eight months post-fledging (Morton et al. Reference Morton, Plentovich and Sharp1999, S. Faegre unpubl. data), generally coincides with dispersal from the natal territory. Recruitment into the breeding population can occur as early as 16 months after fledging (R. Ha unpubl. data). Crows were classified as fledglings during the period of nutritional dependence on their parents, and as sub-adults after reaching independence from their parents but prior to their first nesting attempt. Due to their movements as family units, fledgling movements approximated the movements of their parents during this same time period. During daily or bi-weekly observations, tagged crows were observed and social interactions were recorded.
This study included 22 Mariana Crows (Table 1). Seventeen were either radio-tagged as fledglings or tracked opportunistically (n = 3) from fledging day through dispersal due to having tagged parents. Since fledglings were almost always with their radio-tagged parents, the relocation data from these individuals is equivalent to that of radio-tagged fledglings. Seven birds that were tagged as fledglings were tracked into the sub-adult period, and three individuals were captured and tagged as sub-adults of unknown age. Based on the primarily pink mouth/tongue coloration and the presence of juvenile plumage, these unknown-age individuals were likely less than one year old at the time of capture (Heinrich and Marzluff Reference Heinrich and Marzluff1992, R. Ha unpubl. data). In three cases sibling pairs were radio-tagged and only one fledgling from the pair was used in each analysis to avoid pseudoreplication. However, both siblings were included in the sub-adult dataset because siblings were rarely seen together as sub-adults (S. Faegre unpubl. data).
Home range analyses
We analysed home range data from all crows that were tracked for a minimum of 30 days, with the exception of one individual which had a non-contiguous home range that could not be accurately measured using the minimum convex polygon (hereafter ‘MCP’) method. We used the fixed K local convex hull method (hereafter ‘LoCoH’; Getz and Wilmers Reference Getz and Wilmers2004) to characterise home ranges from full data sets (consisting of a minimum of 120 data points), and the MCP method (Samuel and Fuller Reference Samuel, Fuller and Bookhout1996) for analyses of data sets over shorter time intervals. While there are biological and statistical disadvantages to the MCP method when samples are large (Samuel and Fuller Reference Samuel, Fuller and Bookhout1996), this method is commonly reported and tends to be more robust with smaller numbers of relocation points. We chose the LoCoH method over kernel density estimators because the LoCoH method represents space use more accurately within landscapes characterised by sharp topographical features and fragmented habitats (Getz and Wilmers Reference Getz and Wilmers2004, Getz et al. Reference Getz, Fortmann-Roe, Cross, Lyons, Ryan and Wilmers2007) such as found on Rota. Furthermore, unlike kernel-based methods, the LoCoH is robust to changes in the smoothing parameter (Getz et al. Reference Getz, Fortmann-Roe, Cross, Lyons, Ryan and Wilmers2007), reducing the potential for biased results and improving the accuracy of inter-individual comparisons. All home ranges were measured at the 100% isopleth.
When possible, we present descriptive statistics for home range size based on complete datasets for biologically meaningful life stages (e.g. fledgling period, sub-adult period). However, due to incomplete datasets for the sub-adult period, we used three shortened temporal intervals (30, 60, and 90 days post-fledging or post-dispersal) when testing for differences in cumulative home ranges between fledglings and sub-adults. Only known-age sub-adults were included in analyses comparing fledglings and sub-adults. Sub-adults of unknown age were included for descriptive purposes in the home range area curves and summary statistics.
We evaluated daily mobility among fledglings and known-age sub-adults using daily movement distances (i.e. the Euclidian distance between observations on consecutive days). We generated home range data-area curves in order to better understand the behavioural processes governing ranging behaviour (Maher and Lott Reference Maher and Lott2000, McLoughlin and Ferguson Reference McLoughlin and Ferguson2000). Area curves are useful for determining the number of relocation points needed to fully reveal home ranges and can also be indicative of home range stability (Haines et al. Reference Haines, Hernández, Henke and Bingham2009).
Statistical analyses
All analyses comparing fledgling (n = 17) and sub-adult (n = 7) spatial behaviour were done using linear mixed effects models in R (package nlme; Pinheiro et al. Reference Pinheiro, Bates, DebRoy and Sarkar2017). Age (fledgling or sub-adult) and sex were modelled as fixed effects and individual identity was modelled as a random effect. To satisfy model assumptions, home range area was square-root transformed whenever it was used as a dependent variable and daily movement distance was cube-root transformed. Home range area and overlap were calculated using Reproducible Home Range package in R (Signer and Balkenhol Reference Signer and Balkenhol2015) and ArcView 10.1 (ESRI 2012).
To determine the average length of time it took fledglings to reach full mobility, we calculated the mean number of days it took fledglings to reach their own mean post-fledging daily movement distance, using only fledglings that were tracked from fledging day through to natal dispersal. For all analyses, we ran one model with all data included and a second with the period of limited fledgling mobility removed, to understand the effects of age class both with and without the physical limitations of the early fledgling period.
We created home range area curves using the program BIOTAS 2.0a 3.8 (Ecological Software Solutions 2004). We examined area curves visually (Laver and Kelly Reference Laver and Kelly2008) to determine whether cumulative areas appeared to approach asymptotes over time.
Results
Home range estimates were 86% larger when using the MCP method, as compared to the LoCoH method (Table 2). Despite the shorter radio-tracking periods for most sub-adults, the home ranges of known-age sub-adults were 78% and 58% larger than fledglings with the MCP and LoCoH methods, respectively (Table 2). There was a high degree of home range overlap between neighbours and sibling pairs (Table 3). It took an average of 31.4 days (SD = 9.13, n = 14) for fledglings to reach their own mean daily movement distance during the fledgling period.
Table 2. Home Range area (ha) of Mariana Crows radio-tracked for their full fledgling period (116–316 days) and sub-adults tracked for variable periods (62–404 days).
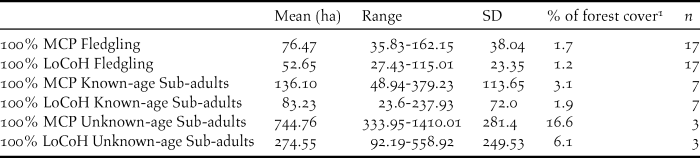
1 The percentage of forested areas on Rota (4,477 ha) covered by each mean home range estimate
Table 3. Percentage of home range overlap between Mariana Crow neighbouring family groups and siblings.

Over 90-day periods, cumulative home range area was larger for sub-adults than for fledglings (F (1, 21) = 31.42, P < 0.001; Figure 1), but was not different between males and females (F (1, 21) = 0.47, P = 0.50). Home ranges also increased in area over time, (F (1, 44) = 120.00, P < 0.001, Figure 1), suggesting that either home ranges were shifting over smaller time intervals, or that movements were increasing over time such that birds were using larger areas. After excluding the first 31 days post-fledging, significant effects of age (F (1, 20) = 14.78, P = 0.001) and time (F (1, 28) = 36.32, P < 0.001) remained, suggesting that these effects were not driven by limited mobility during the early fledgling period.

Figure 1. Increase in cumulative 100% MCP home range area measured over three sequential 30-day intervals (days post-fledging for fledglings or days post-dispersal for sub-adults). Error bars show ± SE.
Non-cumulative home range area was larger in sub-adults than fledglings (F (1, 21) = 13.40, P = 0.002), did not differ by sex (F (1, 21) = 1.39, P = 0.25), but did increase over time (F (1, 99) = 22.28, P < 0.001, Figure 2). After removing the first 31 days post-fledging and re-analysing the data, the effect of age remained (F (1, 20) = 6.65, P = 0.018), but the effect of time was no longer significant (F (1, 84) = 1.50, P = 0.22). Therefore, after the initial month post-fledging, birds used similar amounts of space each month, although they used new areas, thus increasing their cumulative home range.

Figure 2. Non-cumulative 100% MCP home range areas over sequential 30-day intervals (days post-fledging for fledglings or days post-dispersal for sub-adults). Error bars show ± SE.
With all data included, percentage overlap between an individual’s ranges across months was not different between sub-adults and fledglings (F (1, 19) = 0.002, P = 0.96, Figure 3) or between males and females (F (1, 19) = 1.12, P = 0.30). Percentage overlap increased over time (F (1, 75) = 33.93, P < 0.001, Figure 3) and the effect of time remained after removing the first 31 days post-fledging (F (1, 60) = 6.54, P = 0.013). This suggests that fledglings, travelling with family groups, as well as sub-adults, have dynamic, shifting home ranges that lack stable boundaries. The effect of time suggests that initial shifts in home range once fledglings gained mobility were larger than shifts over subsequent months (Figure 3).

Figure 3. Percentage overlap of within-individual 100% MCP home range area over sequential 30-day intervals (days post-fledging for fledglings or days post-dispersal for sub-adults). Error bars show ± SE.
Visual examination of home range area curves also suggest that Mariana Crows have dynamic home ranges (Figure 4). Most fledglings and sub-adults continued to expand into new areas over the study period, resulting in cumulative home range areas that increased over time (Figure 4A). This prevented a meaningful analysis on the minimum number of points needed to accurately estimate home range size. Anecdotally, the three individuals that were tagged as sub-adults of unknown age had home ranges that were larger and increased more rapidly over time than the known-age sub-adults, (dotted lines in Figure 4), suggesting that we might have seen dramatic increases in the home ranges of some of the known-age sub-adults in our study, had we been able to track them longer.

Figure 4. A. Change in 100% MCP home range area over the number of relocation points for each individual. Relocation point 0 represents fledging day (fledglings, solid lines), natal dispersal day (known-age sub-adults, dashed lines), or the day the bird was captured (unknown-age sub-adults, dotted lines). Numbers in parentheses above x-axes indicate the number of days since fledging, dispersal or capture for individuals that were not located daily. Note the variable x- and y-axis scales. B. All individuals shown on the same x-axis scale (days after fledging, dispersal or capture) to highlight differences in the rate of area accumulation between age classes. Note that the y-axis is truncated in order to show area curves clearly for the majority of individuals.
With all fledgling data included, daily movement distances were longer for sub-adults than for fledglings (F (1, 16) = 20.80, P < 0.001), but did not differ by sex (F (1, 16) = 0.027, P = 0.87). Daily movement distances increased over time (F (1, 437) = 190.20, P < 0.001; Figure 5) and age and time interacted (F (1, 437) = 6.51, P < 0.011). After removing the first 31 days post-fledging, the effects of age (F (1, 16) = 5.97, P = 0.027) and time (F (1, 394) = 48.89, P < 0.001) remained. However, the interaction (F (1, 394) = 0.825, P =0.36) was not significant, suggesting that fledgling and sub-adult daily movements increase over time at similar rates.

Figure 5. Average daily movement distance over time in weeks post-fledging (fledglings) or post-dispersal (sub-adults). Error bars show ± SE.
Discussion
Mariana Crows have dynamically shifting home ranges that did not reach area asymptotes (Figures 1-4). When measured cumulatively, Mariana Crow home range size increased over time. However, when non-cumulative home range areas were measured in 30-day intervals for each bird, they did not increase over time. This suggests that monthly shifts in home range caused an increase in cumulative home range areas while area-use on a monthly scale did not increase. While dynamic home ranges were expected for dispersing sub-adults, the lack of stable boundaries in home ranges of fledglings (travelling with their family groups) indicates that, while pairs defend the ever-shifting space around their fledgling(s) from conspecifics, the species does not maintain territories during the non-nesting period. Similar spatial ecology has been reported in other corvids (Rowley Reference Rowley1973, Rolando et al. Reference Rolando, Cavallini, Cursano and Olsen1995, Rolando and Carisio Reference Rolando and Carisio1999, A. Clarke pers. comm.), suggesting that dynamic home range behaviour may be common among wide-ranging, omnivorous birds. For Mariana Crows, the dynamic spatial behaviour of sub-adults and fledglings may be a function of their foraging strategy in a habitat where prey is widespread but where there are also benefits of remaining near a seasonally defended nest area. Dynamic space use during the non-breeding season could indicate that Mariana Crows are relatively flexible and tolerant of non-nesting habitat loss when adjacent habitat is available. However, the lack of stable home ranges in fledglings and sub-adults also highlights the need for large scale management, since improving juvenile survival is critical to species recovery (Ha et al. Reference Ha, Butler and Ha2010).
Sub-adult spatial behaviour
The cumulative home ranges of sub-adults were more than twice the area of family groups (Table 2). Sub-adults may be using large areas after dispersing from their natal territories for a number of reasons, including a need to roam more widely to find resources without intruding on areas occupied by breeding individuals, or to find unpaired conspecifics or vacant nesting habitat. Sub-adults of at least six other Corvus species are also known to range more widely than conspecific adults (Rowley Reference Rowley1973, Heinrich Reference Heinrich1988, Webb et al. Reference Webb, Marzluff and Hepinstall-Cymerman2012) and, given the tendency for non-territorial roaming in sub-adult birds (Penteriani et al. Reference Penteriani, Ferrer and Delgado2011), this is likely true of other species as well.
While sub-adults of other non-cooperatively breeding Corvus species are frequently observed in large aggregations, particularly at rich, ephemeral food sources (Rowley Reference Rowley1973, Heinrich Reference Heinrich1988, Webb et al. Reference Webb, Boarman and Rotenberry2009, Holzhaider et al. Reference Holzhaider, Sibley, Taylor, Singh, Gray and Hunt2011), our observations suggest that sub-adult Mariana Crows may actively avoid conspecifics. Social aggregation of non-breeding individuals may be advantageous in habitats with defendable, patchy resources (Heinrich Reference Heinrich1988, Reference Heinrich2014). In contrast, the frequency of space use is not related to prey availability within home ranges of Mariana Crow family groups (Faegre Reference Faegre2017). Since Mariana Crows forage primarily on widely occurring insects and small animal prey (Jenkins Reference Jenkins1983, Tomback Reference Tomback1986, Faegre Reference Faegre2017), it may be less advantageous to compete directly with conspecifics for food. Indeed, sub-adult Mariana Crows were most often observed foraging and roosting alone and they vocalised infrequently when conspecifics were nearby (R. Ha unpubl. data). While juveniles often appeared reluctant to move away from their parents, they did not appear to be strongly attracted to non-parent conspecifics during the first several months post-dispersal (S. Faegre pers. obs.). For example, in this study, two pairs of siblings that were observed together during 75% and 69% of fledgling observations, were observed together during 4% and 0% of post-dispersal (sub-adult) observations, respectively. Interestingly, home range overlap of these same sibling pairs only decreased from an average of 79% to 59%, between the fledgling and sub-adult periods (Table 3), suggesting that changes in behaviour, rather than loss of home range overlap, explain the low frequency of post-dispersal association between siblings.
The low density of conspecific non-breeders may also be a factor in the lack of sub-adult aggregations among Mariana Crows. Temporary aggregations of 5–15 Mariana Crows were common on Rota during the 1980s (Wiles Reference Wiles1998) when the crow population was approximately 1,350 individuals (Engbring et al. Reference Engbring, Ramsey and Wildman1986), however, such aggregations have rarely been seen since 2007, when the population dropped below 200 individuals (Zarones et al. Reference Zarones, Sussman, Morton, Plentovich, Faegre, Aguon and Ha2015, R. Ha unpubl. data). A similar loss of observed aggregations occurred in Common Ravens Corvus corax in the area around Ithaca, New York, during a period of low population density (1990–2010), and aggregations became more common as the population slowly reestablished itself (A. Clark pers. comm.). Such observations suggest that there may be an Allee effect (Courchamp et al. Reference Courchamp, Berec and Gascoigne2008), if a selection benefit is conferred on sub-adults that are able to associate in groups due to higher conspecific density. While sibling pairs of Mariana Crows in this study made the apparent choice to spend time alone despite ranges that overlapped one another, solitary behaviour could have occurred due to a lack of sufficient additional individuals to gain the benefits of spending time in groups, such as increased vigilance or reduced aggression from territorial individuals. The potential for Allee effects to contribute to decreased sub-adult survival in the dwindling population of Mariana Crows is important to consider.
Family group spatial behaviour
The home range estimates for Mariana Crow family groups tracked for the entire pre-dispersal period (76 and 53 ha for MCP and LoCoH respectively; Table 1) were similar to previous estimates of 64 ha (Morton et al. Reference Morton, Plentovich and Sharp1999). Our MCP and LoCoH home range estimates likely differed from each other because MCP estimates often included large areas of non-habitat, while the LoCoH method more accurately represented the space actually used by Mariana Crows. However, we report MCP home range estimates to be consistent with the majority of the animal home range literature (Laver and Kelly Reference Laver and Kelly2008). The mean MCP and LoCoH home range estimates for various Mariana Crow age classes range from 1.2 to 16.6 % of the island’s total forest cover (Table 2). These observations suggest that cumulatively, Mariana Crow home ranges probably cover most forested areas of Rota.
Like sub-adults, Mariana Crow family groups appeared to be semi-nomadic due to their shifting space use (Figure 3), lack of stable home range boundaries (Figure 4) and high levels of overlap with neighbours (Table 3). Indeed, there were no areas of exclusive use with the home range of family groups. Family groups in more densely populated areas had up to six neighbours (pairs or family groups), each of which overlapped their home range by an average of 45% (S. Faegre unpubl. data). Despite the high levels of overlap from multiple neighbours, neighbouring family groups were rarely observed together (S. Faegre pers. obs.). When neighbours did come together, agonistic interactions between adults were common (S. Faegre pers. obs.). This pattern of avoidance despite high levels of spatial overlap among neighbours may occur frequently among corvids (Rolando et al. Reference Rolando, Cavallini, Cursano and Olsen1995, Rolando and Carisio Reference Rolando and Carisio1999, Holzhaider et al. Reference Holzhaider, Sibley, Taylor, Singh, Gray and Hunt2011, Webb et al. Reference Webb, Marzluff and Hepinstall-Cymerman2012, Loretto et al. Reference Loretto, Reimann, Schuster, Graulich and Bugnyar2016).
Home range dynamics vary according to breeding status in many bird species, with the breeding season often characterised by a decrease in home range area due to territorial behaviour in the vicinity of the nest (Grant and Litchfield Reference Grant and Litchfield2003, Bosch et al. Reference Bosch, Real, Tinto, Zozaya and Castell2010, Lenz et al. Reference Lenz, Bohning-Gaese, Fiedler and Mueller2015). Importantly, we measured home ranges during periods when individuals were not nesting. During active nesting, Mariana Crows have high nest area fidelity. The centroid of the nest area in a given season (i.e. average location of all nest attempts) is located on average 100 m (SD = 112 m) from the centroid of the previous season’s nest attempts. By contrast, the average distance to the nearest neighbouring nest area is 687 m (SD = 427 m). This site fidelity is in stark contrast to the dynamic spatial behaviour we observed and suggests that Mariana Crows maintain stable home ranges when they have an active nest. Opportunistic observations of a notable increase in space use in both Mariana Crows (R. Ha unpubl. data) and American Crows Corvus brachyrhynchos; A. Clark pers. comm.) immediately following nest failure or fledgling mortality suggest that while there may be a period of territoriality during active nesting, dynamic space use is the norm for these and other Corvus spp. (Rowley Reference Rowley1973). Given that effective conservation requires informed management throughout the annual cycle, we recommend future studies of Mariana Crow spatial ecology during active nesting.
Management implications
Following natal dispersal, Mariana Crow daily movements increased by an average of 26% and became more variable with occasional, large movements of up to 1.6 km in a day. The increased energy expenditure of sub-adults, in addition to their shift to nutritional independence, and the risk of encounters with hostile adults, interspecific competitors and predators in novel habitats likely increase the risk of mortality during the first several months following natal dispersal. Low first-year survival in Mariana Crows is a significant threat to species recovery (Ha et al. Reference Ha, Butler and Ha2010), and the combination of environmental stressors and behavioural changes during natal dispersal may play a role in first year mortality. The potential for Allee effects to contribute to decreased sub-adult survival, if aggregating in groups has a survival benefit, should also be considered in future research. Allee effects could be mitigated through captive propagation, if sub-adult birds are released in cohorts that are large enough to form aggregations of 5–15 individuals, as was common historically (Wiles Reference Wiles1998).
Recently, the threat of feral cat predation has been addressed primarily by trapping in the vicinity of active nests (generally within 350 m). Management of threats in nest areas is important, since a successful nest attempt can take up to four months (from nest building to the time the fledglings gain full mobility; R. Ha unpubl. data). However, trapping near active nests can cause disturbance to breeding pairs and may fail to fully protect juvenile Mariana Crows. The five suspected cat predations of immature birds in the study (Table 1) occurred at 92–349 days post-fledgling and three of these occurred at distances > 350 m from the nest area (R. Ha and S. Faegre unpubl. data), likely because family groups shift their space use away from the nest area shortly after fledglings gain mobility (Figure 3). We recommend future study of spatial and temporal variation in juvenile mortality risk, combined with evaluation of whether trapping in the nest area has a detectable effect on the overall feral cat population in crow habitat. Ultimately, high first year mortality (Ha et al. Reference Ha, Butler and Ha2010), much of which occurs 6–12 months post-fledging (Faegre Reference Faegre2017) suggests that larger scale protection measures beyond predator control in nest areas may be necessary to address some of the principal threats to species recovery.
Acknowledgements
We thank the numerous field biologists that assisted in radio-tracking crows for this study: Aaron Wuori, Andria Kroner, Brette Soucie, Calypso Gagorik, Colin Duncan, Cyrus Moqtaderi, Dacia Wiitala, Elizabeth Kain, Emily Cook, Evan Rehm, Gabrielle Robinson, Heather Brown, Henry Fandel, Hillary Henry, Jen Carpenter, Jen Wilcox, Jose Antonio Diaz, Kelle Urban, Kelsey McKune, Laura Bussolini, Lena Ware, Lydia Goy, Marissa Buschow, Matt Henschen, Mike Hitchcock, Phillip Hannon, Rumaan Malhotra, Samantha Lantz, Scott Moore, Sean Jeffreys, Sarena Olson, Sinead Borchert and Steve Seibel. Sexing of our samples was done by Katherine Lofdahl from the University of Guam; Nanda Cortes, Nancy McInerney, and Robert Fleischer from the Smithsonian Institute; Henry Fandel and Andria Kroner from Binghamton University. We thank Manny Pangelinan and Richard B. Seman for their support of our work in the Marianas and Julia Boland, Fred Amidon, Shelly Kremer, Megan Laut, Annie Marshal, Sheldon Plentovich, Paul Radley, and Lainie Berry for additional logistical and permitting support. We thank Michael Beecher, Aaron Wirsing, Andria Kroner, Anne Clark and Aron Faegre for helpful comments on earlier drafts of this manuscript. This work was supported by a National Science Foundation Graduate Research Fellowship to S.F.; it was also supported by the Commonwealth of the Northern Mariana Islands Department of Lands and Natural Resources-Division of Fish and Wildlife from U.S. Fish and Wildlife Section 6 Endangered Species funds, grants F14AF00120, F14AF01145, F15AF01118. Work was conducted in accordance with Federal Fish and Wildlife Permit TE09155B-0 and TE09155B-1, Federal Bird Marking and Salvage Permit 22802 and University of Washington Institutional Animal Care and Use Committee protocol number 2858-04.








