


Introduction
Raptors are wide-ranging, top-level consumers, and are often proposed as potential indicator, umbrella and flagship species (Thiollay Reference Thiollay1989, Primack Reference Primack1993, Simberloff Reference Simberloff1998, Mikusinski et al. Reference Mikusinski, Gromadzki and Chylarecki2001). Their position at the top of trophic food webs, large home ranges and their sensitivity to human disturbance (Newton Reference Newton1979) makes them ideal candidates as biodiversity surrogates, but until recently this had not been formally tested. In a recent examination, however, Sergio et al. (Reference Sergio, Newton and Marchesi2005, Reference Sergio, Newton, Marchesi and Pedrini2006) used the presence and absence of six species of diurnal and nocturnal raptors in Europe as indicator species and found these apex predators were consistently associated with higher species richness of three taxa selected as measures of biodiversity - trees, butterflies and other birds. Given the criticism that the use of indicator species has faced (e.g. Entwistle and Dunstone Reference Entwistle and Dunstone2000) these are important findings that require testing in other areas as originally recommended by Sergio et al. (Reference Sergio, Newton and Marchesi2005, Reference Sergio, Newton, Marchesi and Pedrini2006). We provide such a test here in an area where conservation planning is urgently required because of the high incidence of threatened and endemic flora and fauna – the Cape Floral Kingdom of South Africa (Cowling and Richardson Reference Cowling and Richardson1995). Because our analysis was already underway at the time Sergio et al. (Reference Sergio, Newton and Marchesi2005, Reference Sergio, Newton, Marchesi and Pedrini2006) published their findings, our measures of biodiversity differ from theirs and include small mammals, plants and birds. We set out to investigate if the presence of a globally vulnerable raptor the Black Harrier Circus maurus was associated with higher diversity indices of these three taxa in the Cape Floral Kingdom’s most threatened habitat – renosterveld.
The Black Harrier is one of the few avian endemic species of the Cape Floral Kingdom (von Hase et al. Reference von Hase, Rouget, Maze and Helme2003) and its core breeding area lies within this hugely plant-rich biome of south-western Africa (Curtis et al. Reference Curtis, Simmons and Jenkins2004, Simmons et al. Reference Simmons, Curtis, Jenkins, Hockey, Dean and Ryan2005). In the southern Overberg region, Black Harriers are found nesting almost exclusively in large renosterveld patches (Curtis Reference Curtis2005). Where nesting occurs outside these large (> 100 ha), relatively undisturbed patches, only smaller patches with high connectivity with other patches are used (Curtis Reference Curtis2005). We tested whether the presence of harriers indicated higher biodiversity value than similar sized patches unoccupied by harriers (controls).
Biodiversity value was measured in terms of (i) small mammal species richness and abundance, (ii) bird diversity and abundance and (iii) species richness of rare and endemic plants. This study adds data to the hotly debated topic (Kery et al. Reference Kery, Royle and Schmid2008, Roth and Weber Reference Roth and Weber2008, Sergio et al. Reference Sergio, Newton and Marchesi2008a) of whether top predators can be used as indicators of biodiversity from more diverse regions of the world.
Materials and methods
Study area
Renosterveld is the most threatened habitat type in the Cape Floral Kingdom (Low and Robelo Reference Low and Robelo1996), itself one of the world’s richest and smallest floral hotspots (Myers et al. Reference Myers, Mittermeier, Mittermeier, da Fonseca and Kent2000). This biome lies completely within South Africa and supports at least 8,600 endemic, mainly flowering, plants (Cowling and Richardson Reference Cowling and Richardson1995). Renosterveld is essentially a grassy shrubland, with an exceptionally high diversity of geophytes (bulbous plants), often dominated by the shrub renosterbos Elytropappus rhinocerotis Less., that occurs on moderately fertile, shale-derived soils along the west and south-west coastal lowlands (Kemper et al. Reference Kemper, Cowling and Richardson1999). Structurally it ranges from grassland to shrubland, depending on fire regime and/or grazing pressure, and was formerly considered to have a higher component of grasses (such as Themeda spp.) than shrubs (Newton and Knight Reference Newton and Knight2004). Renosterveld has been reduced drastically by expanding agriculture, mainly cereal and pasture crop production, to < 10% of its original extent (von Hase et al. Reference von Hase, Rouget, Maze and Helme2003).
All patches of renosterveld studied in the Overberg were located between Botriver (34°13’S 19°12’E) in the west, Bredasdorp (34°34’S 20°10’E) in the south and Heidelberg (34°06’S 20°59’E) in the north-east (Figure 1).
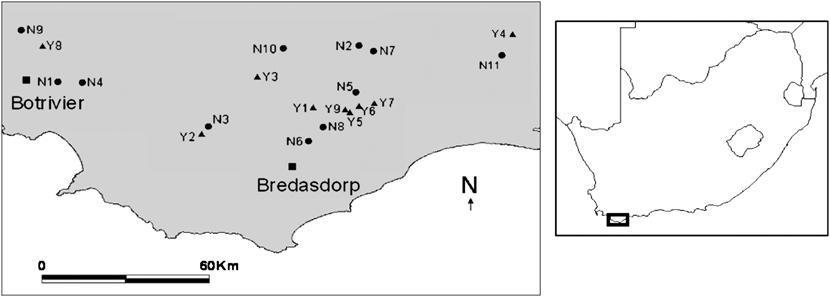
Figure 1. Sites within the Overberg region of South Africa, selected for this study. ▲ sites with breeding black harriers (Y1-9) and ● sites without breeding harriers N1-11)
Average rainfall ranges from 350 to 600 mm per annum (Kemper et al. Reference Kemper, Cowling and Richardson1999) and varies from west to east; winter (May–July) rainfall occurs in the west and a bimodal spring-autumn rainfall regime occurs in the east (Kemper et al. Reference Kemper, Cowling and Richardson1999, Low and Robelo Reference Low and Robelo1996). Mean annual rainfall is 14% higher in the west than the east (436.4 mm versus 381.8 mm; South African Weather Service). The mean minimum annual temperature is 12°C and the mean maximum annual temperature is 24°C (SAWS 2007).
Habitat patch selection
Unlike Sergio et al. (Reference Sergio, Newton and Marchesi2005) we compared the biodiversity of harrier-occupied patches with those of nearby unoccupied patches in the same habitat rather than randomly chosen areas of same habitat. We did so because of the rarity of renosterveld, and the wish for a matched-pair type design to reduce the influence of other variables. No occupied patches were so close to unoccupied ones that harriers, small mammals or passerines might regularly pass between them.
Patches were selected using a GIS data layer from the South African National Biodiversity Institute’s (SANBI) database on the basis of previous knowledge of patches where Black Harriers were known to breed (Curtis Reference Curtis2005). In that survey, patches had been randomly chosen (all sizes) and the larger ones found to contain breeding Black Harriers significantly more often than smaller patches (Curtis Reference Curtis2005). To increase sample size, new patches were identified in the same manner and these were observed for a minimum of three hours to determine the presence of breeding harriers (Curtis Reference Curtis2005). Nine years of monitoring Black Harriers has shown that three hours is sufficient to reveal the presence of a nest through food passing activities (R. E. Simmons unpubl. data). Where possible, patches of 100 ha or larger were surveyed but two patches smaller than 100 ha (one occupied and one unoccupied), were included due to their connectivity to other patches and the presence of breeding harriers. The data were collected over two harrier breeding seasons (2005 and 2006) and a site was classed as active if harriers bred in either of the two years. In total nine patches with Black Harriers (hereafter ‘occupied patches’) and 11 patches without Black Harriers (‘unoccupied patches’) were chosen for biodiversity assessment.
Bird transects
Renosterveld is a low, relatively two-dimensional habitat, therefore the line transect technique (Bibby et al. Reference Bibby, Burgess, Hill and Mustoe2000) was used to survey birds. Transects were walked over a variety of slope aspects and habitats. Each transect was 200 m long except where patch topography disallowed this. In total 4 km of transect per patch were carried out. The length of each transect was determined by recording the start and end points using a Garmin Global Positioning System (GPS) accurate to about 10 m. Transects were carried out during breeding seasons in November and December 2005 and 2006. All patches were surveyed once and all birds within 50 m of each side of the transect line were counted. Although noted, raptor species (including Black Harriers) were not used in the data analysis. Species were identified by sight and/or call. All transects were walked in the early morning or late afternoon, avoiding the middle of the day when birds are known to be less active (Bibby et al. Reference Bibby, Burgess, Hill and Mustoe2000). These data were used to calculate relative abundance measures,
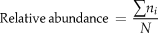 $${\rm{Relative}}\,{\rm{abundance}}\, = \,{{\sum {n_i } } \over N}$$
$${\rm{Relative}}\,{\rm{abundance}}\, = \,{{\sum {n_i } } \over N}$$where ni is the number of times the species occurred and N the total number of species along the transect.
Transects were carried out by three observers. We assessed inter-observer bias by comparing the species detected by each individual within the same two patches (one occupied and one unoccupied). Performing a Kendall Coefficient of Concordance on a Friedman ANOVA showed no significant difference between observers (P = 0.08). Therefore all bird transect data were compared for other variables.
Small mammal trapping
Six lines of 10 Sherman live traps each (8 cm wide by 9 cm high by 30 cm long) were set out for four consecutive nights in 14 patches from 4 October to 28 November 2005 and at 10 patches from 23 September to 15 November 2006. Transects were placed to take advantage of different slopes and were a minimum of 200 m apart. A trapping period was defined as the time between setting the trap and checking the trap. In this study there were 240 night trapping sessions and 240 day trapping sessions per 24 hours. Non-target captures and traps found closed but empty were counted as a half trap period (Sutherland Reference Sutherland and Sutherland1997).
Traps were placed 20 m apart and baited with peanut butter, rolled oats and oil (Dippenaar Reference Dippenaar1974). Crushed maize was used to ground bait for trap-shy species (Sutherland Reference Sutherland and Sutherland1997). Thermal insulation was provided inside the trap. Traps were checked at dawn and dusk. Captured animals were fur clipped, weighed, sexed and recorded as a new or recaptured animal and released at the point of capture. Non- target species (e.g. lizards) were recorded and released at point of capture. Captures were converted into trap success per 100 trap nights where:
Trapping success = (number of captures / No of traps x trapping period in days) x 100.
Because small mammal populations often fluctuate on a year to year basis (David and Jarvis Reference David and Jarvis1985) we re-trapped four patches sampled in 2005 (two occupied and two unoccupied) again in 2006. There was no significant difference between species richness and trapping success on each patch over the two years (T = 3, 10, 3, 2; P = 0.465, 0.917, 0.465, 0.593), data were therefore pooled between years.
In 2005, day trapping was no longer possible after 25 October due to high temperatures and the increased risk of trap mortality (Atyeo Reference Atyeo2006). A regression was developed to predict the day-time capture rate of striped mouse Rhabdomys pumilio from that date onwards. Using patches where both day and night captures had taken place we predicted the success rate per 100 trap periods during the day as a function of the trap success overnight. Predicted R. pumilio trap success during the day = 1.2823 (R. pumilio trapping success at night) -2.842 (R 2 = 0.8797, P < 0.05). Adjusted trapping success for R. pumilio was calculated for five patches. In 2006 the temperature remained below 30°C throughout the trapping period and trapping was done throughout the afternoon session.
Plant transect data
All plant data were collected by a professional botanist (B. A. Walton) by walking two 50 m transects in each patch. Transects were placed down slopes as opposed to across slopes and data were collected every 2 m. Due to the very high species richness, all plant species within a 10 cm radius of the point were recorded and the height of each plant measured to the nearest centimetre.
The number of species recorded for each site was converted to relative abundance using the expression
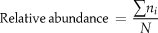 $${\rm{Relative}}\,{\rm{abundance}}\, = \,{{\sum {n_i } } \over N}$$
$${\rm{Relative}}\,{\rm{abundance}}\, = \,{{\sum {n_i } } \over N}$$where ni is the number of times the species occurred and N the total number of species along the transect.
The vegetation height along each transect was calculated by taking the average vegetation height every 2 m. Vegetation density was calculated by taking the number of plants occurring within the 10 cm radius at each point.
Samples of unknown species were collected and identified using herbaria or by consultation with other specialists. Species were identified as rare using the Interim Red Data Lists of South Africa Plant Species (SANBI 2009). Species were identified as endemic to the Cape Floristic Region (CFR) and endemic to the Overberg from plant lists supplied by CREW (Custodians of Rare and Endangered Wildflowers, SANBI). Species endemic to the Overberg were defined as species occurring between Elgin and the Breede River but excluding Elim Fynbos (SANBI 2007). Plant data were not collected at two of the 20 patches; one occupied patch burnt after the 2005 field season and we were unable to contact the owner of an unoccupied patch to gain access.
Data analysis
Species richness and Shannon-Wiener and Brillouin diversity indices were calculated for small mammal, bird and plant species assemblages at each site. Both indices are sensitive to the number of rare species in the sample, and both will increase as the number of rare samples increases (Krebs Reference Krebs1999). The Shannon-Wiener index has been widely used in other studies of small mammal assemblages in South Africa (e.g. Joubert and Ryan Reference Joubert and Ryan1999). The measure of evenness based on the Shannon-Wiener Index was also calculated (Stiling Reference Stiling1999). Dominance was calculated using Simpson’s D statistic.
Data were tested for normality (Kolmogorov-Smirnov test d-statistic). When data were not normally distributed, transformations were attempted to induce normality (Zar Reference Zar1999). Where this was not possible non-parametric tests were used. Data were analysed using Statistica 7.0 (Statsoft 2004).
To determine if Black Harriers are biodiversity indicators, each taxon was compared for higher species richness and biodiversity indices compared between occupied and unoccupied sites. Within birds, species were further separated into feeding guilds and their relative abundances compared. Within small mammals overall abundances were compared between sites. These were compared using Student’s t-tests or Mann-Whitney U tests. We tested if more rare and endemic plant species were present on sites occupied by harriers.
To examine if patches occupied by Black Harriers were similar to each other and significantly different from patches not occupied by Black Harriers, multivariate analyses of bird, small mammal and plant data were undertaken using cluster analysis (CLUSTER) and non-metric multi-dimensional scaling programmes (MDS) in the software package PRIMER 6.0 (Plymouth Marine Laboratory UK). Between-site similarity was assessed using the Bray-Curtis similarity co-efficient. Cluster analysis dendrograms reflecting the hierarchical relationship of patches were produced using group-average linking. SIMPER (Similarity percentages) analysis was also carried out to determine which species contributed most to dissimilarity between patches.
Average plant height and overall vegetation density were compared between patches using t-tests.
Abundance data from birds, small mammals and plants were combined and used to calculate overall diversity indices. These were compared and a multi-dimensional scaling plot of all data produced.
Results
Bird diversity
Across all patches, 67 different species of bird were recorded. The most commonly occurring species on both sites were Grey-backed Cisticola Cisticola subruficapillus (21.8 % of all birds recorded across 20 patches), Karoo Prinia Prinia maculosa (10%), Cape Bunting Emberiza capensis (9.1 %) and Karoo Scrub-robin Cercotrichas coryphaeus (4.8 %). Overall, the relative abundance of birds was 3.5 times higher on occupied patches (4.4 ± 4.7 birds ha-1) than on unoccupied control patches (1.3 ± 3.9 birds ha-1). This difference was not, however, statistically significant (U = 31, P = 0.160) due to high variance.
There was a higher relative abundance of all feeding guilds except nectarivores on Black Harrier patches. These differences were not statistically significant (Table 1). There was a significant difference between species richness of avian carnivores between occupied and unoccupied patches, but other feeding guilds had similar species richness.
Table 1. Relative abundance and species richness of bird guilds on occupied vs unoccupied patches. Significant differences are shown in bold.
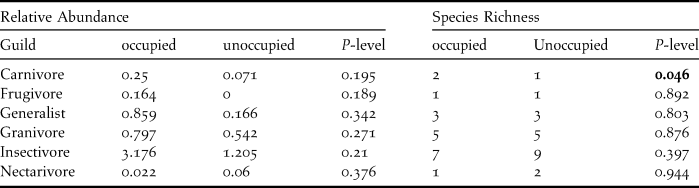
Of the 67 bird species recorded, only the Yellow Bishop Euplectus capensis had a significantly higher relative abundance on occupied patches (0.22 occupied vs 0.02 unoccupied, P < 0.05).
We found no overall significant difference between occupied and unoccupied patches for bird species richness, diversity, dominance and evenness (Table 2).
Overall, there was no pattern for higher relative abundance of birds on patches occupied by breeding harriers. Avian species richness across the occupied and unoccupied patches was similar.
Table 2. Avian species richness and diversity indices for occupied versus unoccupied and east versus west patches. Results in bold are significant.

1 Indicates Mann-Whitney U test was carried out, all other tests were t-tests.
Small mammal diversity
Across all patches, 10 different species of small mammal were caught. Seven species occurred on both occupied and unoccupied patches. One species occurred exclusively on occupied patches and two exclusively on unoccupied patches. Overall the most commonly caught species was the striped mouse R. pumilio, comprising over 70% of all small mammal captures. Abundance of all species (small mammals per 100 trap days) was lower on occupied patches but these differences were not significant (Table 3). There was a higher trapping success for three species on occupied patches and four on unoccupied patches. Surprisingly, diurnal species (prey items, R. pumilio and Otomys irroratus) were slightly more abundant on unoccupied patches (Table 3).
Table 3. Median abundance (small mammals per 100 trap days) by species and overall trapping success for occupied versus unoccupied patches.
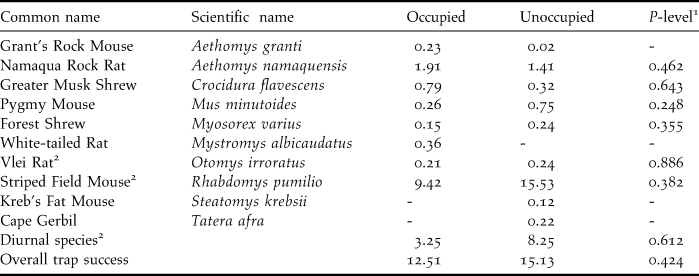
1 Mann-Whitney U test. Where tests are not reported species occurred at too few patches.
2 Diurnal species preyed on by Black Harriers.
Relatively high values of dominance, and low values for evenness were apparent across most patches. This was due to the high abundance of R. pumilio. There were higher diversity values on occupied patches but these differences were not statistically significant because of the low number of species involved (Table 4).
Table 4. Species richness and diversity indices for small mammals in occupied versus unoccupied patches.

1 indicates Mann-Whitney U test was carried out; all other tests were t-tests
Black Harriers breed on patches that have a lower abundance (in terms of captures per 100 trap sessions) of small mammals than unoccupied patches. In this study, vegetation density (measured as the number of plants per point along a transect) was significantly lower on patches occupied by Black Harriers. This suggests that small mammals are more available to Black Harriers on these patches despite the lower small mammal density. However, as a measure of the biodiversity value of patches occupied by Black Harriers, small mammal species richness was not significantly higher on harrier-occupied patches.
For red-listed species there was no trend – the only such species, the white-tailed rat Mystromys albicaudatus occurred at one occupied site.
Plant diversity
Transects for plants recorded 308 species in 60 families over 18 patches. There was no significant difference between the number of species endemic to the Overberg (P = 0.391), and species on the IUCN Red List (P = 0.409) present on occupied and unoccupied patches. All occupied patches contained at least one Red List or endemic plant species (Table 5). Themeda trianda Forssk. was more abundant on occupied versus unoccupied sites (3.36 versus 0.88). This difference however, was not significant (t = -1.179, df = 8, P = 0.272).
Table 5. Number of plant species endemic to the Overberg, endemic to the Cape Floristic Region (CFR) or on the IUCN Red Data List found on occupied versus unoccupied patches

1 Has national Rare status
There was a significant difference in vegetation height (49.31 cm) on occupied patches compared to unoccupied patches (38.87 cm: t = -4.87897, df =934, P < 0.001).
We also found a significant difference in plant density between occupied and unoccupied patches. Unoccupied patches had an average of 3.28 plants per point while occupied patches had an average of 2.88 plants per point (t = 4.152, df = 934, P < 0.001).
We found no other specific difference in occupied versus unoccupied patches in terms of plant species richness, evenness, dominance, or diversity indices. Diversity indices were higher for unoccupied patches suggesting higher species richness (Table 6).
Table 6. Plant species richness and diversity indices for occupied versus unoccupied and east versus west patches.

1 Mann-Whitney U test was carried out, all other tests were t-tests
To our surprise, plant species richness was lower on occupied patches than on unoccupied patches. This may be a function of lower plant density on the occupied patches. Themeda grass T. triandra abundance, however, was higher on the occupied patches. It is thought that renosterveld was once dominated by Themeda grasses and overgrazing has led to present-day domination by bushes such as renosterbos E. rhinocerotis (Cowling et al. Reference Cowling, Pierce and Moll1986). As such, high abundance of Themeda is considered by botanists to be a sign of good quality renosterveld which has not been overgrazed or degraded (Cowling et al. Reference Cowling, Pierce and Moll1986). A grassy layer is also associated with high small mammal abundance in montane areas of western South Africa (Bond et al.1980). In effect harriers may be indicative of pristine renosterveld patches rather than of highly diverse patches.
Vegetation height was significantly higher on occupied patches compared to unoccupied patches. Because nest site selection is very important for ground nesting harriers this result is expected given that they face higher predation rates than tree-nesting species (Simmons Reference Simmons2000). Black Harriers also select nesting patches in thick vegetation to conceal the nest (Chadwick Reference Chadwick1997, Simmons et al. Reference Simmons, Curtis and Jenkins1998, Simmons Reference Simmons2000, Curtis et al. Reference Curtis, Simmons and Jenkins2004).
Rare and endemic plants
This study showed little evidence of a link between Black Harrier presence and a higher abundance of rare and endemic plants. This was contrary to initial analyses (Stanway and Raimondo Reference Stanway and Raimondo2006) using previous plant data from the Overberg. Six Black Harrier patches and six unoccupied patches had been sampled to varying degrees. Their data showed Black Harriers occurred on patches with double the number of rare and double the number of endemic plant species. However, there was a significant sampling bias towards harrier-occupied patches, with 36 sampling records compared to 17 for unoccupied patches (Stanway and Raimondo Reference Stanway and Raimondo2006). This is unlikely to be patch-size related because Kemper et al. (Reference Kemper, Cowling and Richardson1999) showed that fragments less than 1 ha are very similar in plant species composition and diversity relative to large fragments (> 30 ha). Instead we suggest that different methods used in this study (two 50-m transects per patch within the same season) relative to the CREW methods (sampling over a wide area focussing on the rare and endemic plants, year after year), can explain why our methods did not yield the harrier association with rare and endemic plants found by CREW.
We suggest that the lack of association found between harriers and rare and endemic plants may have been masked by inappropriate sampling and our wish to keep transect-sampling for birds, mammals and plants similar. This will be explored in future years.
Cluster analysis
Analysis of bird, rodent and plant data showed no grouping of occupied and unoccupied sites, suggesting all sites were similar.
Combined diversity
The diversity indices and species richness for birds, plants and small mammals combined showed that although species richness (and dominance) may be lower on occupied patches the diversity and evenness indices were higher. This suggests that occupied patches are more diverse and less dominated by commonly occurring species.
Discussion
Given that raptors are top predators, sensitive to changes in prey base and disturbance and have recently been shown to be indicative of high biodiversity value in Western Europe, this analysis assessed whether Black Harriers might also be used as an indicator of biodiversity value in South Africa. We assessed this within renosterveld patches in the highly fragmented Overberg in terms of the abundance and species richness of (i) birds and (ii) small mammals and (iii) the presence of rare or endemic plant species.
Biodiversity vs naturalness in a fragmented habitat
This study did not assess Black Harrier presence in an area of unbroken habitat, unlike the other studies of raptor as indicators of biodiversity (Sergio et al. Reference Sergio, Newton and Marchesi2005, Reference Sergio, Newton, Marchesi and Pedrini2006, Ozaki et al. Reference Ozaki, Masahiro, Kawahara, Iida and Fukuyama2006). Instead we looked at discrete patches of natural vegetation in an agriculturally transformed matrix. Our results may be confounded by the fact that diversity per se is a poor measure of the effect of fragmentation (Kemper et al. Reference Kemper, Cowling and Richardson1999). More important differences may be found in changes within community structure as shown by the frequency of individuals and species with different biological attributes (Saunders et al. Reference Saunders, Hobbs and Margules1991, Holt et al. Reference Holt, Robinson and Gaines1995, Kemper et al. Reference Kemper, Cowling and Richardson1999). The higher abundance of Themeda trianda on patches used by breeding harriers may also suggest that these occupied patches contain more pristine and intact natural renosterveld, independent of overall diversity values.
Black Harriers as biodiversity indicators
Indicators of biodiversity have been proposed as a potential tool for selecting areas for conservation when information about species distributions is scarce (Flather et al. Reference Flather, Wilson, Dean and McComb1997, Lawler et al. Reference Lawler, White, Sifneos and Master2003). The use of raptors as indicators of biodiversity has shown mixed results. Ozaki et al. (Reference Ozaki, Masahiro, Kawahara, Iida and Fukuyama2006) found no relationship between the diversity of bird, butterfly, carabid and native forest-floor plant species and the presence or absence of Northern Goshawks Accipiter gentilis. Roth and Weber (Reference Roth and Weber2008) found that while raptors in Switzerland were reasonable surrogates for biodiversity measures there, Parus tits were better indicators of insect biodiversity. Sergio et al. (Reference Sergio, Newton and Marchesi2005, Reference Sergio, Newton, Marchesi and Pedrini2006) however, found a significant relationship between abundance and diversity of birds and butterflies and the presence or absence of a raptor.
Our results do not show any significant correlation between the presence of Black Harriers and higher levels of biodiversity in similar sized patches. We may have been disadvantaged by the small sample size (nine occupied patches and 11 unoccupied patches), which was limited by the number of similar-sized and nearby patches that were used as controls. Under-sampling of multiple taxa may be a common problem for revealing associations between top predators and indices of diversity (Sergio et al. Reference Sergio, Newton and Marchesi2008a). We also chose a bird that is naturally rare, in need of conservation attention and does not occur at the same levels of abundance enjoyed by the six species assessed by Sergio et al. (Reference Sergio, Newton, Marchesi and Pedrini2006) or the Northern Goshawk assessed by Ozaki et al. (Reference Ozaki, Masahiro, Kawahara, Iida and Fukuyama2006).
Given our results, it may be that harriers are sentinel species linked to the lack of disturbance in the patch rather than the overall biodiversity. Apex predators at the top of the food chain are more vulnerable to alterations of their supporting ecosystem (Sergio et al. Reference Sergio, Caro, Brown, Clucas, Hunter, Ketchum, McHugh and Hiraldo2008b). Therefore conservation managers could use the presence of Black Harriers as a heuristic indicator of ‘pristineness’ or naturalness in this mega- diverse region. They can also use the future presence of Black Harriers to monitor the progress of the restoration of any patches that are protected in future. Therefore Black Harriers are not indicators of biodiversity of renosterveld patches. However their vulnerable status means their presence on a patch should not be ignored by conservation managers.
Acknowledgements
We thank Ben Walton for his meticulous plant surveys of the renosterveld patches and D. Gwynn-Evans, G. Jakowbosky, C. Klak, L. Mucina, K. Oberlander, E. G. H. Oliver, T. Trinder-Smith for plant identifications, Caitlyn von Witt (SANBI) for additional botanical advice, Mike Ford and Iain Fraser for assistance with bird survey work, the Bestlers, S. Foyle, M. Weiner and J. Botha for field accommodation and the landowners who gave us permission to work on their land. Greg Distiller and Colleen Maloney gave statistical advice and Gary Bronner provided small mammal traps and advice. The work was partially funded by the CEPF and NRF. We thank Professor I. Newton and F. Sergio for perceptive comments on earlier drafts of this paper.







