



Ca and phosphorus (P) are two essential macro minerals for vertebrates that play an important role in many biological processes, such as muscle contraction, cell differentiation and bone formation(Reference Rigo, Pieltain, Viellevoye, Milano Buonocore, Bracci and Weindling1). Homeostasis of Ca and P is tightly controlled by the absorption of these minerals in the gastrointestinal tract (GIT), their storage and release in bone and excretion via urine by the kidney, under regulation of hormones such as parathyroid hormone (PTH) and 1,25-vitamin D3 (1,25(OH)2D3)(Reference Blaine, Chonchol and Levi2).
Kidney plays a pivotal role in Ca and P homeostasis in pigs. Several studies have indicated that in pigs absorbed Ca can exceed requirement, and the excess of absorbed Ca is excreted in the kidney via urine. For instance, Stein et al.(Reference Stein, Adeola and Cromwell3) reported that feeding pigs with increasing dietary Ca content from 3·3 to 10·4 g/kg linearly increased daily Ca absorption, which was accompanied by a linear increase in daily urinary Ca excretion. Similar results are reported by Gonzalez-Vega et al.(Reference González-Vega, Liu and McCann4) who found a linear increase in both intestinal absorption and urinary excretion of Ca, in pigs fed diets with increasing dietary Ca content from 3·8 to 10·3 g/kg. These results are in line with our recent study(Reference Hu, Hendriks and van Baal5) where increasing dietary Ca content from 2·0 to 9·6 g/kg elevated daily apparent total tract absorption of Ca, accompanied by an increased urinary Ca excretion and a low post-absorptive Ca utilisation (i.e. retained Ca relative to absorbed Ca, 48 % in the high Ca diets combined with a marginal P deficiency).
Contrary to pigs, intestinal Ca absorption in broilers seems to be under tight control and reaches a plateau when Ca requirement is met. For instance, David et al.(Reference David, Abdollahi and Bedford6) reported that broilers fed incremental dietary Ca from 3·84 to 8·84 g/kg initially increased ileal Ca absorption followed by a plateau phase at approximately 3·8 g absorbed Ca per kg diet. Walk et al.(Reference Walk, Wang and Wang7) also observed a plateau phase in ileal absorbed Ca (approximately 3·0 g absorbed Ca per kg diet) in broilers fed with incremental dietary Ca content from 3·4 to 9·4 g/kg. These reports are in line with our previous findings that incremental dietary Ca content increased ileal Ca absorption to reach a plateau which could not be increased by either further increasing the dietary Ca content, supplemental microbial phytase(Reference Hu, van Harn and Hendriks8) or limestone with a finer/coarser particle size(Reference Hu, Bikker and Duijster9).
The contrast in intestinal Ca absorption between pigs and broilers is in line with differences in mRNA expression of Ca-related transporters and claudins in the GIT. In pigs(Reference Hu, van Baal and Hendriks10), we observed that a high compared with low Ca intake (9·6 v. 2·0 g/kg) reduced jejunal mRNA expression of Ca-related transporters (e.g. TRPV6) but increased mRNA expression of Ca pore-forming claudin-2 (CLDN-2). These results indicate that in growing pigs, increasing dietary Ca intake shift the intestinal Ca absorption from transcellular to the paracellular route, and daily Ca absorption could exceed the requirement rather than being regulated at the intestine. The excess of absorbed Ca was excreted in the urine because it could not be retained in the bone due to P deficiency particularly in the absence of phytase, resulting in a lower post-absorptive Ca utilisation. Apparently, renal Ca reabsorption is tightly regulated to maintain Ca homeostasis and supports that the kidney plays a pivotal role in whole body Ca homeostasis in pigs.
In broilers, transcript levels of both Ca-related transporters and claudins in the duodenum are downregulated by incremental dietary Ca intake and upregulated by a low dietary Ca intake. We found that increasing dietary Ca content gradually reduced mRNA expression of calbindin-D28k (CaBP-D28k) and plasma membrane Ca2+-ATPase 1 (PMCA1), the Ca-sensing receptor (CaSR) and tight junction proteins zonula occludens-1 (ZO-1), CLDN-2 and CLDN-12(Reference Hu, van Harn and Hendriks8,Reference Hu, van Baal and Hendriks11) . The down-regulation of mRNA expression of both Ca-related transporters and claudins, in combination with the observed plateau in intestinal Ca absorption in broilers in contrast to the linear increase in pigs, indicates that intestinal Ca absorption is more regulated in broilers than in pigs to maintain Ca homeostasis. If this is true, the GIT may be key in maintaining whole body Ca homeostasis in broilers, while this is not the case in pigs.
We hypothesised that in pigs, kidney plays a major role in whole body Ca homeostasis, whereas it may be less important in broilers. We previously have conducted a series of studies in pigs(Reference Hu, Hendriks and van Baal5,Reference Hu, van Baal and Hendriks10) and broilers(Reference Hu, van Harn and Hendriks8,Reference Hu, Bikker and Duijster9,Reference Hu, van Baal and Hendriks11) , which allowed us to the test the hypothesis via measuring the mRNA expression of major renal transporters and claudins involved in Ca reabsorption and selected transporters in renal P reabsorption. Regulation on mRNA expression was investigated via maximally challenging Ca and P homeostasis and activating homeostasis mechanism in pigs and broilers fed diets with a wide range of dietary Ca content and inclusion of microbial phytase. Animal studies were conducted under different conditions, but all studies challenged Ca and P homeostasis. All studies were conducted following a design with gradually increasing dietary Ca content from below to above minimal requirement at constant digestible P (dP) content at marginal deficiency (excluding phytase contribution). Previously published results of our studies(Reference Hu, Hendriks and van Baal5,Reference Hu, van Harn and Hendriks8) have confirmed that absorbed P intake was indeed below requirement in the phytase-free diets, although overall dP deficiency was slightly greater in the pig study than in the broiler studies (P absorbed relative to minimal P required in pigs approximately 50 v. 80 % in broilers).
Materials and methods
Kidneys were obtained from one study in growing pigs (Experiment (Exp) 1) and two studies in broilers (Exp 2 and 3) that were approved by the Ethical Committee of Wageningen University & Research and conducted in accordance with the Dutch law on animal experiments. The ethics approval, experimental design, dietary composition, animal husbandry, power analysis and sample collection of the three animal feeding studies have been described in detail elsewhere(Reference Hu, Hendriks and van Baal5,Reference Hu, van Harn and Hendriks8–Reference Hu, van Baal and Hendriks11) . A brief explanation of these studies is provided below.
Experiment 1
Experiment 1 is described in detail in Hu et al.(Reference Hu, Hendriks and van Baal5) and involved forty individually housed male growing pigs (Hypor Libra × Maxter, 30·4 ± 1·3 kg) receiving one of four diets with either a low or high Ca content (2·0 v. 9·6 g/kg, respectively) in the presence or absence of microbial phytase (0 v. 500 FTU/kg) in a 2 × 2 factorial arrangement. The study was conducted over time in two runs with five replicate pigs per treatment per run with each run lasting 21 d. Dietary P content in the basal diet was fixed at marginal deficiency for all treatment groups (total P content 4·7 g/kg, dP 1·7 g/kg excluding the contribution of microbial phytase). Limestone (Sibelco, Maastricht, the Netherlands; particle size < 250 µm as reported by the manufacture) was added to the basal diet to realise the intended Ca content. A 5-d balance trial was conducted to determine daily Ca and P absorption, retention and urinary excretion. Details of the balance trial protocol and results for daily Ca and P absorption and excretion were described in our previous publication(Reference Hu, Hendriks and van Baal5). At dissection, the renal cortex of the left kidney was sliced off with a scalpel, collected in cryovials, snap-frozen in liquid N2 and stored at –80°C until gene expression analysis.
Experiment 2
As described in Hu et al.(Reference Hu, van Harn and Hendriks8), in Exp 2, 1800 male 0-d-old Ross 308 broilers were equally divided among seventy-two pens and allocated to one of eight experimental diets containing either one of four Ca levels in the presence (1000 FTU/kg) or absence of microbial phytase in a 4 × 2 factorial arrangement, with nine replicate pens per treatment and twenty-five birds per replicate pen. The animal trial lasted for 23 d including a starter and grower period from day 0–9 and 10–23, respectively. Dietary retainable P (rP) was fixed at 80 % of CVB(12) recommendation in the basal diet (3·2 and 2·5 g/kg, excluding the contribution of microbial phytase in the starter and grower diet, respectively). Limestone (Faunacal®, Wülfrath, Germany; mean particle size of 90 µm as reported by the manufacture) was added to the basal diet to realise Ca/rP of 1·3, 1·8, 2·3 and 2·8. At dissection, one of the kidneys of one bird per pen was sampled, snap-frozen in liquid N2 and stored at –80°C until further analysis.
Experiment 3
Experiment 3 is described in detail in Hu et al.(Reference Hu, Bikker and Duijster9) and involved 384 0-d-old male Ross 308 broilers equally divided over twenty-four pens and allocated to one of six diets containing one of three Ca levels from either fine or coarse limestone in a 3 × 2 factorial arrangement, with four replicate pens per treatment and sixteen birds per replicate pen. The total P content in the basal diet was sufficient (5·5 g/kg, rP 3·2 g/kg) to meet the requirements of broilers. Fine or coarse limestone was added to the basal diet to realise the intended Ca/P of 0·50, 1·00 and 1·75, respectively. The coarse and fine limestone were derived from the same limestone product (Sibelco) via sieving. The analysed geometric mean diameter was 160 v. 1062 µm for the fine and coarse limestone, respectively. Broilers were given a common commercial starter diet (apparent metabolisable energy 12·5 MJ/kg, crude protein 212 g/kg, Ca 8·6 g/kg and total P 5·5 g/kg) from day 0–13, whereafter dietary treatments were applied for 6 d (day 14–20). At dissection (day 20), the left kidney was sampled (three birds per pen), snap-frozen in liquid N2 and stored at −80°C for gene expression analysis.
Gene expression
Determination of mRNA expression of Ca- and P-related transporters and CLDN was performed following the standard protocol in our laboratory. Briefly, deep-frozen kidney tissue was ground in liquid N2, subsampled (50–100 mg) and used to isolate the total RNA using TRIzol (ThermoFisher Scientific, Waltham, MA). RNA was subjected to on-column DNAse treatment to remove possible genomic DNA contamination with the Nucleospin II kit (Macherey Nagel). Quantity and quality of RNA was determined with the NanoDrop 1000 Spectrophotometer (ThermoFisher Scientific) and 2100 Bioanalyzer and RNA 6000 Nano LabChip kit (Agilent Technologies), respectively. Reverse transcription of total RNA (500 ng) was achieved with a Superscript III kit (ThermoFisher Scientific), and mRNA levels were assessed by real-time quantitative PCR (RT-qPCR) amplification on a QuantStudio 5 Real-Time PCR System (ThermoFisher Scientific) using the SensiFAST™ SYBR® low-ROX Kit (Bioline) under the conditions: 95°C for 15 s and 60°C for 30 s for 40 cycles. This was followed by a melting curve analysis ramping from 60 to 95°C with a rate of 0·1°C/s in order to confirm PCR specificity. Absolute quantitative mRNA levels in the three experiments were calculated by establishing a linear calibration curve using 10-fold serial dilutions of cDNA template for corresponding genes. After evaluating the expression stabilities by the M value of GeNorm, expression levels of genes of interest in Exp 1 (in pig kidney) were normalised to the geometric mean of importin 8 (IPO8), eukaryotic elongation factor 2 (EEF2) and beta actin (ACTB), and expression levels in Exp 2 and 3 (in broiler kidney) to the geometric mean of IPO8, EEF2, ACTB, glyceraldehyde 3-phosphate dehydrogenase (GAPDH) and 60S acidic ribosomal protein P0 (RPLP0). The used primer sequences were designed with Primer Express Software (Life Technologies), and where possible recommended primer sets that span an intron were selected (Table 1). Location and function of Ca- and P-related transporters and claudins are extensively studied before and well reviewed by Yelshanskaya et al. (Reference Yelshanskaya, Nadezhdin and Kurnikova13), Hernando et al. (Reference Hernando, Gagnon and Lederer14) and Otani & Furuse(Reference Otani and Furuse15). Details about relevant transporters are described in the discussion section.
Table 1. Gene-specific primers used for the analysis of mRNA levels using real-time quantitative PCR (RT-qPCR)

-, gene not determined; F, forward primer; R, reverse primer; ACTB, beta actin; CaBP-D9k, calbindin D8k; CaBP-D28k, calbindin-D28k; CaSR, Ca sensing receptor; CLDN, claudin; CYP24, vitamin D3 24 hydroxylase; EEF2, eukaryotic elongation factor 2; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; IPO8, importin 8; NaPi-IIa, Na-dependent phosphate transporter IIa; NaPi-IIb, Na-dependent phosphate transporter IIb; NaPi-IIc, Na-dependent phosphate transporter IIc; NCX1, Na-Ca exchanger; PMCA1, plasma membrane Ca-ATPase 1; PiT-1, inorganic phosphate transporter 1; PiT-2, inorganic phosphate transporter 2; RPLP0, 60S acidic ribosomal protein P0; TRPC1, transient receptor potential canonical 1; TRPM7, transient receptor potential cation channel subfamily M member 7; TRVP5, transient receptor potential cation channel subfamily V member 5; TRPV6, transient receptor potential cation channel subfamily V member 6; VDR, vitamin-D3 receptor; XPR1, xenotropic and polytropic retrovirus receptor 1; ZO-1, zonula occludens-1.
Statistical analysis
Pen was the experimental unit in all three studies. Data were submitted to a two-way ANOVA using the MIXED procedure of SAS (version 9.4, SAS Institute). In Exp 1 and 2, dietary Ca content (or Ca/rP in Exp 2), phytase and their interaction were used as fixed effects and block as random effect. In Exp 3, dietary Ca/P, limestone particle size and their interaction were used as fixed effect, block and pen as random effect. Distribution and variance homogeneity of the Studentized residuals were visually checked via graphics plotted by the ODS GRAPHICS procedure. The LSMEANS procedure with a PDIFF option was used to separate means. In Exp 2, linear and quadratic effect of dietary Ca/rP were determined using the CONTRAST procedure. Probability was considered significant at P ≤ 0·05 and a trend for 0·05 < P ≤ 0·1.
Results
Dietary effects in pig kidney (Exp 1)
The effects on the gene expression levels in the renal cortex of pigs of dietary Ca content in the absence or presence of phytase (i.e. Exp 1) are shown in Table 2. Extracellular CaSR mRNA expression was enhanced more (62 v. 21 %, P interaction = 0·039) with high (9·6 g/kg) dietary Ca content in absence than in presence of microbial phytase (500 FTU/kg), compared with low Ca (2·0 g/kg). calbindin-D9k (CaBP-D9k) mRNA expression was enhanced by high dietary Ca intake in the absence of phytase, with phytase inclusion negating this positive effect (P interaction = 0·002). In contrast, expression of both CaBP-D28k mRNA and Na-Ca exchanger (NCX1) mRNA was reduced with high Ca intake and further reduced with phytase, by 77 v. 36 % (P interaction = 0·025) and 57 v. 21 % (P interaction = 0·024), respectively. TRPV5 mRNA was reduced with high dietary Ca intake in the presence of phytase only (69 % reduction, P interaction = 0·026). Compared with TRPV5, a less abundant expression of TRPV6 (six times lower) was found in the pig renal cortex, which was 40 % lower (P < 0·001) with high Ca intake, regardless of phytase inclusion. Expression of the non-selective active cation channel transient receptor potential cation channel subfamily M member 7 (TRPM7) remained unaffected by the dietary treatments.
Table 2. Least square means of mRNA expression level of various calcium- and phosphorus-related transporters, tight junction proteins, vitamin D3 24 hydroxylase and its receptor in the kidney cortex of growing pigs as affected by dietary Ca content and microbial phytase supplementation (Exp. 1) * , † , ‡
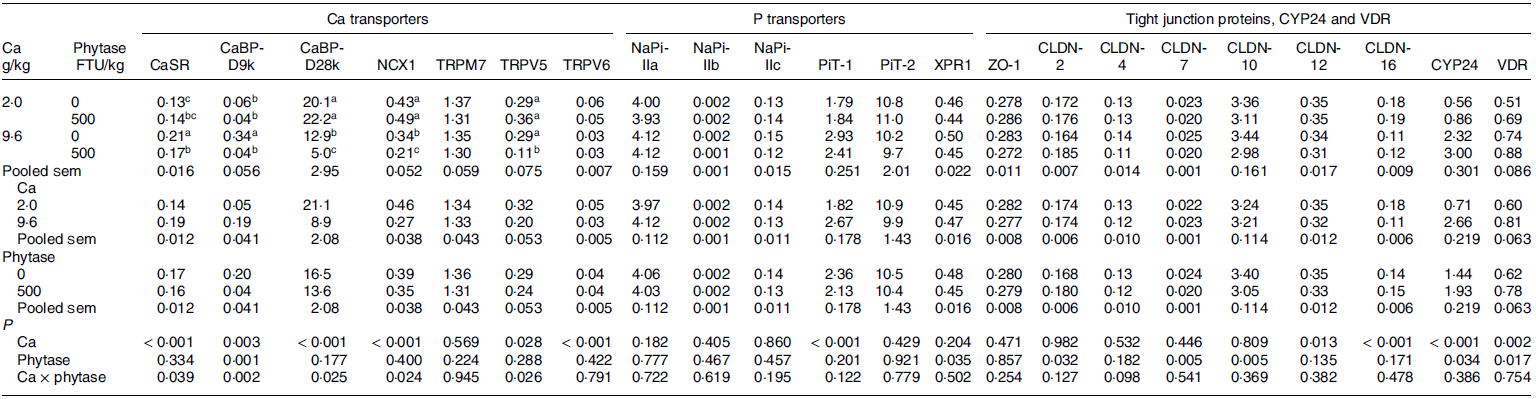
ACTB, beta actin; CaBP-D9k, calbindin-D9k; CaBP-D28k, calbindin-D28k; CaSR, Ca sensing receptor; CLDN, claudin; CYP24, vitamin D3 24 hydroxylase; EEF2, eukaryotic elongation factor 2; IPO8, importin 8; NaPi-IIa, Na-dependent phosphate transporter type IIa; NaPi-IIb, Na-dependent phosphate transporter type IIb; NaPi-IIc, Na-dependent phosphate transporter type IIc; NCX1, Na-Ca exchanger; PiT-1, inorganic phosphate transporter 1; PiT-2, inorganic phosphate transporter 2; TRPV5, transient receptor potential cation channel subfamily V member 6; TRPV6, transient receptor potential cation channel subfamily V member 6; TRPM7, transient receptor potential cation channel subfamily M member 7; VDR, vitamin D3 receptor; XPR1, xenotropic and polytropic retrovirus receptor 1; ZO-1, zonula occludens-1.
a-c Least square means within a column lacking a common superscript differ significantly (P ≤ 0·05).
* Dietary P content was fixed at 4·7 g/kg.
† Data are presented as treatment means, 10 replicate pens per treatment (n 10).
‡ Determined using absolute quantification and normalised by the geometric mean expression of ACTB, EEF2 and IPO8.
Expression of P transporters Na-dependent phosphate transporter IIa (NaPi-IIa), NaPi-IIb, NaPi-IIc, inorganic phosphate transporter 2 (PiT-2) and xenotropic and polytropic retrovirus receptor 1 (XPR1) in porcine renal cortex was not affected by dietary Ca intake or phytase inclusion, apart from PiT-1, which showed a 47 % greater mRNA level in response to high dietary Ca intake (P < 0·001) and for XPR1, which mRNA level was 7 % lower with phytase inclusion (P = 0·035) (Table 2).
Except for tight junction proteins ZO-1 and CLDN-4, mRNA expression of CLDN-2, CLDN-7, CLDN-10, CLDN-12 and CLDN-16 were all affected (P < 0·05) by dietary Ca intake and/or phytase inclusion in pig diets without significant interactions (Table 2). Expression of the Ca-related CLDN, including CLDN-12 (P = 0·013) and −16 (P < 0·001), was reduced by 9 and 38 % with increasing dietary Ca intake, respectively. Expression of Ca pore-forming CLDN-2 was 7 % greater with phytase inclusion (P = 0·032). Phytase inclusion also reduced renal expression of CLDN-7 and CLDN-10 by 17 and 10 %, respectively (P = 0·005). Furthermore, dietary Ca intake and phytase inclusion increased vitamin D3 receptor (VDR) and 24-hydroxylase (CYP24) mRNA expression in the kidney cortex of pigs.
Dietary effects in chicken kidney (Exp 2 and 3)
In the broilers of Exp 2, incremental dietary Ca intake increased renal TRPV6 mRNA (P interaction = 0·002, Table 3) by 67 v. 10 %, and increased CaBP-D28k mRNA (P interaction < 0·001) by 123 v. 36 % in phytase-free and phytase-supplemented diets, respectively. However, CaSR and PMCA1 mRNA level was not significantly affected by the dietary treatments.
Table 3. Least square means of mRNA expression level of calcium- and phosphorus-related transporters, tight junction proteins and vitamin D3 receptor in the kidney of broilers as affected by dietary Ca to retainable P ratio (Ca/rP) and microbial phytase inclusion (Exp. 2) * , † , ‡
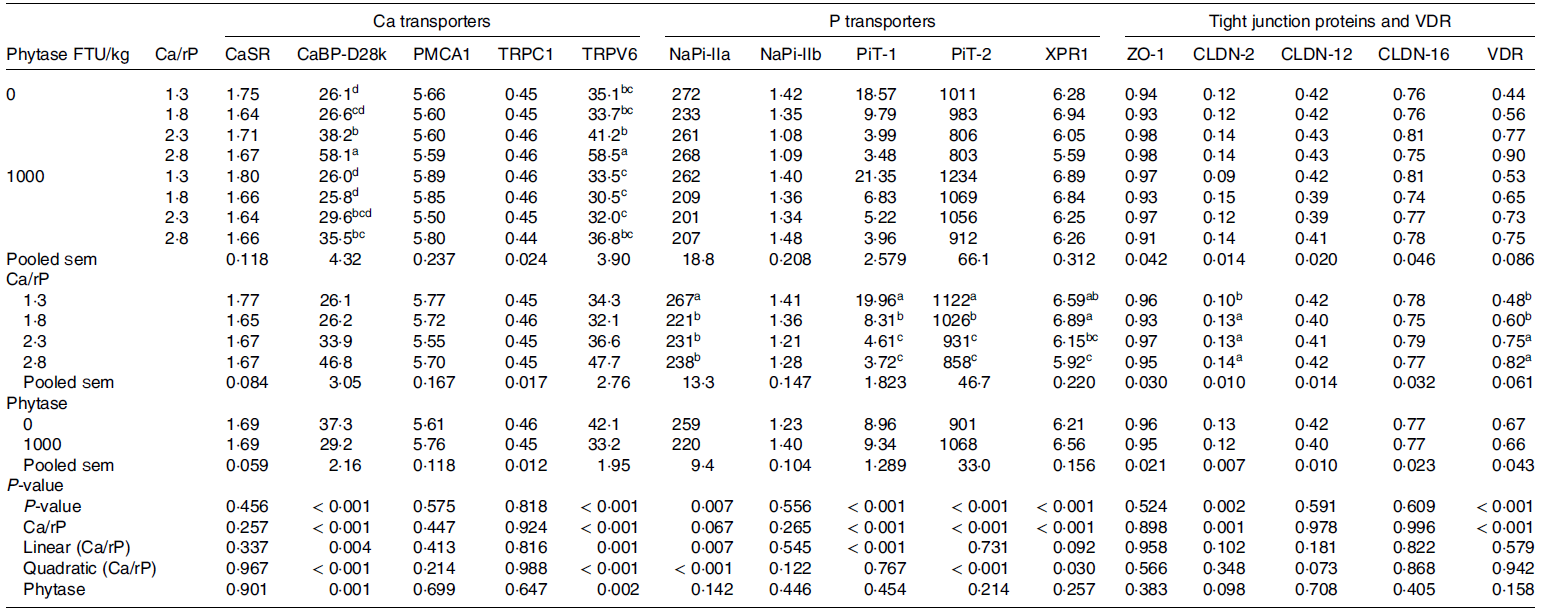
ACTB, beta actin; CaBP-D28k, calbindin-D28k; CaSR, Ca sensing receptor; CLDN, claudin; EEF2, eukaryotic elongation factor 2; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; IPO8, importin 8; NaPi-IIa, Na-dependent phosphate transporter type IIa; NaPi-IIb, Na-dependent phosphate transporter type IIb; PiT-1, inorganic phosphate transporter 1; PiT-2, inorganic phosphate transporter 2; PMCA1, plasma membrane Ca-ATPase 1; RPLP0, 60S acidic ribosomal protein P0; TRPC1, transient receptor potential canonical 1; TRPV6, transient receptor potential cation channel subfamily V member 6; VDR, vitamin D3 receptor; XPR1, xenotropic and polytropic retrovirus receptor 1; ZO-1, zonula occludens-1.
a-d Least square means within a column lacking a common superscript differ significantly (P ≤ 0·05).
* Dietary rP was fixed at 80 % of minimal requirement of rP for broilers (3·2 and 2·5 g/kg excluding the contribution of microbial phytase in the starter and grower period, respectively).
† Data are presented as treatment means, 9 replicate pens per treatment (n 9).
‡ Determined using absolute quantification and normalised by the geometric mean expression of ACTB, EEF2, GAPDH, IPO8 and RPLP0.
No interaction effect between dietary Ca and phytase intake on expression of any of the measured P transporters was observed in the kidney of broilers in Exp 2. Incremental Ca intake reduced NaPi-IIa, PiT-1, PiT-2 and XPR1 mRNA by 11, 81, 24 and 10 %, respectively, in the kidney (P < 0·01). In addition, phytase inclusion reduced (P < 0·001) NaPi-IIa mRNA by 15 % and increased PiT-2 (P < 0·001) and XPR1 (P < 0·030) mRNA by 4 and 19 %, respectively. Renal NaPi-IIb mRNA level was not affected by the dietary treatments.
In Exp 2, CLDN-2 mRNA expression increased linearly with dietary Ca/rP in the kidney of broilers (35 % increase with Ca/rP from 1·3 to 2·8, P = 0·002). CLDN-12 levels tended (P = 0·073) to be lower (4 %) with phytase inclusion. ZO-1 and CLDN-16 mRNA levels were not affected by dietary treatments, while VDR mRNA was linearly increased with incremental dietary Ca/rP (71 % greater with Ca/rP from 1·3 to 2·8, P < 0·001).
In the broiler of Exp 3, incremental Ca/P enhanced renal expression of TRPV6 (P = 0·008) and CaBP-D28k mRNA (P < 0·001) by 34 and 86 %, respectively, and tended (P = 0·073) to enhance renal expression of CaSR by 10 % in broilers, irrespective of limestone particle size (no interaction effects) (Exp 3, Table 4). Limestone particle size had no impact (P > 0·10) on expression of Ca- and P-related transporters, apart from renal CaBP-D28k mRNA expression, which tended (P = 0·053) to increase (16 %) with fine limestone. Dietary treatments did not alter transcript levels of renal PMCA1.
Table 4. Least square means of mRNA expression level of calcium- and phosphorus-related transporters, tight junction proteins and vitamin D3 receptor in the kidney of broilers as affected by dietary Ca/P and limestone particle size (Exp. 3) * , † , ‡
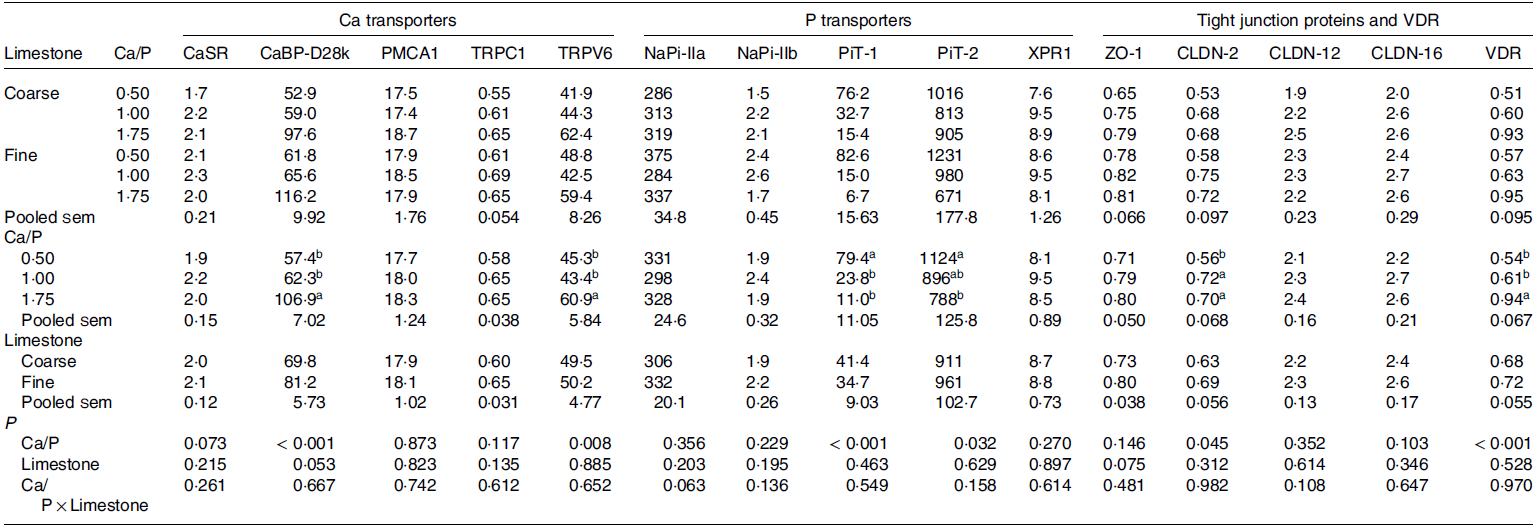
ACTB, beta actin; CaBP-D28k, calbindin-D28k; CaSR, Ca sensing receptor; CLDN, claudin; EEF2, eukaryotic elongation factor 2; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; IPO8, importin 8; NaPi-IIa, Na-dependent phosphate transporter type IIa; NaPi-IIb, Na-dependent phosphate transporter type IIb; PiT-1, inorganic phosphate transporter 1; PiT-2, inorganic phosphate transporter 2; PMCA1, plasma membrane Ca-ATPase 1; RPLP0, 60S acidic ribosomal protein P0; TRPC1, transient receptor potential canonical 1; TRPV6, transient receptor potential cation channel subfamily V member 6; VDR, vitamin D3 receptor; XPR1, xenotropic and polytropic retrovirus receptor 1; ZO-1, zonula occludens-1.
a-b Least square means within a column lacking a common superscript differ significantly (P ≤ 0·05).
* Dietary P was fixed at 5·5 g/kg.
† Data are presented as treatment means, 4 pens per treatment and 3 birds per pen (n 12).
‡ Determined using absolute quantification and normalised by the geometric mean expression of ACTB, EEF2, GAPDH, IPO8 and RPLP0.
Renal NaPi-IIa mRNA expression tended (P interaction = 0·063, Table 4) to be lower (31 %) in diets supplemented with coarse compared with fine limestone at the lowest dietary Ca intake only. Irrespective of limestone particle size (no interaction effect), incremental dietary Ca intake reduced PiT-1 and PiT-2 mRNA by 86 and 30 %, respectively (P < 0·001 and P = 0·032). By contrast, XPR1 mRNA remained unaffected by the dietary treatments.
No interaction effect between limestone particle size and inclusion level on any of the measured tight junction proteins was observed in the kidney of broilers. CLDN-2 mRNA expression was enhanced by 25 % with incremental Ca intake (P = 0·045, Table 4). In addition, ZO-1 mRNA tended (P = 0·075) to be reduced (9 %) in the presence of coarse compared with fine limestone. CLDN-12 and -16 mRNA levels were not affected by dietary treatment. In contrast, VDR mRNA was 74 % greater (P < 0·001) in broilers with a high compared with low Ca intake.
Discussion
Studies conducted to improve dietary P utilisation and reduce P excretion in pigs and broilers indicated a remarkable contrast between pigs and broilers in modulation of daily Ca absorption by dietary Ca content. We previously reported that increasing dietary Ca content linearly increased intestinal Ca absorption and elevated urinary Ca excretion in pigs. In contrast, in broilers incremental dietary Ca intake increased intestinal Ca absorption to a plateau, without further increase thereafter. Intestinal P absorption was measured and confirmed to be below requirement in both pig and broiler studies, although P deficiency was slightly greater in pigs than broilers. Based on the differences in intestinal Ca absorption and mRNA expression of Ca- and P-related transporters and claudins in the GIT between pigs and broilers, we hypothesised that kidney plays a leading role in Ca and P homeostasis in pigs, but not in broilers. Indeed, here we show that the renal expression of Ca- and P-related transporters and CLDN in pigs and chickens respond differently to Ca and microbial phytase intake. In pigs, renal mRNA expression of apical Ca-related transporters (TRPV5 and TRPV6), intracellular located Ca-binding protein (CaBP-D28k), basolateral NCX1 and Ca-related pore-forming claudin-12 (CLDN-12) were all downregulated with incremental dietary Ca intake, as demonstrated in Exp 1 (Table 2). The opposite effect was observed in broilers: incremental dietary Ca intake linearly upregulated the renal mRNA expression of TRPV6, CaBP-D28k and CLDN-2 in both Exp 2 and 3 (Tables 3 and 4).
The mRNA expression of Ca-related transporters in the porcine kidney was largely in line with our previous observations on urinary Ca excretion. We observed that incremental dietary Ca intake substantially increased urinary Ca excretion in pigs(Reference Hu, Hendriks and van Baal5), which is in agreement with the observed down-regulation of renal mRNA expression of Ca-related transporters and CLDN, involved in Ca reabsorption in the kidney, with a higher Ca intake (Table 2). This down-regulation likely reflected a reduction in transcellular and paracellular Ca reabsorption in the porcine kidney and thus a higher urinary Ca excretion.
The mRNA expression of Ca-related renal transporters is generally accepted to be vitamin D3 and/or PTH-dependent. In our companion study(Reference Hu, van Baal and Hendriks10), it was found that in pigs an increase in dietary Ca intake elevated serum Ca concentration, which might have stimulated CaSR signalling in the parathyroid gland, which in turn suppressed the release of PTH into the circulation. A lower PTH concentration might be attributed to the lower serum 1,25(OH)2D3 concentration as observed in pigs fed a high Ca diet by Hu et al.(Reference Hu, van Baal and Hendriks10), probably by catalysing 1,25(OH)2D3 24-hydroxylation by CYP24, as indicated by the upregulated renal expression of CYP24 mRNA by high Ca intake (Table 2). A reduced serum 1,25(OH)2D3 concentration could explain the decreased mRNA expression of TRPV5, TRPV6, CaBP-D28k and NCX1 with high Ca intake, since these four genes share the existence of VDR response elements in their promoter region(Reference Fleet and Feldman16).
The reducing effect of dietary Ca intake on mRNA level of Ca-related transporters in the porcine kidney was greater in phytase-supplemented diets. This interaction could be at least partly attributed to the serum 1,25(OH)2D3 content that was reduced by increasing dietary Ca intake to a greater extent in the presence of microbial phytase(Reference Hu, van Baal and Hendriks10). In the pigs fed phytase-free diets (with P at marginally deficient level), serum P content was rather low, which triggered an elevated serum 1,25(OH)2D3 content irrespective of dietary Ca content(Reference Hu, van Baal and Hendriks10). As a consequence, dietary Ca content showed a limited impact on mRNA expression of Ca-related transporters in diets devoid of microbial phytase, while it reduced Ca transporters mRNA expression in the case where microbial phytase was added to the diet.
Besides Ca transporters, mRNA expression of Ca pore-forming claudins was also regulated by dietary treatment. As shown in Table 2, the reduced mRNA level of TRPV5, TRPV6, CaBP-D28k and NCX1 was accompanied by a lower level of CLDN-12 and −16 mRNA expression in pigs with a higher Ca intake. A study in mice indicated that CLDN-12 formed Ca-selective pores and its mRNA and protein expression was upregulated by exposure to 1,25(OH)2D3 (Reference Fujita, Sugimoto and Inatomi17). In murine kidney, CLDN-16 is mainly expressed at the distal convoluted tubule where it interacts with TRPV5 to mediate Ca reabsorption(Reference Hou, Rajagopal and Yu18,Reference Hou, Renigunta and Nie19) . Thus, besides the well-documented and tightly controlled transcellular route, paracellular Ca permeation in the porcine kidney was also regulated to maintain whole body Ca homeostasis.
In contrast to pigs, incremental Ca intake enhanced renal mRNA expression of Ca-related transporters in broilers (Tables 3 and 4; online Supplementary Fig. 1). We previously observed that a higher Ca intake enhanced serum Ca concentration in broilers(Reference Hu, van Harn and Hendriks8,Reference Hu, Bikker and Duijster9) . Thus, unlike mammals, mRNA expression of Ca transporters in the kidney in this species was positively correlated with serum Ca concentration. The structure of the avian kidney is rather different from that of mammals, as 70–90 % of the chicken’s nephron is reptilian type and does not contain a loop of Henle(Reference Goldstein and Braun20). Kidney structure might play a role in the apparently different regulation of Ca reabsorption in renal tubules of chickens.
Our observations are in line with several in vivo studies in chickens. For instance, Bar et al. (Reference Bar, Shani and Fullmer21) reported that low dietary P intake or high dietary Ca intake increased serum Ca concentration in chickens, which was associated with a higher renal mRNA expression of CaBP-D28k. Similar modulation of renal CaBP-D28k protein expression was observed in another in vivo chicken study from these researchers(Reference Rosenberg, Hurwitz and Bar22) which showed that a lower dietary Ca intake reduced serum Ca concentration, accompanied by an elevated serum 1,25(OH)2D3 level and reduced CaBP-D28k protein expression in the kidney. A similar effect on these three parameters was obtained with low dietary P content(Reference Rosenberg, Hurwitz and Bar22). It is noteworthy that both studies(Reference Bar, Shani and Fullmer21,Reference Rosenberg, Hurwitz and Bar22) also demonstrated that CaBP-D28k protein expression in the GIT was reduced upon high Ca intake in chickens. Those findings align with our previous studies(Reference Hu, van Harn and Hendriks8,Reference Hu, Bikker and Duijster9) that high Ca intake reduced mRNA expression of Ca-related transporters and CLDN in the broilers’ GIT and with the present study that high Ca intake upregulated these genes in broilers’ kidney (Tables 3 and 4).
The up-regulation of Ca-related transporters and CLDN in the kidney of broilers in response to higher Ca intake demonstrates that whole body Ca homeostasis in poultry is differently regulated compared with mammals. An earlier study in chickens indicated that vitamin D3 treatment enhanced CaBP-D28k mRNA expression 8-fold more in the duodenal mucosa than kidney(Reference Christakos and Norman23). Apparently, CaBP-D28k mRNA expression in the GIT was more sensitive to changes in serum 1,25(OH)2D3 than in the kidney, indicating that the GIT might play a primary role in whole body Ca homeostasis in chickens. Indeed, duodenal mRNA expression of CaSR, CaBP-D28k and basolateral Ca2+-ATPase (PMCA1) downregulated by serum Ca concentration in broilers(Reference Hu, van Harn and Hendriks8,Reference Hu, van Baal and Hendriks11) , presumably to maintain serum Ca concentration within a normal physiological range.
Renal expression of P-related transporters appears to be predominantly regulated at a post-transcriptional level in pigs(Reference Saddoris, Fleet and Radcliffe24). We found that urinary P excretion was substantially reduced with incremental Ca intake in pigs(Reference Hu, Hendriks and van Baal5), while renal mRNA expression of P-related transporters was largely unaffected by both dietary Ca and phytase inclusion (Table 2). This discrepancy between urinary P excretion and mRNA expression levels of renal P transporters might indicate the involvement of post-transcription regulation mechanisms, that is, trafficking of NaPi-IIa to and from the apical membrane via its PDZ binding motif(Reference Biber, Hernando and Forster25,Reference Levi and Gratton26) . However, we did not further investigate the post-transcription regulation of NaPi-IIa since it was beyond the scope of this study. It is noteworthy that a pore-forming CLDN for P has not been identified yet, hence renal P reabsorption via the paracellular pathway in response to dietary Ca and phytase intake remains unknown in the kidney of pigs.
In contrast to pigs, incremental dietary Ca intake reduced the mRNA expression of P transporters (NaPi-IIa, PiT-1, PiT-2 and XPR1) in the kidney of broilers. We previously found that dietary Ca intake decreased serum P concentration(Reference Hu, van Harn and Hendriks8,Reference Hu, Bikker and Duijster9) . Thus, mRNA expression of P-related transporters in the kidney of broilers appeared to be positively correlated to serum P concentration. Our results are in line with those of Ren et al.(Reference Ren, Yan and Hu27), who indicated that laying hens fed a reduced P diet had a lower serum P concentration, which was accompanied by a lower NaPi-IIa protein expression in the kidney. Renal NaPi-IIa expression is under tight control of 1,25(OH)2D3 since its promoter contains a VDR-responsive element(Reference Proszkowiec-Weglarz and Angel28). It is conceivable that broilers fed a higher Ca diet had greater serum Ca content(Reference Hu, van Harn and Hendriks8,Reference Hu, Bikker and Duijster9) , which might activate CaSR on the parathyroid gland and trigger a lower secretion of PTH, thereby reducing serum 1,25(OH)2D3 concentration(Reference Bergwitz and Juppner29) and lowering the expression of P-related transporters in broilers.
We also observed a reducing impact of phytase inclusion on renal CLDN-7 and -10 mRNA expression in pigs. A study using the LLC-PK1 cell line indicated that CLDN-7 formed a barrier for chloride but a paracellular channel for Na(Reference Alexandre, Lu and Chen30). In contrast, a murine kidney collecting duct cell line indicated that genetic deletion of CLDN-7 decreased permeability for both Na and chloride(Reference Fan, Tatum and Hoggard31). A similar debate exists for CLDN-10, as it may confer a channel for Na or chloride in the LLC-PK1 cell line depending on the spliced variants(Reference Van Itallie, Rogan and Yu32). While our results suggest that the paracellular permeability to traverse certain minerals, possibly Na, across the renal epithelial layer is affected by phytase inclusion, further studies are needed to elucidate the permeability functions of CLDN-7 and -10 in the porcine kidney.
In conclusion, the mRNA expression of Ca-related transporters and Ca pore-forming CLDN are downregulated in the kidney of pigs, but upregulated in broilers, in response to incremental dietary Ca and/or microbial phytase content. Unlike the central role of the porcine kidneys in maintaining whole body Ca homeostasis, in chickens the intestine appears to be far more important. As such, insights in Ca metabolism obtained from studies in pigs should not be directly extrapolated to poultry and vice versa.
Acknowledgement
The authors thank that Mr L. Kruijt assisted with RT-qPCR analysis, and Y. X. H. received a scholarship form the Chinese Scholarship Council (CSC) to study at Wageningen University & Research.
This research was conducted within the framework of the public private partnership ‘Feed4Foodure’ (AF16123), which is supported by the ‘Vereniging Diervoederonderzoek Nederland (VDN)’ and the Ministry of Agriculture, Nature and Food Quality (LNV, project number BO-31.03-005).
P. B., M. M. v. K., J. v. B., W. H. H. and Y. X. H. designed and conducted the research; Y. X. H. analysed the samples and data; J. v. B. designed and validated the RT-qPCR primers. Y. X. H., J. v. B., P. B. and W. H. H. interpreted the data and wrote the manuscript.
All authors with the exception of the late M. M. v. K. have read and approved the final manuscript and declare to have no conflict of interest.
Supplementary material
For supplementary material/s referred to in this article, please visit https://doi.org/10.1017/S0007114524002332







 Open access
Open access