


INTRODUCTION
Leptospirosis has been reported in more than 150 mammalian species [Reference Ko, Goarant and Picardeau1]. The disease is maintained by the persistent colonization of the proximal renal tubules of carrier animals. A reservoir animal can remain asymptomatic and shed infectious Leptospira in its urine for a transient period of time or for its entire lifetime [Reference Thiermann2, Reference Leonard3]. Infection most frequently results from direct contact with infectious urine, although genital transmission [Reference Lilenbaum4–Reference Harrisson and Fitzgerald7] and contamination by consumption of infected prey [Reference Reilly, Hanson and Ferris8] have also been described. Direct infection of newborns through breastfeeding has also been reported [Reference Bolin9]. The incidence of different Leptospira strains in human populations strongly depends on the reservoir hosts present locally and the strains they carry as well as the amount of contact between humans and fauna, ecological conditions, and cultural and agronomical practices [Reference Bharti10].
Reunion Island is a French tropical overseas region located in the Indian Ocean. With an incidence rate of 15·13 cases/100 000 inhabitants in 2010 (33 times higher than the rate observed in metropolitan France) [11], leptospirosis is an understudied major public health problem on this developed island. The only native mammals on the island are bats; however, paralleling human colonization, settlers have progressively introduced a number of farm animals from Europe (cattle, Bos primigenius; goat, Capra aegagrus; pig, Sus scrofa; sheep, Ovis aeries). Horses (Equus ferus); game species, such as Rusa deer (Rusa timorensis) or the tenrec (Tenrec ecaudatus); and domestic species, such as dogs (Canis lupus) and cats (Felis sylvestris) have also been intentionally introduced to the island. The ship rat (Rattus rattus), the Norway rat (Rattus norvegicus), the mouse (Mus musculus), and the shrew (Suncus murinus) have been accidentally introduced by ships and boats.
The last available data for animal leptospirosis on Reunion Island date back to the 1980s [Reference Moutou12–Reference Mollaret, Mailloux and Debarbat14], and these data consist of seroepidemiological surveys showing that 40% of stray dogs [Reference Moutou12, Reference Mollaret, Mailloux and Debarbat14], up to 32% of cattle [Reference Moutou12, Reference Debarbat, Mollaret and Mailloux13], 5% of pigs [Reference Moutou12], and up to 71% of horses [Reference Moutou12, Reference Debarbat, Mollaret and Mailloux13] were seropositive for leptospirosis. Surprisingly, the rodent reservoir had never been investigated.
The local preventive measures that have been employed against leptospirosis have only focused on health education programmes and rodent control but other potential reservoirs remain neglected. Obtaining information on the prevalence of leptospiral infection among the various animal populations and identification of the animal species that are the predominant carriers is important to inform prevention and control programmes. Thus one of the aims of this work was to conduct a field-to-laboratory survey in order to update data on animal leptospirosis on Reunion Island. To achieve this goal, we first conducted a transversal survey to estimate the seroprevalence of leptospirosis and evaluate the circulating serogroups in nine animal species. Next, we used quantitative polymerase chain reaction (qPCR) to study the prevalence of renal infection at the time of sampling in 12 animal species. To our knowledge, this is the first report of quantitative results of leptospires in the kidney tissues of naturally infected animals. Data detailing the leptospiral status of the animal species living close to humans represent the first step in planning effective control measures to protect humans, and therefore, these data must be regularly updated to guide the development of preventive measures to counter potential epidemiological changes.
METHODS
Field methods
Animal captures were conducted between 12 February 2009 and 20 August 2009 on Reunion Island. Black rats (R. rattus), Norway rats (R. norvegicus), mice (M. musculus), and shrews (S. murinus) were trapped in the field using baited live-traps laid out overnight (Manufrance traps for rats and INRA traps for mice and shrews; BTTm, France). Tenrecs (T. ecaudatus) were hand-captured alive by local hunters during the official hunting season using traditional hunting methods (prefectural decree no. 1268, 30 May 2008). Small mammals (rats, shrews, mice, tenrecs) were euthanized by injection of pentobarbital, following the recommended procedure [15] and blood was collected by cardiac puncture. Stray dogs and cats were captured by employees of the local animal rescue using a special lariat. These dogs and cats were euthanized by the veterinary services using an intravenous pentobarbital overdose (Doléthal®, Vétoquinol, France), and blood was sampled from the cephalic vein. Cattle, goats, deer (R. timorensis), and pigs being prepared for human consumption were subjected to blood sampling from the carotid artery immediately after bleeding and kidney samples were collected at the evisceration and inspection area of the slaughterhouse. For all animals, blood sampling was conducted immediately after death, and the kidneys were removed aseptically after the renal capsule was removed.
Free-tailed bats (Mormopterus francoismoutoui) were trapped at nightfall using a black nylon Japanese mist net (Bonardi, 110D, mesh of 16 mm, 12 m long, 2·4 m high, five pockets), and released immediately after urinary sampling. Blood sampling was not conducted on the bats because the quantity of blood required from each individual bat for the microscopic agglutination test (MAT) would be lethal. Bat urine was collected directly after capture at the urethral opening using a sterile rayon swab.
Just after sampling, the blood was centrifuged and the collected serum was stored at −80 °C until analysed. The urine samples were neutralized with an approximately equivalent volume of phosphate-buffered saline (PBS) [Reference Lucchesi16] and frozen at −80 °C. Rayon swabs with urine samples were placed into transport media and vortexed before being frozen at −80 °C.
A total of 574 animals belonging to 12 species were sampled. Mice, shrews and bats were not sampled for blood. Ten individual urine samples were collected in a small colony of free-tailed bats located in a house. All but one of the pigs slaughtered during our survey were 5- to 6-year-old reproductive sows. None of the animals that we sampled were vaccinated against leptospirosis. We analysed 462 sera samples by MAT, and 546 kidney and 10 bat urine samples were screened for the presence of leptospiral DNA by qPCR.
MAT
Sera were tested for the presence of anti-leptospiral antibodies using the MAT following standard procedures [Reference Faine17]. Seventeen reference strains belonging to 15 serogroups were used as antigens in the MAT (Table 1). We considered 1:100 as the cut-off value for positivity. The serogroup with the highest titre was considered to be the presumptive single infecting serogroup and serum with this result was classified as being infected with the corresponding single Leptospira serogroup. In contrast, if two or more serogroups induced equally high titres, we lableled these animal samples as having multiple successive infections or co-infections.
Table 1. Strains used as capture antigens in the microscopic agglutination test

* Strains provided by the Pasteur Institute, Paris were isolated from a patient in Mayotte.
Positive control and standard DNA construction
We cloned the 242-bp fragment of the lipL32 gene of L. interrogans serovar Copenhageni strain Wijinberg into the pGEM®-T Easy Vector (Promega, USA) following the manufacturer's recommendations and using the primers LipL32-45F/LipL32-286R (Table 2) [Reference Stoddard18]. A twofold dilution series of the plasmid DNA was prepared to 10−12 to test the analytical sensitivity (determined by the number of plasmid copies that was detected ten times out of ten repetitions) of the real-time qPCR and to construct a standard curve for DNA quantification of positive samples. DNA content was quantified using the QuantIt™ PicoGreen® kit (Invitrogen, France) using the LightCycler 2.0 System (Roche Diagnostics, France) according to the manufacturer's recommendations.
Table 2. Primers and probes used for qPCR assays
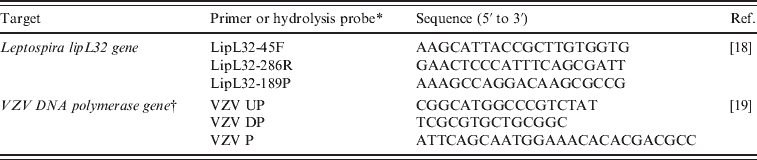
qPCR, Quantitative polymerase chain reaction.
* All probes were labelled with FAM at the 3′ end and with TAMRA at the 5′ end.
† Gene bank accession no. AB059828-31 and X04370.
Internal control (IC) construction
To control for the DNA extraction step and to detect the presence of PCR inhibitors in the biological samples, an IC was chosen based on the following criteria: the IC should not be present in the DNA of the different animal species studied, the IC DNA sequence should not be present in any pathogen that could infect the animals, and the IC DNA sequence should not share similarity with a Leptospira DNA sequence. Thus, the IC was constructed using the 63-bp sequence of the DNA polymerase gene of varicella-zoster virus (VZV) isolated from a human VZV-positive clinical sample and cloned into the pGEM®-T Easy Vector (Promega) using the primers VZV UP/VZV DP (Table 2) [Reference Weidmann, Meyer-König and Hufert19]. To identify the optimal concentration for use in the qPCR, a tenfold serial dilution of the IC, ranging from undiluted to a 10-12-fold dilution, were tested by qPCR. The optimal IC concentration was established on the criterion that a reliable IC amplicon was always detected in the samples.
Total DNA extraction
DNA was extracted from 20–25 mg of cortex kidney tissue using the Dneasy® Blood and Tissue kit (Qiagen, France) according to the manufacturer's instructions. When adding the proteinase K, we also added 10 μl of the IC plasmid at the optimal IC concentration. Thus, the IC was co-purified with the sample DNA and was detected as a positive control for the extraction process.
Gene amplification by qPCR
The sensitivity of our qPCR assay was 7400 copies/ml. Because one leptospire contains an average of five genome equivalents [Reference Bourhy20], our analytical sensitivity was calculated to be 1-2 bacteria/μl (2·5–5 bacteria/reaction). All real-time PCR reactions were performed using the LightCycler® 480 (Roche Diagnostics). The detection of Leptospira DNA was performed using a Taqman probe targeting the lipL32 gene as previously described by Stoddard et al. [Reference Stoddard18] (Table 2). The absence of PCR inhibition in each sample was assessed in a separate qPCR assay targeting the IC using primers VZV UP/VZV DP and detected with the hydrolysis probe VZV P (Table 2) [Reference Weidmann, Meyer-König and Hufert19]. For each reaction, the lipL32-containing plasmid was used as a positive control and two negative controls (sterilized water) were included to detect the presence of contaminating DNA. Analyses of samples were performed in triplicate, and to avoid intra-PCR contamination, we repeated the qPCR within different runs for each sample. For each sample, interpretation of the qPCR results could be done when the amplification of the IC was positive, the two negative controls within the run were negative, and the cycle threshold (Ct) value of the positive control was between 24 and 27 cycles. A negative result was assigned in the cases where no amplification of the lipL32 gene occurred (i.e. the Ct value was greater than 40 cycles). A positive result was assigned to a sample when the three qPCRs targeting the lipL32 gene were positive.
Statistical methods
All variables were reported as percentage (or mean) ± standard deviation (s.d.). We used a χ2 test to compare seroprevalences and prevalences of renal carriage between species; P values were adjusted using Holm's adjustment method and were considered significant when <0·05. Comparisons of the mean renal concentrations of leptospires between species were not biologically relevant due to wide inter-individual variations within each species. Data analysis was performed using R statistical software [21].
Sampling authorization and ethics statement
Rats, mice and shrews are introduced invasive mammals on Reunion Island, thus no particular authorization was required for their capture and study. Euthanasia/slaughtering of animals was conducted ethically by following the recommended procedures of the Parliament and the Council of the European Union [15]. This research adhered to the French legal requirements articles R.214-93 and R.214-99 to R.214-102 of the French Rural Code and Order of 19 April 1988, giving the authorization to experiment on living animals. For stray carnivores, French law no. 99-5 (6 January 1999) concerning ‘dangerous and stray animals and protection of animals’ was followed. The treatment of livestock followed European Regulation no. 1099/2009 (24 September 2009) and the French Order (12 December 1997) defining ‘the ethical procedures concerning welfare, protection, immobilization, stunning, and euthanasia of livestock animals at the slaughterhouse’. Our study received the approval of the Health Veterinary Inspector, the Director of the Veterinary Services of Reunion Island, the Director of the animal rescue, and the Director of the slaughterhouse of Saint-Pierre. Capture and sampling of the protected bat M. francoismoutoui required an official authorization that was issued on 10 March 2009 from the Direction Régionale de l'Environnement (DIREN) of Reunion Island. Sampling of the game species, T. ecaudatus, required an official authorization from the Direction de l'Agriculture et de la Forêt (DAF) and from the Hunting Federation of Reunion Island, that we obtained on 13 January 2009 (no. BD/BF/012).
RESULTS
The results of trapping showed that the three species of rodents are sympatric, but the black rat was the most common rodent caught. Black rats accounted for 85·1% of all rodents sampled, while mice accounted for 9·9%, and the Norway rat for 4·9%.
All results are summarized in Supplementary Table S1.
Serological results
We found that the seroprevalence varied considerably depending on the species (Fig. 1). The seroprevalence (%±s.d.) was highest in rats (Norway and black) with 79·5 ± 9·3% (58/73) of seropositive animals; followed by Rusa deer (61·7 ± 16·3%, 21/34); goats (60·0 ± 12·4%, 36/60); pigs (47·2 ± 10·4%, 43/89); stray dogs (46·0 ± 13·8%, 23/50); cattle (34·0 ± 9·9%, 30/88); stray cats (26·6 ± 15·8%, 8/30); and tenrecs (13·2 ± 10·7%, 5/38).

Fig. 1. Seroprevalence of leptospirosis (and standard deviation). Seroprevalence was assessed by MAT on eight animal species from Reunion Island.
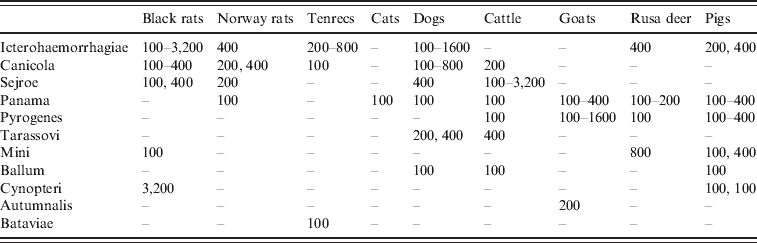
The serological results are summarized by species in Tables 3 and 4. Serogroup Icterohaemorrhagiae accounted for 39·7% of all seropositive reactions in Norway and black rats (titres 100–3200), and serogroup Canicola was the second highest serogroup in rats, representing 15·5% of the seropositive reactions (titres 100–400). In tenrecs, the main circulating serogroup was Icterohaemorrhagiae (titres 200–800) but serogroups Canicola and Bataviae were also reported. Stray cats were more frequently seropositive for several serogroups (87·5%) and all antibody titres were low (⩽400). Canicola was the most prevalent serogroup in stray dogs representing 43·5% of the seropositive animals (titres 100–800), while Icterohaemorrhagiae accounted for 21·7% (titres 100–1600). In cattle, 56·6% of positive animals had antibodies to serogroup Sejroe, and titres reached 3200. Panama was the main serogroup found in goats, Rusa deer, and pigs accounting for 69·4%, 38·1%, and 37·2% of the seropositive reactions for these animals, respectively. Pyrogenes was the second highest serogroup found in these species, representing 16·6%, 23·8%, and 25·6% of the seropositive results for goats, Rusa deer, and pigs, respectively. Antibody titres to Leptospira were low in goats, Rusa deer, and pigs (⩽400) except for two goats that had titres of 800 and 1600 to serogroup Pyrogenes.
Table 3. Results of microscopic agglutination test conducted on animal sera from Reunion Island

* A serum showing agglutinations for more than one serogroup with no serogroup presenting a highest titre.
Table 4. Titres observed by microscopic agglutination test by species and serogroup. Minimum and maximum titres are presented. If one or two animals were seropositive to a serogroup, the titres are presented. Seropositive reactions to more than one serogroup are not included
The seroprevalence was significantly higher in rats than in tenrecs (P = 3·1 × 10−6), cats (P = 0·008), and cattle (P = 0·0003). Seroprevalence was also significantly higher in goats (P = 0·041) than in cattle, whereas seroprevalence in tenrecs was significantly lower than seroprevalence in goats (P = 0·0003), deer (P = 0·018), and pigs (P = 0·011) (pairwise P values are shown in Supplementary Table S2).
Renal carriage
Of the 541 kidney samples, 78 contained qPCR inhibitors; thus, only 463 kidneys samples could be analysed for the presence of Leptospira. Renal carriage (% ±s.d.) of Leptospira was most prevalent in mice with 84·6 ± 19·6% (11/13) positive animals, followed by rats (65·9 ± 10·3%, 54/82), shrews (31·2 ± 13·1%, 15/48), stray dogs (29·2 ± 19·3%, 7/24), stray cats (28·6 ± 18·2%, 6/21), goats (26·5 ± 12·4%, 13/49), Rusa deer (18·8 ± 13·5%, 6/32), cattle (18·2 ± 8·7%, 14/77), and pigs (15·6 ± 7·8%, 13/83). No leptospiral DNA was detected in tenrec kidneys (0·0 ± 5·0%, 0/18) (Fig. 2). The carriage rate between rats and mice was not significantly different, but both of these rodents showed significantly higher prevalence of renal carriage than the other species (pairwise P values are shown in Supplementary Table S3). There was no other significant difference in the prevalence of renal carriage between species.

Fig. 2. Prevalence of renal carriage (and standard deviation). Presence of leptospires in kidney tissue was assessed by qPCR targeting the lipL32 gene on ten animal species from Reunion Island.
The mean concentrations of leptospires in the kidneys for each species are presented in Table 5.
Table 5. Mean leptospiral load in kidney tissue by species. The concentration of leptospires in kidney tissue was assessed by qPCR targeting lipL32 gene in each carrier animal

qPCR, Quantitative polymerase chain reaction; s.d., standard deviation.
Urinary shedding in bats
Eight out of ten individual samples of bat urine contained qPCR inhibitors but both of the amplifiable urine samples were positive for the presence of Leptospira DNA.
DISCUSSION
Although it is often neglected, descriptive epidemiology is the first step required for in-depth studies of leptospirosis (i.e. molecular epidemiology and phylogenetic classification). The aim of this study was to update the data on animal leptospirosis on Reunion Island. The epidemiological investigation of 12 out of the 16 mammalian species living on Reunion Island contributes to global knowledge of the disease at the international level (hare, horse, sheep, and the protected insectivorous bat Taphozous mauritianus were not investigated due to difficulties in the sampling protocol and/or in gaining the appropriate authorizations).
Our results indicate that Leptospira infection is unexpectedly common in all of the investigated mammals on Reunion Island (Figs 1 and 2, Supplementary Table S1). In this study, PCR-based detection of leptospiral DNA in kidney or urine samples was used as the definitive proof of carrier or infectious status [Reference Tulsiani22]. Because leptospires only transiently colonize the kidneys in non-reservoir species [Reference Hartskeerl and Terpstra23], the detection of leptospires in the various kidney samples was expected to be minimal if the species was not a reservoir (~1%). Interestingly, our results indicate that all of the mammals on Reunion Island, with the exception of tenrecs, are potential renal carriers of Leptospira (Fig. 2). The absence of leptospiral DNA in the kidney samples of tenrecs and the low seroprevalence (compared to other species) may indicate that tenrecs are either less exposed than other species or that they may be susceptible to infection but are most likely not chronic reservoirs of Leptospira. In contrast, our results showed that 31·2% of shrews are carriers of pathogenic Leptospira, which is lower than the prevalence rate recently reported in Madagascar (43·5%, n = 26) [Reference Rahelinirina24]. Nevertheless, our study corroborates other findings and provides further support that this insectivore may act as a maintenance host for Leptospira [Reference Rahelinirina24, Reference Kundin25].
Furthermore, we demonstrated that stray carnivores are frequently renal carriers of Leptospira. Although dogs are known to be a potential zoonotic reservoir host of leptospirosis [Reference Jimenez-Coello26], the possibility of persistent renal infection in cats by Leptospira is contentious [Reference Everard27–Reference Felt29]. To our knowledge, leptospires have rarely been found in cat kidneys: Leptospira from serogroup Canicola was isolated from a cat on the island of Trinidad (West Indies) [Reference Everard27], while in Spain, Millán et al. [Reference Millan30] reported a prevalence of renal carriage in 20% (5/25) of the roaming cats investigated. Additionally, a recent study described three clinical cases of leptospirosis in naturally infected cats [Reference Arbour31]. Our study adds to these data and unambiguously indicates that cats could be renal carriers of Leptospira and therefore possibly be a source of infection to humans. We hypothesize that stray dogs and cats may be exposed to Leptospira via consumption of small infected mammals (rodents and shrews) [Reference Reilly, Hanson and Ferris8] which are known to be Leptospira carriers, or by roaming in contaminated environments [Reference Jorge32].
The significant prevalence of renal carriage in livestock (Fig. 2) probably reflects the fact that these animals are chronically infected, which raises a significant concern due to the zoonotic nature of the disease. Widespread circulation of leptospirosis in livestock may also have financial implications as infection may result in a decrease in milk quality and quantity, an increase in abortions, higher culling rates, more stillbirths, and death [Reference Langoni33–Reference Ramos, Souza and Lilenbaum35].
The carriage rate reported in rats in our study (65·9%) was higher than the prevalence rate generally found on other tropical islands such as Madagascar where 40% of renal carriers were reported in rats (n = 190) [Reference Rahelinirina24], or Hawaii, where 60·2% of renal carriers were found in the R. norvegicus population (n = 510) [Reference Minette36], or even New Caledonia, where 26·7% of rats were carriers of Leptospira [Reference Perez37]. Nevertheless, our prevalence rate was close to that reported in rats in Metro Manila in the Philippines (63%, n = 56) [Reference Villanueva38] and lower than the prevalence rate described in Norway rats in Brazil (80·3%, n = 142) [Reference de Faria39]. We reported a prevalence of renal carriage of 84·6% in mice, which was in accord with other studies conducted in Hawaii (79·5%, n = 31/39) [Reference Minette36], and the Azores (85·4%, n = 41) [Reference Collares-Pereira40]. The differences observed in carriage prevalence could be due to study design and the detection strategies employed but may also reflect different dynamics in the different ecosystems.
In consideration of the fact that the bat, M. francoismoutoui, is a protected species, we chose to conduct non-invasive sampling on this mammal. The number of non-amplifiable bat urine samples (8/10) may indicate that the sampling technique used for bats was not optimal and that contamination by PCR inhibitors occurred. Nevertheless, two bats were shown to be urinary shedders of Leptospira. The role that bats play in the transmission of Leptospira is not fully understood. Most of the studies focused on domestic animals because of their close association with humans; however, because of their abundance, and close contact with both domestic and wild animals and humans, bats may be involved in the maintenance and transmission of leptospires on Reunion Island, as reported elsewhere [Reference Tulsiani22, Reference Bunnel41]. Studies in Peru reported that bats can be reservoirs of leptospirosis [Reference Bunnel41, Reference Matthias42] and the survey conducted by Matthias et al. [Reference Matthias42] described a prevalence of renal carriage of 3·4% (n = 589) in insectivorous bats. Furthermore, on Reunion Island, bat guano is used for agricultural purposes and collecting the guano under a roosting site could expose people to Leptospira infection.
Seroprevalence was high in all the species investigated (Fig. 1) suggesting that contacts with Leptospira are frequent. We report a higher seroprevalence of Leptospira infection in rats than that commonly reported [Reference Collares-Pereira40, Reference Desvars, Cardinale and Michault43], except in the Philippines, where 92% of rats showed antibodies to Leptospira [Reference Villanueva38].
In this study, we demonstrated that the seroepidemiological results in dogs were similar to those found in 1979 (seroprevalence 41% in 1979 [Reference Moutou12] vs. 46·0% in our study) and that Canicola has remained the main circulating serogroup in stray dogs (69% of those seropositive in 1979 [Reference Moutou12] vs. 43·5%) followed by Icterohaemorrhagiae (26% [Reference Moutou12] vs. 21·7%). Serogroups Sejroe, Mini, and Cynopteri in rats, as well as Sejroe, Panama, Tarassovi, and Ballum in dogs (Tables 3 and 4) showed low seroprevalence and/or low titres, suggesting that contact with these serogroups was rare or was detected in our study as the result of non-specific co-agglutination. Between 1998 and 2009, serogroup Icterohaemorrhagiae and Canicola were reported to represent 59·3% and 17·5%, respectively, of the human cases diagnosed on Reunion Island [11]. Our results suggest that rats and dogs are the main source of human infection and corroborate recent findings reporting that rats may also act as a reservoir for serogroup Canicola [Reference Rahelinirina24, Reference Doungchawee44]. The seroprevalence in cattle was similar to that reported by two studies conducted on Reunion Island in 1979 (29% [Reference Debarbat, Mollaret and Mailloux13] to 32% [Reference Moutou12]), and serogroup Sejroe has remained the major serogroup circulating in this species. Eight out of 17 cattle seropositive to serogroup Sejroe had antibody titres ⩾800, probably indicating a recent infection [Reference Adesiyun45]. Between 1998 and 2009, Sejroe was involved in 5·4% of human infections [11], suggesting that cattle are seldom involved in human contamination on Reunion Island. Panama was the most prevalent serogroup circulating in Rusa deer, goats, and pigs, whereas Pyrogenes was the second most common serogroup in these three species (Table 3). Titres were most frequently low (⩽400) suggesting a chronic or prior infection. High animal density and humid breeding conditions could explain the high seroprevalence rate of leptospirosis in farm animals. The Panama and Pyrogenes serogroups have been involved in 0·2% and 1·4% of human infections, respectively [11]. Two hypotheses could explain the differential frequencies of these serogroups in humans and animals: the frequency of transmission from pigs, goats, and deer to humans is low, or infections of humans by serogroup Pyrogenes and Panama are asymptomatic or mild, as previously observed in Cambodia and Mexico [Reference Berlioz-Arthaud46, Reference Vado-Solis47] and are therefore rarely diagnosed at laboratory level. The presence of anti-Leptospira antibodies has rarely been reported in cats [Reference Millan30, Reference Agunloye and Nash48], but we report a seroprevalence of 26·6% in this species. Most of the seropositive cats had antibodies to several serogroups, indicating that exposure to multiple strains is frequent in this species and that no specific serogroup seems to be maintained in the cat population.
The main serogroups found in cattle (Sejroe), rats (Icterohaemorrhagiae), and dogs (Canicola) have also been reported to infect the same European species. This finding is not surprising given that all of the farm and wild species living on Reunion Island originated from Europe [Reference Cheke49] and that the black rat introduced to the island is genetically similar to those found in Europe [Reference Tollenaere50]. Considering that leptospires were probably introduced on Reunion Island via the non-native animal hosts that were introduced by humans, it is not surprising to find the same epidemiological patterns of the disease that have been reported in Europe.
MAT is a serogroup rather than a serovar-specific assay and because of the high degree of cross-reactivity that occurs between different serogroups, the serological data should be used only to gain a broad idea of the serogroups present at the population level [Reference Levett51]. Moreover, paradoxical reactions and cross-reactions between serogroups are common [Reference Levett52, Reference Smythe53]. Furthermore, an important limitation of MAT is the number of antigens used, which corresponds to the antigens that are expected to be present in the area being tested and thus minimizes the probability of detecting a response to a serogroup that is not expected [Reference Levett51, Reference Levett52]. Another confounding factor in areas of high endemicity is the possibility of multiple successive or even co-infections with multiple serovars [Reference Levett51]. Culturing and typing of infectious isolates and/or direct molecular typing of strains from clinical samples [Reference Perez and Goarant54] could be used for further epidemiological studies.
To our knowledge, the current study is the first to report the quantification of Leptospira in the kidneys of non-laboratory mammals. We showed that the concentration of leptospires in the kidney of carrier animals presented a large amount of inter-individual variation (standard deviations are shown in Table 5). A large variation in the concentration of leptospires in the urine of experimentally infected rats [Reference Monahan, Callanan and Nally55] and in naturally infected deer [Reference Subharat56] has been described. This variation could be related to variable concentrations of leptospires in the kidney tubules. Various factors could impact the renal concentration of leptospires, including the age of the infected animal [Reference Nally57], other factors inherent in the host [Reference Nally57], the amount of time that has elapsed since infection, and the infecting Leptospira serogroup [Reference Thiermann2]. Moreover, common associations between particular serovars and their animal reservoirs have largely been recognized [Reference Faine17]. Isolation and serotyping or direct molecular typing of the strains would be essential for better understanding of these data. Rats do not develop disease from Leptospira infection [Reference Thiermann2], and our results suggest that naturally infected animals belonging to different species have the same capacity to harbour leptospires in their kidneys. These results constitute the basis of our plan for further studies aimed at evaluating the contribution of each species to Leptospira environmental contamination in the field. At the herd level, the detection or quantification of leptospires in the kidneys of slaughtered animals might be correlated with indicators of herd productivity and could help in the development of veterinary control programmes.
On Reunion Island, the tropical climate and presence of flooded areas increase the environmental exposure of humans and animals to Leptospira [Reference Lau58]. As a consequence, the risk of transmission should be considered at the ecosystem level. Rodents, dogs and most likely shrews are the major sources of human infection whereas livestock and cats seem to mostly maintain leptospire reservoirs of lower relevance to human health. Thus, preventive measures aimed at reducing the burden of leptospirosis in the human population should first focus on relevant control measures against rodents and stray dogs. In livestock, intra-herd transmission could be reduced by the detection of carriers (in particular reproductive animals used for natural breeding) [Reference Lilenbaum4, Reference Vinodh6] and relevant treatment, whereas inter-herd transmission could be reduced by the control of introduced animals, environmental control measures [Reference Martins, Penna and Lilenbaum59], and management of the pasture [Reference Martins, Penna and Lilenbaum59, Reference Schoonman and Swai60]. Knowledge of the animal hosts involved in leptospirosis epidemiology on a small geographical scale contributes to a better understanding of the disease on a global scale.
ACKNOWLEDGEMENTS
The authors are very grateful to I. Sanchez, S. Candat, T. Hue, T. Duval, B. Warren (University of Reunion Island), S. Boyer (IRD), E. Buffard (BNOI/ONCFS), S. Roué, J-M. Probst (Parc National), R. Baronce (FDC974), and the hunters of FDC974 for their help during sampling in the field. We are also grateful to the veterinary services of Reunion Island, Pôle de Protection des Plantes (CIRAD Saint-Pierre), Pôle Elevage (CIRAD Saint-Pierre), the dog pound of Tampon, the slaughterhouse of Saint-Pierre and the slaughterhouse workers for their collaboration, as well as C. Goarant (Pasteur Institute of Noumea) for his helpful advice. We thank the Pasteur Institute of Paris for providing reference strains for MAT; G. Bertil, M. Picard, J. Souprayen, G. Hoarau, and J. Gigan (GHSR) for technical support; and Frédéric Chiroleu for statistical support.
SUPPLEMENTARY MATERIAL
For supplementary material accompanying this paper visit http://dx.doi.org/10.1017/S0950268812002075.
DECLARATION OF INTEREST
None.









