



Introduction
Limestone deposits yielding large lucinid bivalves have been known for centuries from the Miocene deposits in Italy, and were termed ‘Calcari a Lucina’ (Manzoni, Reference Manzoni1876; Coppi, Reference Coppi1877; Scarabelli, Reference Scarabelli1880; Sacco, Reference Sacco1901; Di Stefano, Reference Di Stefano1903). Due to their isolated occurrence in deep-water sediments and the large bivalves preserved in them, they were considered to have been transported from shallow water (Ricci Lucchi and Veggiani, Reference Ricci Lucchi and Veggiani1967). This view changed after the first discovery of faunal communities at methane seeps in the deep Gulf of Mexico with similarly large bivalves (Paull et al., Reference Paull, Hecker, Commeau, Freeman-Lynde, Neumann, Corso, Golubic, Hook, Sikes and Curray1984) and the recognition that methane seep carbonates can be identified based on their distinctive, light carbon isotope signature (Hovland et al., Reference Hovland, Talbout, Qvale, Olausson and Aasberg1987). The ‘Calcari a Lucina’ deposits throughout Italy are now considered as ancient deep-water methane seep deposits (Clari et al., Reference Clari, Gagliardi, Governa, Ricci and Zuppi1988; Conti et al., Reference Conti, Gelmini and Ponzana1993; Terzi, Reference Terzi1993; Aharon and Sen Gupta, Reference Aharon and Sen Gupta1994; Berti et al., Reference Berti, Cuzzani, Landuzzi, Taviani, Aharon and Vai1994; Ricci Lucchi and Vai, Reference Ricci Lucchi and Vai1994; Taviani, Reference Taviani1994; Terzi et al., Reference Terzi, Aharon, Ricci Lucchi and Vai1994; Peckmann et al., Reference Peckmann, Thiel, Michaelis, Clari, Gaillard, Martire and Reitner1999; Clari et al., Reference Clari, Conti, Fontana and Taviani2004b; Conti et al., Reference Conti, Fontana, Gubertini, Sighinolfi, Tateo, Fioroni and Fregni2004, Reference Conti, Fontana, Mecozzi, Panieri and Pini2010). Despite this wealth of geologic literature on these deposits, modern studies on the macrofauna are relatively rare (Moroni, Reference Moroni1966; Taviani, Reference Taviani1994, Reference Taviani2011, Reference Taviani2014; Taviani et al., Reference Taviani, Angeletti and Ceregato2011). The purpose of the present contribution is to provide a revision of the major taxa of chemosymbiotic bivalves of the ‘Calcari a Lucina’ deposits, with exclusion of solemyids.
Materials and methods
Specimens were coated with ammonium chloride for photography. The material is from twelve seep deposits of middle to late Miocene age (Fig. 1) associated with deep-water hemipelagic marls or turbidites, mostly ascribed to the Marnoso-arenacea Formation. Their geological and stratigraphic context is described in various publications (Vai et al., Reference Vai, Taviani, Conti and Aharon1997; Conti and Fontana, Reference Conti and Fontana1999; Clari et al., Reference Clari, Conti, Fontana and Taviani2004b; Taviani, Reference Taviani2011). A short description of the localities is provided in the Appendix.

Figure 1 Maps showing the fossil localities discussed in the text. Left panel, general map of Italy showing all localities: 1=Abisso “Mornig”, 2=Ca’ Carnè, 3=Ca’ Cavalmagra, 4=Monticino-Limisano – Castelnuovo junction, 5=Ca’ Fornace, 6=Ca’ Pianté, 7=Case Rovereti, 8=Castelvecchio, 9=Deruta, 10=Le Colline, 11= Marmorito, 12=Montepetra, 13=Sasso delle Streghe, 14=Verzino.
Repositories and institutional abbreviations
MGGC: Museo Geologico Giovanni Capellini, University of Bologna; MSF: Museo Civico di Scienze Naturali, Faenza; MZB: Museo dell’Evoluzione (formerly Zoologia), University of Bologna; MRSN: Museo Regionale di Scienze Naturali, Torino (managing the Bellardi and Sacco collection, property of the Turin University).
Systematic paleontology
Class Bivalvia Linnaeus, Reference Linnaeus1758
Subclass Pteriomorphia Beurlen, Reference Beurlen1944
Order Mytilida Férussac, Reference Férussac1822
Family Mytilidae Rafinesque, Reference Rafinesque1815
Genus Bathymodiolus Kenk and Wilson, Reference Kenk and Wilson1985
Type species
Bathymodiolus thermophilus Kenk and Wilson, Reference Kenk and Wilson1985, Recent, Galapagos Rift Zone, by original designation.
Remarks
Molecular phylogenetic studies have shown that species currently classified as Bathymodiolus belong to at least two clades within the bathymodiolins (Gustafson et al., Reference Gustafson, Turner, Lutz and Vrijenhoek1998; Jones et al., Reference Jones, Won, Maas, Smith, Lutz and Vrijenhoek2006; Lorion et al., Reference Lorion, Buge, Cruaud and Samadi2010, Reference Lorion, Kiel, Faure, Masaru, Ho, Marshall, Tsuchida, Miyazaki and Fujiwara2013; Thubaut et al., Reference Thubaut, Puillandre, Faure, Cruaud and Samadi2013), of which those related to B. childressi may be placed in a separate genus. This species group is often referred to as the ‘childressi clade’ and it is recommended to use “Bathymodiolus” only in quotation marks for these species until the taxonomic uncertainties are settled (Gustafson et al., Reference Gustafson, Turner, Lutz and Vrijenhoek1998). Morphologically, the two clades can be distinguished based on muscle-scar pattern (Gustafson et al., Reference Gustafson, Turner, Lutz and Vrijenhoek1998), which, unfortunately, is not preserved in any of the fossils reported here from northern Italy. Therefore, we refer to them as Bathymodiolus (sensu lato).
Bathymodiolus (s.l.) moroniae new species

Figure 2 Bathymodiolus (s.l.) moroniae new species from Miocene seep deposits in Italy: (1, 2) holotype (MGGC 21907) from Case Rovereti, isolated right valve; (3) paratype (MZB 27273) from Case Rovereti, internal mold of large specimen, left valve; (4) paratype (MZB 27270) from Case Rovereti, left valve with preserved shell; (5, 6) paratype (MRSN PU 40607.01) from Deruta, internal mold of articulated specimen; (7, 8) paratype (MGGC 21922) from Case Rovereti, left valve of juvenile specimen showing fine taxodont dentition on posterior side of shell; (9) paratype (MSF 2119) from Abisso “Mornig,” left valve, anterior side with shell remains, posterior side internal mold showing pallial line and adductor muscle scar; (10) paratype (MRSN PU 40607.02) from Deruta, left valve with preserved shell; (11) paratype (MSF 1100) from the Monticino-Limisano – Castelnuovo junction, large right valve with preserved shell.
1966Modiolus (Modiolus) exbrocchii exbrocchii Sacco; Reference MoroniMoroni, p. 78, pl. 5, fig. 2, pl. 6, fig. 4.
1996Bathymodiolus exbrocchii Sacco; Reference TavianiTaviani, fig. 2a, 4c.
2001Modiolinid Reference TavianiTaviani, fig. 20.7a.
2011“Bathymodiolus” cf. exbrocchii (Sacco, Reference Sacco1898); Reference TavianiTaviani, fig. 3c.
Type specimens
Holotype: MGGC 21907, single right valve from Case Rovereti. Paratypes: seven specimens from Case Rovereti: MZB 27218, 27268, 27270, 27272, 27273; MGGC 21921, 21922; one from Monticino-Limisano – Castelnuovo junction: MSF 1100; one from Ca’ Piantè (MSF 2135, on same large block as MSF 1360); one from Abisso “Mornig” (MSF 2119).
Diagnosis
Medium- to large-sized “Bathymodiolus” with blunt, elevated umbo at anterior 10% of total shell length; dorsal and ventral margins nearly straight, posterodorsal corner at posterior third of shell; maximum inflation just anterior to anterior third of shell.
Occurrence
Late Miocene seep carbonates in Italy.
Description
Moderately inflated modioliform shell, maximum inflation at ~27% of total shell length; blunt, elevated umbo at anterior 10% of total shell length, anterior margin narrow, convex; ventral margin straight, slightly concave in large specimens; dorsal margin straight to very slightly curved; posterodorsal corner at posterior third of shell; surface smooth except for growth increments; hinge area in juvenile specimens with row of taxodont teeth below the ligament attachment line, smooth in adults; pallial line distant from shell margin, numerous mantle muscle scars toward interior; shell composed of a calcitic fibrous prismatic outer layer and a nacreous inner layer.
Etymology
To honor Maria Antonietta Moroni, for her seminal work on the ‘Calcari a Lucina’ fauna.
Materials
Forty-four valves (including MZB 27064, 27271, 27278, 27281) from Case Rovereti; three specimens from Verzino (MGGC 21923, 21924, 21925); one from Deruta (MGGC 21926); seven articulated shells from Deruta (MRSN, under cf. Bathymodiolus exbrocchii, PU 40607); one articulated specimen from Verzino MGGC 21927; three articulated shells (MSF 1360) plus one valve from Ca‘ Piantè; and specimens from Montepetra; four valves from block from Monticino-Limisano – Castelnuovo junction, Brisighella (MSF 1106-1109); five valves from Abisso “Mornig” (MSF 1086, 1090, 1094, 1097, 2120); see Table 1 for measurements.
Table 1 Measurements of Bathymodiolus (s.l.) moroniae new species; H=height, L=length, W=width of two valves, except when indicated otherwise.

Remarks
The assignment of this species to Bathymodiolus (sensu lato) is based on its general shape, the change from a hinge with denticles in young specimen to an edentulous hinge in adults, its shell microstructure (cf., Génio et al., Reference Génio, Kiel, Cunha, Grahame and Little2012), and its mass occurrence at seep deposits. The early juvenile shell shape and adductor and retractor muscle scars, which have been used to identify other fossil bathymodiolins (Kiel, Reference Kiel2006; Kiel and Goedert, Reference Kiel and Goedert2007; Saether et al., Reference Saether, Little, Campbell, Marshall, Collins and Alfaro2010; Amano and Jenkins, Reference Amano and Jenkins2011; Kiel and Amano, Reference Kiel and Amano2013) are not preserved in Bathymodiolus (s.l.) moroniae n. sp.
Nelli (Reference Nelli1903) and Moroni (Reference Moroni1966) had identified this species as Modiolus (Modiolus) exbrocchii exbrocchii Sacco, Reference Sacco1898, which was named by Sacco (Reference Sacco1898) based on an illustration of Modiola brocchii Mayer in Hörnes, Reference Hörnes1870 from the Vienna Basin (Hörnes, Reference Hörnes1870, p. 345, pl. 45, fig. 13a, b). However, this species differs from B. (s.l.) moroniae by being more elongate and by having a distinct and sharp ridge running from the umbo to the posteroventral margin, whereas B. (s.l.) moroniae is smoothly convex. Furthermore, specimens of B. moroniae with the size of M. exbrocchii as figured by Hörnes (11 cm) are more compact and less elongate that M. exbrocchii.
Most similar in general shell shape are specimens of B. brevior von Cosel, Métivier, and Hashimoto, Reference von Cosel, Métivier and Hashimoto1994 living at hydrothermal vents in the Indian Ocean (this population was formerly called B. marisindicus Hashimoto, Reference Hashimoto2001, but later synonymized with B. brevior, based on molecular data), and B. puteoserpensis von Cosel, Métivier, and Hashimoto, Reference von Cosel, Métivier and Hashimoto1994 from the Mid-Atlantic Ridge (von Cosel et al., Reference von Cosel, Métivier and Hashimoto1994; Hashimoto, Reference Hashimoto2001). Another similar species is B. brooksi from the Gulf of Mexico (Gustafson et al., Reference Gustafson, Turner, Lutz and Vrijenhoek1998). Interestingly, these three morphologically most similar species all belong to Bathymodiolus sensu strictu and not to the childressi clade.
Unlike bathymodiolins at most other fossil seep deposits, Bathymodiolus (s.l.) moroniae n. sp. occurs mostly as disarticulated valves, especially in adult stages.
Bathymodiolus (s.l.) miomediterraneus new species

Figure 3 Bathymodiolus (s.l.) miomediterraneus new species from the middle Miocene seep deposit at Le Colline, northern Italy: (1) paratype (MSF 1353) with strongly concave ventral margin, right valve; (2) holotype (MGGC 21908) showing broadly arched dorsal margin, left valve; (3, 4) paratype (MSF 1352), small specimen showing right valve and in ventral view, respectively; (5) block with two paratypes (MSF 1351), view of left valves; (6) strongly arched, gerontic specimen (MSF 1190).
Type specimens
Holotype: MGGC 21908; paratypes: MSF 1351 (two articulated shells), MSF 1352 (two articulated shells, one figured), MSF 1353 (right valve), MSF 1190 (left valve); all types from Le Colline.
Diagnosis
Small- to medium-sized “Bathymodiolus” with subterminal umbo, broad anterior margin, and arched dorsal margin; maximum inflation just anterior to middle of shell; blunt ridge running from umbo to posteroventral corner.
Occurrence
Middle Miocene (early Serravallian) seep carbonates at Le Colline in northern Italy.
Description
Well inflated, moderately sized, arched modioliform shell with low, subterminal umbo; maximum inflation just anterior of center; anterior margin broad and well rounded, posterodorsal margin broadly arched in large specimens, smaller specimens with distinct posterodorsal corner, ventral margin slightly concave in small specimens, strongly concave in large ones; posterior margin gently rounded; blunt ridge running from umbo to posteroventral corner; surface smooth except for growth increments.
Etymology
Referring to the Miocene Mediterranean Basin.
Materials
Three specimens from Le Colline (MSF 2136); see Table 2 for measurements.
Table 2 Measurements of Bathymodiolus (s.l.) miomediterraneus new species, all specimens from Le Colline, northern Italy; H=height, L=length, W=width of two valves, except when indicated otherwise.
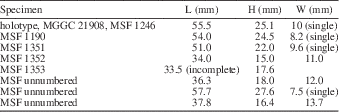
Remarks
Similar in shape is ‘Modiola’ pistacina Sacco, Reference Sacco1904 from the Burdigalian of Piedmont (Sacco, Reference Sacco1904, pl. 29, fig. 6a, b; Merlino, Reference Merlino2007, pl. 6, figs. 8, 9); the type specimen is small (~13 mm long) and has the umbo in a more posterior position and a broader anterior margin than B. miomediterraneus n. sp. ‘Modiola’ pistacina may be related to small bathymodiolins such as “Idas” and should be further investigated in the context of whale- and wood-fall communities in the Italian Neogene. The middle Miocene ‘Modiola’ exbrocchii (e.g., the specimen illustrated as Modiola brocchii by Hörnes, Reference Hörnes1870) and its variation M. exbrocchii var. tauroparva Sacco, Reference Sacco1898, a small species of ~20 mm length (Sacco, Reference Sacco1898, pl. 11, figs. 28, 29; Merlino, Reference Merlino2007, pl. 6, fig. 7), have a more elevated umbo and a more distinctive ridge running from the umbo to the posterior-ventral margin than B. miomediterraneus. The new species differs from Bathymodiolus (s.l.) moroniæ n. sp. by having the umbo in a more anterior (subterminal) position. The most similar Recent species are Bathymodiolus mauritanicus von Cosel, Reference von Cosel2002, which occurs off West Africa and in the Gulf of Cadiz (von Cosel, Reference von Cosel2002; Génio et al., Reference Génio, Johnson, Vrijenhoek, Cunha, Tyler, Kiel and Little2008) and appears to have a broader posterior margin than B. (s.l.) miomediterraneus n. sp., and the Japanese Bathymodiolus hirtus Okutani, Fujikura, and Sasaki, Reference Okutani, Fujikura and Sasaki2004, which has a more elevated umbo as adult than B. (s.l.) miomediterraneus n. sp. and is broader (less slender) as juvenile (Okutani et al., Reference Okutani, Fujikura and Sasaki2004). A fossil species with a similar subterminal umbo is the presumably Oligocene Bathymodiolus (s.l.) palmarensis from Colombia (Kiel et al., Reference Kiel, Campbell and Gaillard2010), but it differs from B. (s.l.) miomediterraneus n. sp. by being more elongate and by having almost straight anterior and posterior margins. Bathymodiolus (s.l.) miomediterraneus n. sp. is typically found with articulated valves, in contrast to Bathymodiolus (s.l.) moroniae n. sp. and Samiolus iohannesbaptistae n. gen. n. sp.
Genus Samiolus new genus
Type species
Samiolus iohannesbaptistae new genus new species, from late Miocene seep deposits in northern Italy.
Diagnosis
Small- to medium-sized modioliform shell; strong, almost angular ridge from umbo to posteroventral corner; posterior part of shell sculptured by roughly commarginal wrinkles, unlike any other mytilid.
Etymology
The name combines the name Sami, crediting the paleontologist Marco Sami, who was responsible for collecting a substantial number of valuable macrofossils from ‘Calcari a Lucina’, including this new taxon, with the mytilid genus Modiolus, which has a similar shell shape.
Remarks
Samiolus n. gen. contains presently only the type species. A similar strong and sharp ridge can be seen in Gibbomodiola taurarcuata Sacco, Reference Sacco1898 from the Italian Miocene (Sacco, Reference Sacco1898, pl. 11, figs. 34a, 35b; Merlino, Reference Merlino2007, pl. 6, fig. 10), but this species lacks wrinkles (own observations on Sacco’s type material housed in the Turin Museum). Another species with a similar ridge is Modiola dombraviensis Kittl, Reference Kittl1887 from middle Miocene ‘blue deep-water clays’ in eastern Czech Republic and southern Poland (Kittl, Reference Kittl1887, p. 272, pl. 9, figs. 7–11). But Modiola dombraviensis has a nearly smooth surface with only fine growth increments and lacks the wrinkled sculpture of Samiolus iohannesbaptistae n. sp. (own observation of material housed in the Natural History Museum Vienna). This wrinkled surface sculpture distinguishes Samiolus iohannesbaptistae n. gen. n. sp. also from all fossil and extant bathymodiolins (Squires and Goedert, Reference Squires and Goedert1991; von Cosel and Olu, Reference von Cosel and Olu1998; Gustafson et al., Reference Gustafson, Turner, Lutz and Vrijenhoek1998; von Cosel, Reference von Cosel2002; Sasaki et al., Reference Sasaki, Okutani and Fujikura2005; Desbruyères et al., Reference Desbruyères, Segonzac and Bright2006; von Cosel and Janssen, Reference von Cosel and Janssen2008; Lorion et al., Reference Lorion, Buge, Cruaud and Samadi2010; Amano and Jenkins, Reference Amano and Jenkins2011).
Samiolus iohannesbaptistae new species

Figure 4 Samiolus iohannesbaptistae new genus new species from late Miocene seep deposits in northern Italy: (1–4) paratype (MSF 1073) from Ca’ Carnè, four views on right valve; (5, 6) interior of small specimen (MGGC 23010) from Ca’ Carnè, showing hinge dentition of right valve; (7–10) paratype (MSF 1120) from Ca’ Carnè, right valve; (11) large specimen (MSF 1355) from Ca’ Piantè, left valve; (12–16) holotype (MGGC 21910) from Ca’ Carnè, five views of right valve.
Type specimens
Holotype MGGC 21910 from Ca’ Carnè; paratypes: three specimens from Ca’ Carnè: MSF 1073, MSF 1120 (left valve), MZB 60090; one specimen from Ca’ Piantè: MSF 1355.
Diagnosis
Same as generic diagnosis.
Occurrence
Late Miocene (late Tortonian) seep carbonates in northern Italy.
Description
Modioliform, small- to moderately sized shell, umbo subterminal; posterodorsal margin slightly arched, posterodorsal corner at ~60% total shell length, ventral margin convex; strong ridge starting posterior of umbo, running to posteroventral corner, anterodorsal margin of ridge almost angular, followed by broad groove; shell surface anterior of ridge smooth except for growth lines, posterior side shows ribbing or wrinkles, resulting from irregular, roughly commarginal thickenings. Juvenile specimen with fine taxodont hinge teeth along entire posterodorsal margin and on oval hinge plate below beak; hinge area in adult smooth. Largest specimen 33 mm long, 17 mm high, single valve 7 mm wide; a juvenile specimen with taxodont hinge teeth measures 8.3 mm in length.
Etymology
To honor Gian Battista Vai in recognition of his leading role in the modern study of ‘Calcari a Lucina.’
Materials
Eight specimens from Ca’ Carnè; six from Ca’ Piantè; three valves from Montepetra (MZB 60095), and one valve from Case Rovereti (MSF 1356); see Table 3 for measurements.
Table 3 Measurements of Samiolus (s.l.) iohannesbaptistae new genus and species; H=height, L=length, W=width of two valves, except when indicated otherwise.
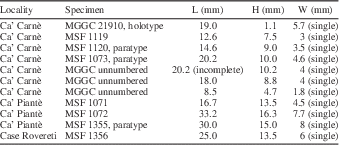
Remarks
We have found mostly disarticulated specimens of this species. One specimen has been figured by Sami and Taviani (Reference Sami and Taviani2015, fig. 7).
Subclass Heterodonta Neumayr, Reference Neumayr1884
Family Thyasiridae Dall, Reference Dall1900
Genus Channelaxinus Valentich-Scott and Coan in Coan and Valentich-Scott, Reference Coan and Valentich-Scott2012
Type species
Channelaxinus oliveri Valentich-Scott and Coan in Coan and Valentich-Scott (Reference Coan and Valentich-Scott2012), (by original designation); Recent, Bahía Santiago, Colima, México.
Channelaxinus sp.

Figure 5 Channelaxinus sp. from the middle Miocene (Langhian) of Ca’ Cavalmagra, northern Italy: (1, 2) left valve (MSF 2113), internal mold and counterpart with small amount of shell preserved; (3) left valve (MSF 1287), internal mold with few shell remains; (4) right valve (MSF 2112) of small specimen.
Description
Shells subrectangular, thin-shelled, reaching 20 mm in length and 16 in height, only slightly inflated, sculptured by irregular growth lines and indistinct, blunt ridge running from umbo to postero-ventral corner; beak prosogyrate, projecting slightly above dorsal margin; anterior margin straight to slightly concave, ventral margin gently convex, postero-ventral corner well rounded, posterior margin gently convex below sulcus, postero-dorsal margin slightly convex, with deep sinus at end of posterior sulcus; posterior sulcus deep, bordered by sharp ridge, area between sulcus and postero-dorsal margin narrow, with narrow submarginal sulcus; anterior adductor muscle scar thin, elongate, weakly impressed, separated from pallial line, length just over half of shell length; interior radiating striae distinct.
Materials
Three specimens from Ca’ Cavalmagra (MSF 1287, 2112, 2113).
Occurrence
Only known so far from the late Langhian of Ca’ Cavalmagra.
Remarks
A superficially similar thyasirid from the Miocene of the Turin Hills is Thyasira michelottii (Hörnes, Reference Hörnes1875), but it clearly differs from Channelaxinus sp. reported here by having a much shorter anterior adductor muscle scar (Zuschin et al., Reference Zuschin, Mandic, Harzhauser and Pervesler2001). The extant type species Channelaxinus oliveri differs from Channelaxinus sp. mainly by having a narrower angle between the anterior and dorsal margins and a blunter (less pointed) umbo (Coan and Valentich-Scott, Reference Coan and Valentich-Scott2012; Oliver and Frey, Reference Oliver and Frey2014). This is the first fossil record of Channelaxinus.
Family Lucinidae Fleming, Reference Fleming1828
Genus Meganodontia Bouchet and von Cosel, Reference Bouchet and von Cosel2004
Type species
Meganodontia acetabulum Bouchet and von Cosel, Reference Bouchet and von Cosel2004 (by original designation); Recent, in 256–472 m depth off northern Taiwan.
Meganodontia hoernea (Des Moulins, Reference Des Moulins1868)

Figure 6 Meganodontia hoernea (Des Moulins, Reference Des Moulins1868) from the late Miocene seep deposit at Ca’ Piantè in northern Italy: (1) internal mold of left valve (MGGC 23003) showing the elongate anterior adductor muscle scar; (2) dorsal view of same specimen as Figure 6.1; (3) left valve in foreground showing posterodorsal shell margin, right valve in background showing edentulous hinge (MGGC 23004); (4) outer shell of right valve seen in the background of Figure 6.3; (5) hinge and ligament nymph in left valve (MGGC 23005).

Figure 7 Meganodontia hoernea (Des Moulins, Reference Des Moulins1868), specimens from Sacco’s collection from the Turin Hills: (1–3) left valve (MRSN BS 154.01.002) from Albugnano, with well-preserved shell material, showing hinge area and external sculpture; (4) view of left valve of internal mold (MRSN PU 40603.01) from an unknown locality, showing the elongate anterior adductor muscle scar and the internal pustules; (5, 6) right valve (MRSN BS 154.01.001) from Albugnano, showing external sculpture.
1868Lucina hoernea Reference Des MoulinsDes Moulins, p. 368.
1901Lucina globulosa var. hörnea Reference SaccoSacco, p. 67, pl. 15, figs. 31–33, pl. 16, fig. 1.
1901Lucina globulosa var. alta Reference SaccoSacco, p. 68, pl. 15, fig. 4.
1901Lucina globulosa var. perinaequilatera Reference SaccoSacco, p. 68, pl. 15, fig. 5.
1966Lucina hoernea; Reference MoroniMoroni, p. 80, pl. 3, fig. 3.
2001Lucina Reference TavianiTaviani, fig. 20.1.
2011“Lucina” sp. Reference TavianiTaviani, fig. 3a.
Description
Very large, globular shells, height/length-ratio 0.9; umbones elevated, prosogyrate; external sculpture of fine, irregular, commarginal growth lines, posterodorsal sulcus shallow but distinct; lunule lanceolate, bordered by sharp, smooth ridge. Hinge plate narrow to moderately broad, edentulous, ligament nymph narrow. Anterior adductor muscle scar elongate, detached from pallial line for ~80% of its length, angle of deviation ~30°, width about half the distance to pallial line, curved in the opposite direction as shell margin, reaching below midline to ~44% of total shell height; somewhat constricted posteriorly just below the point of detachment. Interior of shell with numerous pustules, often in rows radiating form umbo; pallial blood vessel scar visible.
Materials
Eight specimens from Ca’ Piantè (MSF 1357; 2100–2104, 2110); 45 articulated shells from Deruta (as Lucina globulosa hoernea Desmoulins, Bellardi and Sacco collection in MRSN, PU 40597–40601); one articulated shell from Bersano (as Lucina globulosa [Deshayes] var hoernea, Bellardi and Sacco collection in MRSN, BS 154.01.001); one articulated shell from Albugnano (as Lucina globulosa [Deshayes] var hoernea Desmoulins, Bellardi and Sacco collection in MRSN, BS.154.01.002); two articulated shells from Pino Torinese (as Lucina globulosa [Deshayes] var hoernea Desmoulins, Bellardi and Sacco collection in MRSN, BS.154.01.003 and BS.154.01.004); one articulated shell from Pera del Gal, Turin Hills (as Lucina globulosa [Deshayes] taurofuchsi Sacco, Reference Sacco1901, Bellardi and Sacco collection in MRSN, BS.154.01.007); one articulated shell from Pino Torinese (as Lucina globulosa [Deshayes] alta Sacco, Reference Sacco1901, Bellardi and Sacco collection in MRSN, BS.154.01.008); one articulated shell from Pino Torinese (as Lucina globulosa [Deshayes] perinaequilatera Sacco, Reference Sacco1901, Bellardi and Sacco collection in MRSN BS.154.01.009); more than 20 articulated shells (molds) from Sasso delle Streghe (Modena), a dozen articulated shells from other localities of the Modena and Forlì Apennines; a dozen articulated shells from the Turin Hills (Sacco’s collection in MRSN); see Table 4 for measurements.
Table 4 Measurements of Meganodontia hoernea (Des Moulins, Reference Des Moulins1868), all specimens from Ca’ Piantè, northern Italy; H=height, L=length, W=width of two valves, except when indicated otherwise.
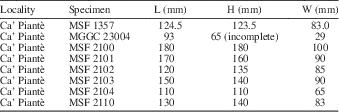
Occurrence
Middle to late Miocene seep carbonates in Italy.
Remarks
Bouchet and von Cosel (Reference Bouchet and von Cosel2004) considered “Lucina” hoernea not to belong to Meganodontia because it has more globose and prominent umbones than M. acetabulum and because internal features of “Lucina” hoernea were not known to them. Here, we show that “Lucina” hoernea has an edentulous and narrow hinge plate, as well as an elongate, blade-shaped anterior adductor muscle scar just like Meganodontia acetabulum. Therefore we consider “Lucina” hoernea as belonging to Meganodontia, but as distinct from M. acetabulum. In addition to the differences mentioned above, Meganodontia hoernea has a less distinctive posterior adductor muscle scar than Meganodontia acetabulum. Moroni (Reference Moroni1966) provided an extensive synonymy for Meganodontia hoernea. In the Meganodontia species from the early Miocene of Cuba (Kiel and Hansen, Reference Kiel and Hansen2015), the anterior adductor muscle scar deviates from the pallial line at a broader angle, as in M. hoernea. Examination of Sacco’s type material showed that his “variations” L. globulosa var. alta Sacco, Reference Sacco1901 and L. globulosa var. perinaequilatera Sacco, Reference Sacco1901 represent deformed specimens of Meganodontia hoernea and should therefore be considered as synonyms.
Taylor et al. (Reference Taylor, Glover, Smith, Dyal and Williams2011) tentatively assigned Meganodontia to their new subfamily Pegophyseminae, whose species are united by some unusual morphological characters, including thin, globular, smooth shells, a narrow edentulous hinge with an internal and laterally extended ligament in most species (Taylor et al., Reference Taylor, Glover, Smith, Dyal and Williams2011). Meganodontia hoernea is a common and widespread species in the Miocene of Italy, associated with ‘Calcari a Lucina’ limestones and reducing deep-sea marly sediments.
Genus Lucinoma Dall, Reference Dall1901
Type species
Lucina filosa Stimpson, Reference Stimpson1851 (by original designation); Recent, northwest Atlantic.
Lucinoma perusina (Sacco, Reference Sacco1901)

Figure 8 Lucinoma perusina (Sacco, Reference Sacco1901) from the late Miocene type locality at Deruta, northern Italy: (1–3) lectotype (MRSN BS.154.03.034), originally illustrated as “Dentilucina perusina var. pseudorotunda” (Sacco, Reference Sacco1901, pl. 19, fig. 15); (4) large internal mold (MRSN PU 40604.01) showing muscle scars and pallial line in right valve; (5) right valve of medium-sized specimen (MRSN PU 40604.02) with defoliated outer shell layer, giving it a smooth appearance; (6) right valve of small specimen (MRSN PU 40606.01) showing surface sculpture; (7) right valve of small specimen (MRSN PU 40606.02).

Figure 9 Lucinoma perusina (Sacco, Reference Sacco1901) from late Miocene seep deposits in northern Italy: (1) partial internal mold of a left valve (MGGC 23006) showing anterior adductor muscle scar, from Ca’ Piantè; (2) dorsal view of same specimen as in Figure 9.1, showing lunule; (3) internal mold of left valve (MGGC 23007) showing anterior adductor muscle scar, from Ca’ Piantè; (4) left valve (MGGC 23008) showing external sculpture, from Ca’ Piantè; (5) hinge of same specimen as in Figure 9.4; (6) hinge of a large specimen (MSF 2116) from Ca’ Fornace; (7) block from Ca’ Fornace with several specimens (MSF 2118) showing their external surfaces; (8) dorsal view showing umbones, lunule and escutcheon, middle specimen from Figure 9.7.
1901Dentilucina perusina Reference SaccoSacco, p. 83, pl. 19, figs. 12–14.
1901Dentilucina perusina var. pseudorotunda Reference SaccoSacco, pl. 21, fig. 15.
1966Phacoides (Lucinoma) perusinus (Sacco); Reference MoroniMoroni, p. 82, pl. 5, figs. 1, 3, pl. 6, figs. 1, 2.
1996Lucinidae Reference TavianiTaviani, fig. 4b.
Description
Medium- to large-sized Lucinoma, outline of shell oval in small specimens, becoming more round in large ones; umbones central, prosogyrate, elevated; surface sculptured by fine, irregular growth increments or commarginal ribs, posterior sulcus only weakly developed; lunule elongate, heart-shaped, moderately excavated, bordered by distinct angulation; escutcheon lanceolate, wide; anterior adductor muscle scar thin, elongate, reaching well below midline of shell, detached from pallial line for more than three-quarters of its length, deviates from pallial line by ~10°; posterior adductor muscle scar oval, dorsally pointed; hinge plate broad, two strong, radiating cardinal teeth in each valve, one anterior lateral tooth in each valve, posterior lateral teeth not seen.
Materials
Ten specimens from Ca’ Piantè and six specimens from Santa Sofia; six shells from Montepetra, all articulated; one specimen on block with articulated Archivesica sp. (MSF 2111); four articulated shells (MSF 2114–2117) and a cluster (MSF 2118) from erratic blocks in Sintria Creek, near Ca’ Fornace; 26 articulated shells from Deruta (as Dentilucina cf perusina Deshayes, Bellardi and Sacco collection in the Turin Museum, PU 40604); ten articulated shells from Deruta (as Lucina globulosa Deshayes, Bellardi and Sacco collection in the Turin Museum, PU 40602); one articulated shell from Deruta (Bellardi and Sacco collection in the Turin Museum, as Dentilucina perusina Sacco pseudorotunda Sacco, Reference Sacco1901, BS.154.03.034) ; see Table 5 for measurements and specimen numbers.
Table 5 Measurements of Lucinoma perusina (Sacco, Reference Sacco1901); H=height, L=length, W=width of two valves, except when indicated otherwise.

Occurrence
Middle to late Miocene seep carbonates, from northern Italy to Sicily.
Remarks
There is no formal description of Dentilucina perusina in Sacco (Reference Sacco1901); he used this name in the discussion of Dentilucina barrandei (Mayer, Reference Mayer1871), and illustrated three specimens as D. perusina, including one as Dentilucina perusina var. pseudorotunda Sacco, Reference Sacco1901; all specimens are from Deruta. The figured specimens of D. perusina sensu strictu are not present at MRSN and are presumably lost; here, we illustrate the specimen previously illustrated as Dentilucina perusina var. pseudorotunda Sacco (Reference Sacco1901, pl. 21, fig. 15) and designate it as lectotype for Dentilucina perusina. Sacco’s illustrated specimens of D. perusina sensu strictu are more oval than the rather round D. p. pseudorotunda; however, virtually all of the 37 specimens of Bellardi and Sacco collection from Deruta (housed in MRSN) are of the round shape of D. p. pseudorotunda. Hence, we consider this as the typical shape of D. perusina. Moroni (Reference Moroni1966) provided an extensive synonymy for this species.
Externally similar is the Miocene “Dentilucina” barrandei from the Turin Hills, but its cardinal teeth are smaller than in L. perusina and they are nearly parallel, in contrast to the radiating teeth in D. perusina. Among the extant species, Lucinoma kazani Salas and Woodside, Reference Salas and Woodside2002 living at seeps in the Mediterranean Sea, has a similar shell but less inflated, a similarly short and well-defined lunule, and similar fine commarginal ribs, whereas the North Atlantic-Mediterranean L. borealis has a broader and more elongate lunule, and its ribs are more lamellar than in L. perusina and L. kazani (Salas and Woodside, Reference Salas and Woodside2002). None of the Lucinoma species known from Japan are particularly similar to L. perusina (Okutani, Reference Okutani2000).
Family Vesicomyidae Dall and Simpson, Reference Dall and Simpson1901
Genus Archivesica Dall, Reference Dall1908
Type species
Callocardia gigas Dall, Reference Dall1895 (by original designation); Recent, Gulf of California.
Remarks
There is currently little consensus which species belong to the genus Archivesica. While the concept of Amano and Kiel (Reference Amano and Kiel2007), which includes middle Eocene to Recent species is probably too broad (cf., Amano and Kiel, Reference Amano and Kiel2012), the view of Krylova and Sahling (Reference Krylova and Sahling2010) to include only two extant species in addition to the type, may be a little too narrow. In a recent molecular phylogenetic analysis (Audzijonyte et al., Reference Audzijonyte, Krylova, Sahling and Vrijenhoek2012), a monophyletic clade called the ‘gigas group’ emerged and included 14 named and five unnamed species, including the type species of Archivesica. Because the species described below are morphologically most similar to several members of this clade—Archivesica soyoae (Okutani, Reference Okutani1957) A. kilmeri (Bernard, Reference Bernard1974), A. okutanii (Kojima and Ohta, Reference Kojima and Ohta1997), and A. kawamurai (Kuroda, Reference Kuroda1943)—we assign them to Archivesica. Other authors (Krylova and Janssen, Reference Krylova and Janssen2006; Krylova and Sahling, Reference Krylova and Sahling2010) have separated these species into the genera Archivesica, Phreagena, and Akebiconcha, but they have not consistently emerged as monophyletic groups in molecular phylogenetic studies (Audzijonyte et al., Reference Audzijonyte, Krylova, Sahling and Vrijenhoek2012; Valdés et al., Reference Valdés, Sellanes and D’Elía2013).
The first claim of the occurrence of large deep-water vesicomyids in the Mediterranean Miocene was based on the discovery in 1992 (Taviani et al., Reference Taviani, Aharon and Vai1992; Taviani, Reference Taviani1996) of a bed containing ‘Calyptogena’ (=Archivesica) associated with turbidites in the Romagna Apennines (for details see Berti et al., Reference Berti, Cuzzani, Landuzzi, Taviani, Aharon and Vai1994; Taviani, Reference Taviani2014). However, large vesicomyids such as those reported here are not rare in some well-studied outcrops and their occurrence could not have escaped the attention of former investigators. They most likely were identified as Lutraria or veneroideans, or, as in the case of the Bellardi and Sacco collection at MRSN from Deruta, as cf. “Tellinidae?”. In this respect, Taurotapes craverii (Michelotti, Reference Michelotti1847) from the lower Miocene of the Turin area (Sacco, Reference Sacco1904) deserves special attention. Elongated bivalves were reported under this name from a putative Miocene hydrocarbon seep at Roccapalumba (Nicosia, Reference Nicosia1956), which most likely represent Archivesica-like vesicomyids (unpublished observation, M. Taviani, 2016).
Archivesica aharoni new species

Figure 10 Archivesica aharoni new species from Miocene seep deposit in Italy: (1) holotype (MGGC 21909), semi-articulated specimen, internal mold of left valve showing anterior adductor muscle scar, and interior of right valve, from Ca’ Pianté; (2) paratype (MSF 1048), internal view of left valve, from Ca’ Pianté; (3) paratype (MSF 1051), right valve with partially preserved shell, from Ca’ Pianté; (4) paratype (MZB 23846), internal mold of left valve, from Ca’ Rovereti; (5) paratype (MSF 1046), exterior of left valve, from Ca’ Rovereti; (6) paratype (MGGC 21928), rubber cast of right valve showing hinge and the lacking pallial sinus, from Marmorito.

Figure 11 Archivesica aharoni new species from Miocene seep deposit in Italy: (1, 2) holotype (MGGC 21909) from Ca’ Pianté, dorsal view, and close-up of hinge of right valve, respectively; (3) paratype (MSF 1048) from Ca’ Pianté, close-up of hinge of left valve; (4) paratype (MGGC 21928) close-up of hinge of right valve; (5) paratype (MGGC 23009) from Verzino, close-up of hinge of left valve.
1994Calyptogena n. sp. Reference TavianiTaviani, fig. 3e.
1996?Calyptogena sp. Reference TavianiTaviani, figs. 3a, b.
2001Calyptogena sp. Reference TavianiTaviani, fig. 20.9b.
2011Calyptogena n. sp. Reference TavianiTaviani, fig. 3e.
Type specimens
Holotype MGGC 21909 from Ca’ Piantè. Paratypes: three specimens from Ca’ Piantè (MSF 1048, 1051, 1053); one from Marmorito (MGGC 21928); two from Case Rovereti (MZB 23846, 28075); three from Verzino (MZB 28076, 28077, 23009); see Table 6 for measurements.
Table 6 Measurements of Archivesica aharoni new species; H=height, L=length, W=width of two valves.
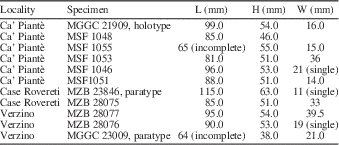
Diagnosis
Large Archivesica with pointed posterior margin, often with posterior ridge and sulcus below; anterior adductor muscle scar large and round, right valve with three radiating, equally sized teeth.
Occurrence
Late Miocene seep carbonates in Italy.
Description
Moderately large (up to 115 mm long), elongate-veneriform shell with umbo anterior at ~28% total shell length; anterior side strongly convex, slightly pointed; ventral side evenly curved; posterodorsal margin only slightly convex, transition to posterior margin at clear angulation; posterior margin pointed at its ventral side. Beak elevated, blunt, slightly prosogyrate; external sculpture of rough growth lines only; no lunula or lunular incision, escutcheon narrow with sharp edges, length about two-thirds of posteroventral margin. Anterior adductor muscle scar distinct and round, pallial line starting at its posteroventral corner, distinct only in anterior half of shell. Hinge plate high but short; RV with three equally strong cardinal teeth that radiate anteroventrally, a distinctive inverted-V-shaped groove between hinge teeth and nymph plate forms the subumbonal pit; cardinal 1 straight, oblique to anterodorsal margin, cardinals 3a and 3b not connected, 3a subparallel to anterodorsal margin, 3b almost at right angle to it; nymph plate narrow. LV with three teeth, cardinal 2a parallel to anterodorsal margin, wedge-shaped, tapering anteriorly, 2b short, perpendicular to 2a, 4 short, wedge-shaped, nearly perpendicular to ventral shell margin.
Etymology
For Paul Aharon, Tuscaloosa, Alabama, in recognition of his relevant work on Gulf of Mexico recent cold seeps and ‘Calcari a Lucina’ fossil counterparts.
Materials
Two specimens from Ca’ Piantè (MSF 1055, 1358) and four shells from Verzino (MGGC 23002).
Remarks
The two most similar species are the extant Japanese Archivesica soyoae and A. kawamurai. The former differs from A. aharoni n. sp. mainly by having a broader nymph plate and the teeth in the left valve are more elongate (Sasaki et al., Reference Sasaki, Okutani and Fujikura2005). In addition to the left valve teeth of Archivesica kawamurai being more elongate than in A. aharoni n. sp., the teeth of the right valve are more slender (Sasaki et al., Reference Sasaki, Okutani and Fujikura2005; Amano and Kiel, Reference Amano and Kiel2010). The geographically close Phreagena sp. from a ca. 25,000 years old vent field on the Mid-Atlantic Ridge (Lartaud et al., Reference Lartaud2010) differs from A. aharoni n. sp. by having the anterior onset of the pallial line in a more anterior position, cardinal tooth 3b in the right valve points posteriorly rather than ventrally as in A. aharoni n. sp., and its escutcheon is much broader. The late Miocene A. shikamai Amano and Kiel, Reference Amano and Kiel2010 from Japan differs by generally having straighter and more parallel dorsal and ventral margins.
Archivesica aff. aharoni Kiel and Taviani, herein

Figure 12 Archivesica aff. aharoni new species from the middle Miocene seep deposit at Deruta in northern Italy: (1) left valve of small specimen (MRSN PU 40611.02) showing shell outline; (2) right valve of medium-sized specimen (MRSN PU 40609.01) showing shell outline; (3) large specimen (MRSN PU 40608-01) showing muscle attachment scars and pallial line; (4–6) moderately sized specimen (MRSN PU 40608.02), (4) view of left valve showing pointed posterior end, muscle attachment scars and pallial line, (5) dorsal view showing hinge area, (6) close-up of hinge area; (7, 8) large specimen (MRSN PU 40608.03), (7) view of left valve showing anterior muscle attachment scars and pallial line, (8) dorsal view showing hinge area; (9) (MRSN PU 40609.01) detached hinge region showing fractured teeth of the right valve and molds of the teeth of the left valve.
Description
Shells up to 94 mm long, elongate-veneriform shell with umbo anterior at ~26–35% total shell length; anterior margin either pointed or truncate; ventral margin evenly curved; posterodorsal margin almost straight to slightly convex, transition to posterior margin with clear angulation in specimens with truncate posterior margin, or more rounded in specimens with pointed posterior margin. Beak elevated, blunt, slightly prosogyrate; external sculpture of rough growth lines only; no lunula or lunular incision. Anterior adductor muscle scar distinct and round, pallial line starting at its posteroventral corner, pallial sinus small, shallow; posterior adductor muscle scar large, round, with anteriorly pointed hook at dorsal margin. Hinge plate high but short; RV with three equally strong cardinal teeth that radiate anteroventrally; LV with three teeth, cardinal 2a wedge-shaped, tapering anteriorly, parallel to anterodorsal margin, 2b and 4 short.
Materials
Four lots of articulated bivalves, mostly molds, but some with preserved portions of the shell, labeled as “cf. Tellinidae (?)” in the Bellardi and Sacco collection at MRSN (N40608: 20 specimens, largest specimen L×H×W=100×54×35 mm; PU 40609: 25 specimens, largest specimen 82×38×20 mm; N40610: 30 specimens, largest specimen 79×40×24 mm; PU 40611: 16 specimens., largest specimen 45×22×6 mm); four additional specimens (MGGC 21930, 21931; MZB 60096, 60097).
Occurrence
Middle Miocene (Serravallian) seep carbonate at Deruta in northern Italy.
Remarks
These specimens share all features of A. aharoni, but are slightly smaller and consistently lack the posterior ridge and groove that can be seen in many specimens of A. aharoni from the type locality. Furthermore, the posterior part of the pallial line and the posterior adductor muscle scar can be more clearly seen than in specimens of A. aharoni from the type locality, but this might be a taphonomic artifact.
Archivesica strigarum new species

Figure 13 Archivesica strigarum new species from the middle to late Miocene seep deposits in northern Italy: (1) paratype (MGGC 21929) from Sasso delle Streghe, left valve showing the round anterior adductor muscle scar; (2–5) holotype (MZB 28095) from Sasso delle Streghe, internal mold; (2) left valve, (3) dorsal view, (4) close-up of hinge area, (5) silicon rubber cast of hinge area; (6, 7) paratype (MRSN PU 40611.01), small specimen from Deruta. Abbreviations: aprs, anterior pedal retractor scar; LV, left valve; RV, right valve.
2001 Calyptogena sp. Reference TavianiTaviani, fig. 20.8a.
Type specimens
Holotype: MZB 28095, internal mold from Sasso delle Streghe (87×38×24.5 mm). Paratypes: MGGC 21929, specimen with partially preserved external features from Sasso delle Streghe; MRSN PU 40611, small specimen from Deruta (41×19×11.5 mm).
Diagnosis
Elongate Archivesica, beak situated anteriorly at ~26% of total shell length; onset of pallial line at posteroventral margin of anterior adductor muscle scar; no pallial sinus.
Occurrence
Middle Miocene seep carbonates in northern Italy.
Description
Elongate veneriform shell (max. dimensions L=95 mm, H=45 mm, W=30 mm) with beak situated in anterior third of shell; anterior margin well rounded, posterodorsal margin only slightly tapering, posterior margin broadly rounded, ventral margin gently rounded, shell surface smooth except for growth lines. Anterior adductor muscle scar broadly oval, bordered posteriorly by distinct ridge; pedal retractor scar oval, distinct from adductor muscle scar; onset of pallial line at posteroventral margin of anterior adductor muscle scar, obliquely cutting across anteroventral shell margin; pallial sinus small, shallow, pointed. RV hinge with thick cardinal 1; cardinal 3a thick, elongate; cardinal 3b thin and very long; LV hinge with cardinal 2a thick, elongate; cardinal 2b thin and close to almost equally shaped cardinal 4b.
Etymology
For the type locality at Sasso delle Streghe (the witches’ rock; Striga Latin for witch).
Materials
Slab with five decalcified shells from Sasso delle Streghe (MGGC 23000); one incomplete articulated mold (MSF 1357); one specimen from Deruta (MRSN PU 40610).
Remarks
Extant species with similar hinge dentition and pallial line include Archivesica gigas, A. kawamurai, A. kilmeri, and A. okutanii (Dall, Reference Dall1895; Kuroda, Reference Kuroda1943; Bernard, Reference Bernard1974; Kojima and Ohta, Reference Kojima and Ohta1997). Among the characters used to distinguish Phreagena from Archivesica was the anterior pedal retractor scar, which is deeply impressed in Phreagena, but shallow in Archivesica (Krylova and Janssen, Reference Krylova and Janssen2006); in A. strigarum it is deeply impressed. Archivesica strigrarum n. sp. is more elongate than Archivesica aharoni n. sp. and has a more oval (rather than round) anterior adductor muscle scar. It lacks the arched posterodorsal margin and posterodorsal ridge of A. apenninica n. sp. and its anterior adductor muscle scar is oval in contrast to the more quadrate scar of A. apenninica n. sp. Compared to the elongate specimens of A. apenninica n. sp., it has a longer anterior portion. It is also more elongate than most other species of Archivesica, except the early Miocene A. sakoi Amano et al., Reference Amano, Jenkins, Ohara and Kiel2014 from southern Japan.
Archivesica apenninica new species

Figure 14 Archivesica apenninica new species, from the middle Miocene seep deposit at Castelvecchio in northern Italy: (1, 2, 10) holotype MGGC 23001, right valve with damaged posterior margin; view of outer shell surface, dorsal view, and close-up of hinge dentition, respectively; (3, 4, 9) paratype MSF 2125, left valve; view of outer shell surface, dorsal view, and close-up of hinge dentition, respectively; (5) paratype MSF 2123, right valve of very elongate specimen; (6) paratype MSF 2121, right valve with damaged posterodorsal margin; (7, 8) paratype MSF 2126, internal mold showing muscle attachment scars and pallial line; view of left valve and dorsal view, respectively.
Type specimens
Holotype: MGGC 23001. Paratypes: MSF 2121, 2123, 2125, 2126; all types from Castelvecchio.
Diagnosis
Large for the genus; dorsal margin broadly arched, ridge running from umbo to posterior margin; anterior adductor muscle scar oval with groove running toward umbo; no pallial sinus; three radiating and almost equally strong cardinal teeth in each valve.
Occurrence
Only known from the middle Miocene (early Serravallian) seep carbonate at Castelvecchio in northern Italy.
Description
Elongate-veneriform shell, up to 125 mm long, umbo broad, elevated, slightly prosogyrate, anterior at ~28–33% total shell length. Anterior shell margin strongly convex; ventral margin gently and evenly curved; posterodorsal margin broadly arched, transition to posterior margin indistinct, angulate only in very elongate specimens; posterior margin strongly convex and somewhat pointed. External sculpture of rough growth lines; distinct ridge or angulation from umbo to posterior margin; no lunula or lunular incision, escutcheon narrow with sharp edges, length at least half of posteroventral margin. Anterior adductor muscle scar distinct, oval, somewhat pointed anteroventrally, most deeply impressed on posterior side, with groove extending toward umbo; anterior pedal retractor scar small, oval, distinct from adductor muscle scar; pallial line starting at posteroventral corner of anterior adductor scar, running oblique toward ventral shell margin in anterior half of shell, parallel to it afterward, sharply bent upward below center of posterior adductor scar, pallial sinus small, shallow. Hinge plate high but short; RV with three equally strong cardinal teeth that radiate toward ventral margin, cardinals 3a and 3b not connected, subumbonal pit present, nymph plate narrow; LV with three widely radiating teeth, cardinal 2a weak, 2b short, wedge-shaped, not connected to 2a, 4 short, nearly perpendicular to 2a.
Etymology
For the Apennine Mountains, Italy.
Materials
All specimens from Castelvecchio; MSF 2121–2132 and several fragmentary specimens at MSF; see Table 7 for measurements.
Table 7 Measurements of Archivesica apenninca new species, all specimens from Castelvecchio; H=height, L=length, W=width of two valves, except when indicated otherwise.

Remarks
Archivesica apenninica n. sp. is variable in elongation, but the characteristic broadly arched posterodorsal margin, as well as the nearly equally strong hinge teeth are seen in all available specimens. A similar broadly arched posterodorsal margin is present in the Recent Japanese species A. solidissima (Okutani, Hashimoto, and Fujikura, Reference Okutani, Hashimoto and Fujikura1992), A. soyoae (Okutani, Reference Okutani1957), and A. okutanii (Kojima and Ohta, Reference Kojima and Ohta1997), but these species differ from A. apenninica either by having a concave ventral margin or by having more anteriorly pointing cardinal teeth (Sasaki et al., Reference Sasaki, Okutani and Fujikura2005). The pallial sinus is more pronounced in extant species than in A. apenninica.
Genus Pliocardia Woodring, Reference Woodring1925
Type species
Anomalocardia bowdeniana Dall, Reference Dall1903, late Pliocene Bowden Formation, Jamaica, by monotypy.
Remarks
Pliocardia is currently a dustbin taxon used for small to moderately sized, oval and thick-shelled species with an Archivesica-like hinge dentition. Molecular phylogenetic studies indicate two separate groups among species with the general shape of Pliocardia: one includes the species Vesicomya stearnsii, Calyptogena ponderosa, C. cordata, and ‘Pliocardia’ krylovata, the other includes Vesicomya kuroshimana, V. crenulomarginata, and specimens called “cf. venusta” (Kojima et al., Reference Kojima, Fujikura and Okutani2004; Martin and Goffredi, Reference Martin and Goffredi2011; Audzijonyte et al., Reference Audzijonyte, Krylova, Sahling and Vrijenhoek2012). Several additional extant species for which no molecular data are available are also included in this genus (Krylova and Sahling, Reference Krylova and Sahling2010), as well as several fossil taxa (Amano and Kiel, Reference Amano and Kiel2007, Reference Amano and Kiel2012). As it is still unclear to which of those groups (if at all) the type species of Pliocardia belongs. The identity of this genus remains unclear and it is only used in quotation marks for the species reported below.
“Pliocardia” italica new species

Figure 15 “Pliocardia” italica new species from the middle Miocene seep deposits at Ca’ Cavalmagra in northern Italy, holotype (MGGC 21909); (1) exterior of left valve; (2) ventral side; (3) anterior side.
Type specimens
Holotype: MGGC 21909 (cast: MSF 1277). Paratype: MSF 1275; all types from the middle Miocene of Ca’ Cavalmagra.
Diagnosis
Large, inflated Pliocardia with strong, protruding, prosogyrate beak, small anterior portion and broadly expanded posterior portion.
Occurrence
Known only from Ca’ Cavalmagra.
Description
Well-inflated specimens with large, protruding beak in anterior third of shell; anterior part of shell short, somewhat pointed, posterior part broadly expanded; anterior adductor muscle scar in ventral half of shell, broadly oval, onset of pallial line at its posteroventral corner, pallial sinus small and shallow, posterior adductor muscle scar indistinct, very close to posterior shell margin; interior of posterior part of shell with distinct radial grooves.
Etymology
Refers to the geographic area that is the source of all studied material.
Materials
One articulated specimen (MSF 1276) and nine isolated valves (MSF 1273, 1274, 1279, 2105–2109; MZB 60218); see Table 8 for measurements.
Table 8 Material of “Pliocardia” italica new species, all specimens from Ca’ Cavalmagra; H=height, L=length, W=width of two valves.

Remarks
Most similar to “Pliocardia” italica n. sp. are two extant species from the central Indo-Pacific Ocean: “Pliocardia” solidissima (Prashad, Reference Prashad1932) and “Pliocardia” ticaonica (Dall, Reference Dall1908). Both have a similarly large and prosogyrate umbo and broad posterior margin, but differ from the Italian Miocene “Pliocardia” italica n. sp. by having a more convex posterodorsal margin; in addition, P. ticaonica is shorter than “Pliocardia” italica n. sp. Among the taxa from the Caribbean Sea and the Gulf of Mexico, “Pliocardia” caribbea (Boss, Reference Boss1967) has a broader anterior margin and has the anterior adductor muscle scar in a more dorsal position; “Pliocardia” cordata (Boss, Reference Boss1968) is shorter and has a smaller and more pointed beak; and “Pliocardia” ponderosa (Boss, Reference Boss1968) seems to have a steeper sloping posterodorsal margin.
Discussion
The present study is intended to provide a taxonomic baseline for future biogeographic and evolutionary studies. At first glance, the bivalve fauna of the ‘Calcari a Lucina’ seep deposits show a wide range of biogeographic links to places as distant as the Recent western Pacific (Bathymodiolus, Archivesica, and Meganodontia), the Recent Indian Ocean (“Pliocardia”), as well as the Miocene Caribbean seep faunas (Meganodontia). We identified 11 chemosymbiotic bivalve species from the middle to late Miocene ‘Calcari a Lucina’ methane-seep deposits in northern Italy, among them three mytilids, one thyasirid, two lucinids, and four or five vesicomyids.
Apart from Bathymodiolus (s.l.) moroniae n. sp. and B. (s.l.) miomediterraneus n. sp. (two species that resemble extant bathymodiolins) the taxonomic affinity of the new genus and species Samiolus iohannesbaptistae is unclear. This taxon might represent a rare case of a genus that occurs exclusively at deep-water methane seeps, but does not belong to the bathymodiolins. An interesting taphonomic aspect of the mytilids is the common occurrence of B. (s.l.) moroniae n. sp. and Samiolus iohannesbaptistae n. gen. n. sp. as disarticulated shells, which is in contrast to the articulated mode of occurrence of the vast majority of other fossil seep-associated bathymodiolins (Squires and Goedert, Reference Squires and Goedert1991; Amano et al., Reference Amano, Jenkins, Aikawa and Nobuhara2010; Kiel et al., Reference Kiel, Campbell and Gaillard2010; Saether et al., Reference Saether, Little, Campbell, Marshall, Collins and Alfaro2010; Amano and Jenkins, Reference Amano and Jenkins2011; Kiel and Amano, Reference Kiel and Amano2013). Articulated shells, including juveniles and giant specimens up to 133 mm in length, indeed occur at Ca’ Piantè, as well as in Deruta, but this is rather an exception than the rule. At the well-sampled sites at Montepetra and Case Rovereti, where disarticulated mussel shells are common, the co-occurring vesicomyids and lucinids were found as articulated or semi-articulated shells. This excludes the possibility that the mussel shells disarticulated due to transport, but suggests in-situ disarticulation, either by the force of the ligament, or perhaps due to large scavengers.
In addition to the seep-inhabiting bathymodiolins reported here, a bathymodiolin named Adipicola apenninica Danise, Bertolaso, and Dominici, Reference Danise, Bertolaso and Dominici2016 was recently described from a whale bone from the middle Miocene (Langhian) Pantano Formation in the vicinity of the town of Carpineti (Reggio Emilia province, northern Italy). Adipicola apenninica was reported as being very abundant, where the associated thyasirids and lucinids were rare (Danise et al., Reference Danise, Bertolaso and Dominici2016).
Most Cretaceous to extant lucinids at methane seeps belong to either the Codakiinae or the Myrteinae (Taylor et al., Reference Taylor, Glover, Smith, Dyal and Williams2011; Kiel, Reference Kiel2013). Thus, if the inclusion of Meganodontia in Pegophyseminae (cf., Taylor et al., Reference Taylor, Glover, Smith, Dyal and Williams2011, Reference Taylor, Glover and Williams2014) is correct, it would be the third lucinid subfamily to have a considerable fossil history at methane seeps (Taylor et al., Reference Taylor, Glover and Williams2014; Kiel and Hansen, Reference Kiel and Hansen2015).
The taxonomy of the Vesicomyidae is still in flux, partially due to the frequent convergence among their few shell characters (Krylova and Sahling, Reference Krylova and Sahling2010). Amano and Kiel (Reference Amano and Kiel2010) recently questioned Paleogene records of Archivesica, and Amano et al. (Reference Amano, Jenkins, Ohara and Kiel2014) identified the early Miocene Japanese A. sakoi Amano et al., Reference Amano, Jenkins, Ohara and Kiel2014 as earliest member of Archivesica sensu strictu. The middle Miocene Archivesica strigarum n. sp. introduced here is thus among the earliest Archivesica species and is, just as A. sakoi, very elongate compared to extant Archivesica species.
Acknowledgments
This paper is a tribute to the late paleontologist M.A. Moroni for her intuition in the ‘60s that the Santa Sofia ‘Calcari a Lucina’ was a peculiar habitat of its own. We are grateful to P. Aharon, S. Conti, G.B. Vai, R. Barbieri, S. Cau, L. Angeletti, G. Bini, S. Gualtieri, for their cooperative work in recognizing and sampling ‘Calcari a Lucina’ limestones and their equivalents along the Italian Apennines over more than 25 years. Above all, we warmly thank M. Sami (Museo Civico di Scienze Naturali, Faenza) for the generous access to the vast collection of fossils from the ‘Calcari a Lucina’ deposits and for comments on various aspects of stratigraphy, and the collaborators of the Museo Civico di Scienze Naturali di Faenza, A. Benericetti, M. Diversi, and V. Liverani, for collecting part of the material. Thanks to D. Ormezzano (Museo Regionale di Scienze Naturali, Torino) for access to, and help with, the collection under his care, and S. Cavagna and G. Pavia for their cooperation. Furthermore, we thank K. Amano (Joetsu) for discussions on vesicomyid systematics; A.G. Beu (Lower Hutt) for advice on the ICZN; E. Krylova (Moscow) for providing images of the subfossil vesicomyid from the Rainbow vent field; C.T.S. Little (Leeds) for donating a specimen from his collection; I. Zorn, M. Harzhauser, and O. Mandic (Vienna) for access to specimens in their care; G. Hundertmark (Göttingen) and J. Bouchal (Stockholm) for photography; and J.D. Taylor (London) for his constructive review of the manuscript. Research on the ‘Calcari a Lucina’ by MT was funded over the years by various projects supported by the Italian Ministry of Education, University and Research (MIUR), and CNR; additional financial support was provided by the Deutsche Forschungsgemeinschaft through grant Ki802/6-1 and the Alméns fund of the Kungliga Vetenskaps-Akademien to SK. This is ISMAR-CNR Bologna scientific contribution n. 1833.
Appendix
Details of localities (see Figure 1) of Miocene methane-seep bivalves reported in this paper. Regarding the localities from Piedmont mentioned here in the context of the Sacco collection at MRSN (Albugnano, Bersano, Pera del Gal , Pino Torinese, etc.), the reader must refer to the catalog edited by Merlino (Reference Merlino2007). The material illustrated by Moroni (Reference Moroni1966) was reported to have been stored in the Museo Geologico Giovanni Capellini, University of Bologna, with the S. Sofia collection (catalog numbers 121–140); however, the collection is not present in this museum and is possibly kept in the University of Palermo, but not traceable at present (G. Ruggieri, personal communication, 1996).
1. Abisso “Mornig”.—Micritic limestone blocks near the entrance of “Mornig” cave, SE of Castelnuovo, not far from Brisighella (Ravenna province), Romagna Apennine, northern Italy; coordinates: 44°13'40''N, 11°44'72''E; Tossignano marls, late Miocene (late Tortonian).
2. Ca’ Carnè.—Marly to micritic limestone deposits just S-SE of Ca’ Carnè (“Vena del Gesso Romagnola” Regional Park visit center), near Brisighella (Ravenna province), Romagna Apennine, northern Italy; coordinates: 44°13'40''N, 11°44'72''E; Tossignano marls, late Miocene (late Tortonian).
3. Ca’ Cavalmagra.—Limestone (calcarenitic) deposit SSE of Palazzuolo (Firenze province), Tuscan Romagna Appenine, northern Italy; coordinates: 44°05'78''N, 11°33'62''E; Marnoso-arenacea Formation, middle Miocene (upper Langhian).
4. Monticino-Limisano – Castelnuovo junction.—Few scattered limestone blocks near the junction between Castelnuovo street and Calbane street, not far from Brisighella (Ravenna province), Romagna Appenine, northern Italy; coordinates 44°14'03.07''N; 11°44'18.72''E; late Miocene (late Tortonian).
5. Ca’ Fornace.—Erratic limestone blocks in the Sintria Creek downstream of the bridge near Ca’ Fornace, near Brisighella (Ravenna province), Romagna Apennine, northern Italy; coordinates: 44°14'14''N, 11°44'40''E; late Miocene (late Tortonian).
6. Ca’ Pianté.—Scattered marly to micritic limestone blocks SW of Ca’ Pianté, not far from Ca’ Carnè and near Brisighella (Ravenna province), Romagna Appenine, northern Italy; late Miocene (late Tortonian); additional information can be found in Conti and Fontana (Reference Conti and Fontana1998).
7. Case Rovereti.—Large isolated limestone block just NW of the small village of Raggio, near Santa Sofia (Forlì province), Romagna Appenine, Italy; coordinates: 43°55'38''N, 11°56'43''E; located on San Paolo Marls but not necessarily belonging to such stratigraphic unit, late Miocene (middle Tortonian–early Messinian). This deposit has previously been described in more detail (Moroni, Reference Moroni1966; Taviani, Reference Taviani1994; Terzi et al., Reference Terzi, Aharon, Ricci Lucchi and Vai1994; Clari et al., Reference Clari, Cavagna, Martire and Hunziker2004a).
8. Castelvecchio.—Limestone (calcarenitic) deposit just W of small village of Castelvecchio, Piancaldoli, near Firenzuola (Firenze province), Tuscan Romagna Apennine, northern Italy; coordinates: 44°11'53''N, 11°24'18''E; Marnoso-arenacea Formation, middle Miocene (early Serravallian).
9. Deruta.—Small blocks at Fosso Castelleone 2 km SE of Deruta (Perugia province), central Italy; coordinates 42°58'15.47''N; 12°26'35.48''E. Upper Miocene (Serravallian). This area was described by Clari et al. (Reference Clari, Conti, Fontana and Taviani2004b).
10. Le Colline.—Scattered marly to micritic limestone blocks outcrop NE of Salecchio, near Palazzuolo (Firenze province), Tuscan Romagna Appenine, northern Italy; coordinates: 44°06'17''N, 11°34'53''E; Marnoso-arenacea Formation, middle Miocene (early Serravallian); further information on this outcrop is provided by Conti and Fontana (Reference Conti and Fontana1998).
11. Marmorito.—Limestone deposits near the village of Marmorito in the Monferrato hills, east of Torino in the Piedmont Basin in northern Italy; coordinates: 45°03'24''N, 08°01'11''E; late Miocene (Tortonian). This well-known deposit was described in more detail elsewhere (Clari et al., Reference Clari, Gagliardi, Governa, Ricci and Zuppi1988; Martire et al., Reference Martire, Natalicchio, Petrea, Cavagna, Clari and Dela Pierre2010).
12. Montepetra.—Large limestone deposit on the southeastern margin of the village of Montepetra (Forlì-Cesena province), northern Italy; coordinates: 43°55'50.67''N, 12°11'38.22''E. (late Tortonian). This deposit is well described in the literature (Taviani, Reference Taviani1994; Conti and Fontana, Reference Conti and Fontana1998; Clari et al., Reference Clari, Conti, Fontana and Taviani2004b; Conti et al., Reference Conti, Fontana, Mecozzi, Panieri and Pini2010).
13. Sasso delle Streghe.—A 30 m high marly limestone peak near the village of Rocca Santa Maria (Modena province), northern Italy; coordinates: 44°28'40.02''N, 10°47'40.11''E; Termina Formation, Miocene (late Serravallian–Tortonian) (Taviani, Reference Taviani1994; Conti and Fontana, Reference Conti and Fontana1998; Taviani, Reference Taviani2001; Clari et al., Reference Clari, Conti, Fontana and Taviani2004b; Conti et al., Reference Conti, Fontana, Mecozzi, Panieri and Pini2010). Also known in the past as ‘Sasso delle Cappe’ (Clams’ rock).
14. Verzino.—Small isolated outcrops in the Crotone basin along the steep right slope of the Vitravo river, between the villages of Verzino and Pallagorio at the localities of Maradera-Occhito (Crotone province), southern Italy; coordinates 39°31'90''N 16°87'32''E to 39°30'97''N 16°87'92''E (late Miocene: Tortonian to Messinian?). More information is reported by Gualtieri (Reference Gualtieri1998).























