



Introduction
Oenonella Fortey, Reference Fortey1980 has previously been known only from the type species, O. paulula Fortey, Reference Fortey1980, from the Middle Ordovician (Dapingian) of Svalbard, arctic Norway. Oenonella paulula was described on the basis of four illustrated cranidia and two pygidia, all of which are reillustrated herein. Peng (Reference Peng1990) recognized that his new genus, Proscharyia, with a type species, P. sinensis, from the Lower Ordovician (upper Tremadocian) of South China, was related to Oenonella. Both Fortey (Reference Fortey1980) and Peng (Reference Peng1990) assigned Oenonella to Scharyiinae Osmólska, Reference Osmólska1957, and this attribution has been followed by most other workers who have commented on either genus (e.g., Owens, Reference Owens1981; Adrain and Fortey, Reference Adrain and Fortey1997; Owens and Fortey, Reference Owens and Fortey2009; Zhou et al., Reference Zhou, Zhou and Yin2016). Scharyiinae has been considered a subfamily of Aulacopleuridae Angelin, Reference Angelin1854 (e.g., Thomas and Owens, Reference Thomas and Owens1978; Fortey, Reference Fortey1980) or of Brachymetopidae (e.g., Owens in Owens and Hammann, Reference Owens and Hammann1990). Following Adrain and Kloc (Reference Adrain and Kloc1997), it has generally been recognized as an independent aulacopleuride family (e.g., Ivanova and Owens, Reference Ivanova, Owens, Kim, Salimova, Kim and Meshchankina2008; Karim, Reference Karim2009; Owens and Fortey, Reference Owens and Fortey2009; Pärnaste et al., Reference Pärnaste, Popp and Owens2009; Hughes and Thomas, Reference Hughes and Thomas2014).
The phylogenetic affinity of Oenonella and Proscharyia has been controversial. Adrain and Chatterton (Reference Adrain and Chatterton1993, table 1) considered Proscharyia to be Aulacopleuroidea incertae sedis. Adrain in Jell and Adrain (Reference Jell and Adrain2003) assigned Proscharyia with question to Bathyuridae Walcott, Reference Walcott1886 and considered Oenonella to be of uncertain affinity. Adrain and Kloc (Reference Adrain and Kloc1997, p. 708–709) noted that new silicified material of species of Oenonella had been recovered from the Middle Ordovician of western Newfoundland, Canada, and that its description might shed light on the affinities of the genus. The primary goals of the present work are the description of two new species of Oenonella from the Middle Ordovician Table Cove Formation of western Newfoundland, together with revision of the type species, and a reconsideration of the relationships of the genus. It is concluded that Oenonella forms a small but well defined clade with a known range from early Tremadocian to Darriwilian, together with Proscharyia and the previously overlooked Amechilus Ross, Reference Ross1951 (which is revised with some new material). The new silicified material seems to effectively rule out a relationship with Scharyiidae, and in fact there is no clear evidence that the group even belongs to the order Aulacopleurida Adrain, Reference Adrain and Zhang2011. Its affinities are at present obscure, but given that there is evidence that a highly distinctive, relatively long-lasting, albeit low diversity, clade is involved, it is appropriate to recognize it as a family-group taxon rather than leaving all three genera “incertae sedis.” The new family Oenonellidae is proposed for their reception.
Localities and stratigraphy
New material is described from the Middle Ordovician of western Newfoundland, Canada, and the Early Ordovician of western Utah, USA.
Table Cove Formation, Newfoundland
The trilobite fauna of the Table Cove Formation was described in a classic monograph by Whittington (Reference Whittington1965) on the basis of calcareous “crack-out” specimens collected on the west coast of Newfoundland's Great Northern Peninsula. Exposures of the formation on the east coast of the peninsula in the Hare Bay region have yielded well preserved secondarily silicified trilobites. Work on these silicified faunas was begun by Adrain et al. (Reference Adrain, Karim and McAdams2020), who proposed a new genus and species of calymenines and detailed the history of study, geological background, and locality information, most of which will not be repeated. Subsequently, Adrain and Pérez-Peris (Reference Adrain and Pérez-Peris2021) described most of the cheirurid taxa in the faunas. The silicified faunas include all the species described from the unit by Whittington (Reference Whittington1965) but are considerably more diverse, with multiple new genera and species. This is almost certainly due mainly to the different preservation, with silicification facilitating the retrieval of types of taxa that are difficult to extract via mechanical preparation. This includes spinose and strongly tuberculate trilobites and, as is the case with Oenonella, very small species. Examples of Oenonella are moderately common at some horizons in our section but were not described by Whittington (Reference Whittington1965).
The rocks of the Table Cove Formation in the study area are bioclastic, thin-bedded, nodular to ribbon limestones interbedded with dark calcareous shale, inferred to have been deposited in a deep subtidal environment, below storm wave base. Their age is mid-Darriwilian (Eoplacognathus suecicus Conodont Zone [Stouge, Reference Stouge1984]; Holmograptus spinosus Graptolite Zone [Maletz et al., Reference Maletz, Egenhoff, Böhme, Asch, Borowski, Höntzsch, Kirsch and Werner2011]). A stratigraphic column with collecting horizons from the section from which material is described, Table Cove Marechal (TCM), is given in Figure 1. The section is on the coast of Hare Bay a few kilometers north of the town of Main Brook. A locality map was given by both Adrain et al. (Reference Adrain, Karim and McAdams2020, fig. 1) and Adrain and Pérez-Peris (Reference Adrain and Pérez-Peris2021, fig. 1).

Figure 1. Stratigraphic column of Table Cove Formation at Section TCM, near Marechal Island, Hare Bay, Great Northern Peninsula, western Newfoundland. Horizons from which material is illustrated are shown. Light colored lithologies are limestones; dark interbeds are calcareous shales. Wavy symbols next to the column indicate beds that are debris flows.
Fillmore Formation, Utah
The specimens of Amechilus palaora Ross, Reference Ross1951, illustrated herein were recovered during an ongoing field-based revision of the silicified Lower and Middle Ordovician trilobite faunas of the Great Basin originally described by Ross (Reference Ross1951) and Hintze (Reference Hintze1953). Comprehensive background, locality maps, and section descriptions were given by Adrain et al. (Reference Adrain, McAdams and Westrop2009, Reference Adrain, Karim, Westrop and Landing2014). The latter work established a new species level trilobite zonation for the mid-Tremadocian Stairsian Stage that is followed herein. New figured material of Amechilus palaora is from section MME 84.0 m, at Middle Mountain, Ibex area, Millard County, western Utah. A simplified graphical log for the section in biostratigraphic context with other Stairsian sections and a detailed meter-scale log were given by Adrain et al. (2014; fig. 4 and appendix 1, respectively).
Materials
Repositories and institutional abbreviations
Material figured or referred to is housed in the Geological Survey of Canada, with specimen number prefix GSC; the Paleontology Repository, Department of Earth and Environmental Sciences, University of Iowa, with specimen number prefix SUI; the Natural History Museum, University of Oslo, with specimen number prefix PMO NF; the Department of Earth Sciences, Natural History Museum, with specimen number prefix NHM It; and Yale Peabody Museum, with specimen number prefix YPM.
Systematic paleontology
Family Oenonellidae new family
Genera included
Oenonella Fortey, Reference Fortey1980 (Floian to Darriwilian, Laurentia affinity, East Svalbard Terrane and Laurentia); Amechilus Ross, Reference Ross1951 (mid-Tremadocian, Laurentia); Proscharyia Peng, Reference Peng1990 (lower to upper Tremadocian, South China).
Diagnosis
Small trilobites with long anterior borders, preglabellar fields, and frontal areas; palpebral lobes very narrow (tr.); glabella long, narrow, and tapering, broadly triangular in outline; S1 usually visible and moderately impressed, running posteromedially toward SO but terminating well anterior to SO to partially define subtriangular L1; S2 and S3 present as subdued, short furrows on internal mold or as small, shallow notches dorsally; posterior fixigenal projections directed mostly laterally, turned posteriorly only at distal portions; cranidium lacking dorsal granular or tuberculate sculpture; pygidium nearly semicircular in outline, with deeply arcuate posterior margin; axis tapered steadily posteriorly, narrow, with five or six axial rings; pleural bands and furrows strongly expressed on entirety of pleural region; rear of pygidial axis set well forward from posterior margin; no pygidial border developed; pleural ribs run directly to margin.
Remarks
When he proposed Oenonella, Fortey (Reference Fortey1980, p. 62) argued that it should be assigned to Scharyiinae on the basis of the “tapering glabella, long preglabellar field, and large, long pygidium.” In addition, Fortey (Reference Fortey1980, p. 63) noted that some species of Scharyia have subdued glabellar furrows, similar to the situation seen in O. paulula. He further argued that while the Katian to Devonian Scharyia Přibyl, Reference Přibyl1946, had been considered by Thomas and Owens (Reference Thomas and Owens1978) to root within the (hence paraphyletic) Upper Ordovician genus Panarchaeogonus Öpik, Reference Öpik1937, Oenonella shared more features with Scharyia than did Panarchaeogonus and seemed “a more plausible alternative” for an ancestor. Similarly, when Peng (Reference Peng1990, p. 108) proposed Proscharyia, he was of the opinion that the “shape and structure of the pygidium, the shape of the glabella, and the nature and the proportions of the preglabellar area of the new genus are almost indistinguishable from those of Scharyia” and that “as far as is known Proscharyia n. g. represents the earliest scharyiine.”
Considering what has been known of the type species—internal molds of cranidia and pygidia for Proscharyia sinensis Peng, Reference Peng1990, and tiny calcareous cranidia and pygidia for Oenonella paulula—this assignment was not unreasonable and was followed by most subsequent authors. As noted in the Introduction, Adrain in Jell and Adrain (Reference Jell and Adrain2003) assigned Proscharyia with question to Bathyuridae and considered Oenonella to be of uncertain affinity. For Proscharyia, this questionable assignment was based on work in progress on a lower Tremadocian group from the Laurentian Skullrockian Stage, but the conclusion was premature, following several new discoveries of species that have yet to be described and formally named. This group is represented in Ross's (Reference Ross1951) monograph by a pygidium (Ross, Reference Ross1951, pl. 29, figs. 4, 6, 7) he tentatively associated with a species of Clelandia Cossman, Reference Cossman1902. This pygidium is broadly similar to those of Proscharyia sinensis. However, it is now clear that such pygidia are associated with cranidia such as those figured by Ross (Reference Ross1951, pl. 14, figs. 1–3) as “Hystricurus ? sp. G.” This group will be treated in detail in a forthcoming work, but the cranidia do not resemble those of Proscharyia, and there is no reason to consider this Laurentian taxon related to it, and there are no grounds for assigning Proscharyia to Bathyuridae.
With the information provided by the new silicified Darriwilian material, the assignment of Oenonella and Proscharyia to Scharyiidae can also be revised. To help comparison and discussion, sclerites of a Sandbian species of an undoubted scharyiid, Panarchaeogonus acris (Hu, Reference Hu1976), are illustrated in Figure 2. The earliest scharyiid is the Darriwilian Lasarchopyge correae Chatterton, Edgecombe, Waisfeld, and Vaccari, Reference Chatterton, Edgecombe, Waisfeld and Vaccari1998 from the Las Aguaditas Formation, San Juan Province, Argentina (Cuyunia), and most of the comparisons apply equally well to that species. Panarchaeogonus acris will be comprehensively revised in a work in progress. As explained by Adrain and Chatterton (Reference Adrain and Chatterton1995, p. 310), when Hu (Reference Hu1976) proposed acris, he misassigned it to the dimeropygid Mesotaphraspis Whittington and Evitt, Reference Whittington and Evitt1954, and misassociated it with pygidia belonging to both the aulacopleurid Harpidella triloba (Hu, Reference Hu1975) (Hu, Reference Hu1976, pl. 27, figs. 27–29) and what is now Strasburgaspis cona (Hu, Reference Hu1971) (Hu, Reference Hu1976, pl. 27, figs. 32, 34–36) (see Adrain, Reference Adrain2005 for revisions of both). Pygidia belonging to Panarchaeogonus acris had previously been misassociated with cranidia and librigenae belonging to Harpidella triloba plus an encrinurid protaspis (Hu, Reference Hu1975, pl. 1, figs. 22, 24–26). The association shown in Figure 2 is correct.

Figure 2. Panarchaeogonus acris (Hu, Reference Hu1976) from the lower part of the Edinburg Formation (Sandbian), Locality 3 of Whittington (Reference Whittington1956, Reference Whittington1959), section in field on south side of road, just east of Strasburg Junction, just west of Strasburg, Shenandoah County, Virginia. (1, 2) Cranidium, SUI 148414, dorsal and ventral views. (3, 4, 7, 8) Pygidium, SUI 148415: (3) dorsal view; (4) ventral view; (7) right lateral view; (8) posterior view. (5, 6) Left librigena, SUI 148416: (5) external view; (6) internal view. Scale bar = 1 mm.
As noted, a relationship with scharyiids was proposed by both Fortey (Reference Fortey1980) and Peng (Reference Peng1990) mainly on the basis of the dimensions of the cranidium and pygidium. The affinities of scharyiids are not in any serious doubt. Their ontogeny has been well documented (e.g., Šnajdr, Reference Šnajdr1981; Chatterton and Speyer, Reference Chatterton, Speyer and Kaesler1997), and they have the suite of adult-like protaspides and juvenile paired glabellar and fixigenal spines characteristic of the Order Aulacopleurida Adrain, Reference Adrain and Zhang2011. A set of cranidia of Oenonella wasisnamei n. sp. covering a range of sizes shows no evidence of paired glabellar or fixigenal spines. These spine pairs are often retained in adults of species of Aulacopleurida, but they are almost invariably reflected on early holaspid and meraspid specimens. Their absence from cranidia of any of the taxa illustrated as Oenonellidae in this paper rules out aulacopleuride affinity.
Cranidia of both Panarchaeogonus (Fig. 2.1) and Scharyia do resemble those of Oenonella and particularly Proscharyia in their subtriangular glabella and more or less extended preglabellar and frontal areas. However, there are numerous significant differences. The scharyiids usually have some form of granular to moderately tuberculate sculpture, whereas the oenonellids are dorsally smooth. The scharyiids often have a tropidial ridge or ridges traversing their frontal area and preglabellar field (Fig. 2.1; Perry and Chatterton, Reference Perry and Chatterton1979, pl. 68, fig. 36). Such a structure is unknown in oenonellids. Scharyiids have a cephalic doublure that exactly underlies the border, with the inner edge aligned with the anterior and lateral border furrows. In Oenonella, the doublure is much broader and transgresses far adaxial to the border furrows, underlying much of the librigenal field. In scharyiids, the rostral plate is short (sag., exsag.) and wide, and in some cases such as Lasarchopyge, the connective sutures are fused. In Oenonella, the inferred shape of the plate is elongate to match the librigenal doublure, flattened, and subtrapezoidal (the plate itself has not been recovered, but its shape is revealed by the intact anterior projections of librigenae of O. wasisnamei; see description of that species). Scharyiids all have relatively large eyes and very long, relatively wide palpebral lobes. The lobes usually have a small pit in the middle of their dorsal area, another typically aulacopleuride feature (Fig. 2.1). Oenonellids have small eyes and concomitantly short (exsag.), very narrow palpebral lobes lacking any evidence of a dorsal pit. Scharyiid pygidia have a true border against which the pleural bands and furrows mostly terminate (Fig. 2.3) and which is underlain exactly by a doublure that is of similar width anteriorly and posteriorly (Fig. 2.4). This contrasts markedly with the condition in Oenonella, in which the doublure is very broad, narrower anteriorly than posteriorly, and underlies much more of the pleural regions, with no true border developed dorsally. In summary, there are few reasons to suppose that oenonellids and scharyiids are related and many morphological features that suggest they are not.
Peng (Reference Peng1990, p. 108) recognized that Proscharyia and Oenonella are related. Comparison is hampered by the fact that Proscharyia sinensis is known only from internal molds of cranidia and pygidia, but cranidia differ only in the more extended and elongate anterior regions of species of Oenonella and the better impressed glabellar furrows of Proscharyia (although that feature is subject to enhancement on internal molds). They otherwise have very similar dimensions, including long, divergent anterior sections of the facial suture, relatively long but very narrow palpebral lobes, and subtriangular glabellae. Pygidia of P. sinensis are wider relative to their length than are those of species of Oenonella, and they feature five versus six segments. However, they show a similar pattern of strong expression of pleural ridges and interpleural furrows across all segments, and just as in Oenonella, these features extend to the pygidial margin, with no true dorsal pygidial border developed.
Amechilus Ross, Reference Ross1951, has been known from a single cranidium assigned to its type species, A. palaora. This cranidium (Ross, Reference Ross1951, pl. 28, fig. 15), from the mid-Tremadocian of the Garden City Formation, southeastern Idaho, has not previously been compared with either Oenonella or Proscharyia, but it has many features in common and is especially comparable to Oenonella paulula, with which it shares almost identical proportions of the extended anterior region, similarly sized and shaped glabella, an eye ridge in the same position and inclination, and narrow palpebral lobes. It differs mainly in having a deeper anterior border furrow, a more robust eye ridge, and somewhat more extended posterior fixigenal projections. Amechilus seems almost certain to be related to Oenonella and Proscharyia, although new field collecting has yielded only two more cranidia (described in the following) and no librigenae or pygidia.
Erben (Reference Erben1961, p. 88, text-fig. 2a) assigned Amechilus to a family Amechilidae, along with Hypothetica Ross, Reference Ross1951. If Amechilus is related to Oenonella and this family is an available name, then obviously it would be the proper name to apply to the group. However, the only other reference to such a family is by Hupé (Reference Hupé1953, fig. 5), who illustrated drawings of both Amechilus and Hypothetica in a text-figure and referred them to “Amechilidae.” Nowhere else in Hupé's work is such a family mentioned, formally proposed, or diagnosed, and the name seems to be a nomen nudum. Later, in the second part of his work, Hupé (Reference Hupé1955, p. 274) instead assigned Amechilus and Hypothetica to a new family, Hypotheticidae, along with Licnocephala Ross, Reference Ross1951, which is now generally considered a bathyurid. This family was listed in the Treatise (Moore, Reference Moore1959, p. O152) and has seen brief mention in the literature (e.g., Černyševa, Reference Černyševa and Orlov1960, p. 178; Balašova, Reference Balašova1967, table 2). It is evidently the name made available for the concept earlier termed “Amechilidae.”
Genus Oenonella Fortey, Reference Fortey1980
Type species
Oenonella paulula Fortey, Reference Fortey1980, from the lowermost Profilbekken Member, Valhallfonna Formation (Dapingian), northern Ny Friesland, Svalbard, arctic Norway (Laurentian-affinity East Svalbard Terrane).
Other species
Oenonella otherfellersorum n. sp., Table Cove Formation (Darriwilian), western Newfoundland, Canada (Laurentia); Proscharyia platylimbata Adrain and Fortey, Reference Adrain and Fortey1997, Tourmakeady Formation (upper Floian), County Mayo, western Ireland (Laurentian-affinity Northwestern Terrane); O. wasisnamei n. sp., Table Cove Formation (Darriwilian), western Newfoundland, Canada (Laurentia); Oenonella sp. 1, Shallow Bay Formation (Floian), western Newfoundland, Canada (Laurentia); Oenonella sp. 2, Antelope Valley Formation (Dapingian), southern Nevada, USA (Laurentia).
Diagnosis
Tiny trilobites with elongate anterior facial sutures; eyes small; dorsal sculpture in large adult specimens of subdued caecal pitting, lacking tubercles of any kind; glabella elongate, subtrapezoidal to subtriangular in outline, with small, independently inflated L1, small notch-like S1, and tiny, ovoid L2; other glabellar furrows and lobes not discernible; pygidia with deeply arcuate outline having six axial rings and pleural segments; pleural furrows weakly expressed or effaced; interpleural furrows bounded anteriorly by distinct transverse ridge developed along the rear of each pleural segment.
Remarks
Oenonella had been known exclusively from its Dapingian type species, revised in the following. New data show it to have a stratigraphic range from Floian to Darriwilian and indicate that Proscharyia platylimbata Adrain and Fortey, Reference Adrain and Fortey1997, is a member of Oenonella. All its species are very small, and the taxon ranks among the trilobites with the smallest known mature size. Given that this has proved true of several species, it seems unlikely to be a matter of sample size or taphonomic bias. At horizons in the Table Cove Formation, Oenonella wasisnamei is fairly common and occurs with thousands of sclerites covering a normal size spectrum for Ordovician trilobites, the far end of which involves species of illaenids, asaphids, and nileids several orders of magnitude larger. Oenonella seems simply to have been tiny.
Oenonella paulula Fortey, Reference Fortey1980
Figure 3.1–3.18
- Reference Fortey1980
Oenonella paulula Fortey, p. 63, text-fig. 7, pl. 11, figs. 1–8, 11.
- Reference Owens1981
Oenonella paulula; Owens, p. 90.
- Reference Owens and Hammann1990
Oenonella paulula; Owens in Owens and Hammann, p. 234.
- Reference Romano and Benton1993
Oenonella paulula; Romano et al., p. 291.
- Reference Jell and Adrain2003
Oenonella paulula; Adrain in Jell and Adrain, p. 413.
Holotype
Cranidium, PMO NF 1324 (Fortey, Reference Fortey1980, pl. 11, figs. 2, 6; Figure 3.3, 3.7, 3.10), from the basal Profilbekken Member, Valhallfonna Formation (Dapingian), Ny Friesland, Svalbard, arctic Norway.

Figure 3. (1–18) Oenonella paulula Fortey, Reference Fortey1980, from the basal Profilbekken Member, Valhallfonna Formation (Dapingian), Ny Friesland, Svalbard, arctic Norway. (1, 2, 5) Cranidium, PMO NF 1327: (1) dorsal view; (2) left lateral view; (5) anterodorsal view. (3, 7, 10) Cranidium, holotype, PMO NF 1324: (3) dorsal view; (7) anterodorsal view; (10) right dorsolateral view. (4, 6, 9) Cranidium, PMO NF 1343: (4) dorsal view; (6) anterior view; (9) right lateral view. (8, 11, 12) Cranidium, PMO NF 1328: (8) dorsal view; (11) left lateral view; (12) anterodorsal view. (13, 14, 17) Pygidium, PMO NF 1325: (13) dorsal view; (14) right lateral view; (17) posterior view. (15, 16, 18) Pygidium, PMO NF 1337: (15) right lateral view; (16) posterior view; (18) dorsal view. (19) Oenonella sp. 1, from talus boulder 8E/8, Shallow Bay Formation (Floian), Back Cove, Cow Head, western Newfoundland, Canada; pygidium, GSC 142626, dorsal view. Scale bars = 0.5 mm.
Diagnosis
Palpebral lobes large, with distinctly impressed palpebral furrow, lobes anteriorly positioned, with rear margin opposite L2; subdued, linear eye ridge running from anterior edge of palpebral lobe to intersection of axial and preglabellar furrows; pygidial axis large, broad, and long, occupying about 80% of sagittal pygidial length, with rear of axis broadly U-shaped; pygidial pleural furrows faintly expressed but discernible, transverse ridges at rear of each pleural segment only very faintly expressed.
Occurrence
Basal Profilbekken Member, Valhallfonna Formation (Dapingian), Ny Friesland, Svalbard, arctic Norway (Laurentian-affinity East Svalbard Terrane).
Materials
In addition to the holotype, assigned specimens PMO NF 1325, 1327, 1328, 1337, 1343.
Remarks
In cranidial features, Oenonella paulula is most similar to Oenonella n. sp. 2, with which it is compared in detail in the following. Among other species, the closest comparison is with O. platylimbata. The Irish species is known from relatively coarsely preserved silicified specimens of a cephalon, cranidia, and pygidia and a calcareous pygidium (Adrain and Fortey, Reference Adrain and Fortey1997, pl. 13, fig. 8, pl. 16, figs. 7–10, 12–16). Two thoracic segments bearing stout medial axial spines (Adrain and Fortey, Reference Adrain and Fortey1997, pl. 15, figs. 3a, b, 4) were very tentatively assigned, but they are far larger than the cephalic and pygidial material and no similar spines have been associated with any other species of oenonellid. It now seems very unlikely that they belong. The cranidium of Oenonella paulula particularly resembles that of O. platylimbata in its nearly parallel-sided glabella that tapers forward only slightly. The species also share a long anterior border, although that of O. platylimbata is longer and separated from the preglabellar field by a much deeper anterior border furrow. Other similarities include a faint but discernible eye ridge and a well impressed palpebral furrow. Cranidia of the species differ conspicuously in the much shorter preglabellar field of O. platylimbata. Comparison of pygidia is hampered by preservation, but that of O. platylimbata appears to have much more strongly inflated pleural bands and more deeply incised pleural and interpleural furrows. Pygidia of the species are similar, however, in overall dimensions, including the possession of a relatively large axis as compared with the overall pygidial length and width.
Of the two new species from the Table Cove Formation, Newfoundland, O. paulula more closely resembles O. wasisnamei. The species share a cranidial anterior border that lacks any dorsal raised line, anterior sections of the facial suture that are only slightly laterally bowed, and an elongate glabella that, in most specimens, is bounded by relatively straight axial furrows that merge into a transverse, anteriorly curved, preglabellar furrow. The structure of an independently inflated L1, notch-like S1, and the subdued L2 are also very similar. As compared with the subtriangular shape of the glabella in O. otherfellersorum, specimens of both O. paulula and O. wasisnamei mostly have a subtrapezoidal glabella with a broad anterior end. There is, however, considerable variation in glabellar shape in O. wasisnamei. A majority of cranidia, including the holotype, have subtrapezoidal glabellae, but the anterior width is unusually variable, and some specimens (e.g., Fig. 4.3, 4.17) are much narrower anteriorly. Cranidia of O. paulula differ from those of O. wasisnamei in the possession of a shorter anterior border, a shorter preglabellar field, the presence of a subdued but readily discernible eye ridge (Fig. 3.1, left side, Fig. 3.3, right side, Fig. 3.4, both sides) versus its complete effacement in O. wasisnamei, considerably larger palpebral lobes with a distinctly impressed palpebral furrow (Fig. 3.3, right side), more anteriorly positioned palpebral lobes, with the rear margin opposite L2 versus opposite the anterior portion of L1 in O. wasisnamei, and broader interocular fixigenae.
Pygidia of the two species are much less similar. In O. paulula, the axis is both broader and much longer, accounting for about 80% of the sagittal pygidial length versus about 64% in O. wasisnamei. The rear of the axis in O. paulula is broadly U-shaped whereas in all other species it is narrow and V-shaped. Transverse pleural furrows are visible on at least the first three segments of the largest pygidium of O. paulula (Fig. 3.13, best seen on right side). Pleural furrows are either extremely faint or completely effaced on pygidia of the other species. The raised ridges at the rear of each pleural segment that are so prominent in O. wasisnamei are present in O. paulula, but they are very subdued (Figure 3.13, best visible in right rear region).
Oenonella wasisnamei new species
Figures 4–6
Holotype
Cranidium, GSC 142627 (Figure 4.1, 4.2, 4.6, 4.7, 4.11), from horizon TCM 18, Table Cove Formation (Darriwilian), near Marechal Island, Hare Bay, western Newfoundland, Canada.

Figure 4. Oenonella wasisnamei n. sp. from horizons TCM 2, TCM 13, and TCM 18, Table Cove Formation (Darriwilian), near Marechal Island, Hare Bay, western Newfoundland, Canada. (1, 2, 6, 7, 11) Cranidium, holotype, GSC 142627: (1) dorsal view; (2) ventral view; (6) left lateral view; (7) anterior view; (11) oblique view (TCM 18). (3, 4, 8, 12) Cranidium, GSC 142578: (3) dorsal view; (4) ventral view; (8) right lateral view; (12) anterior view (TCM 2). (5, 9, 10) Cranidium, GSC 142579: (5) dorsal view; (9) right lateral view; (10) anterior view (TCM 2). (13, 21, 26) Cranidium, GSC 142580: (13) dorsal view; (21) right lateral view; (26) anterior view (TCM 2). (14, 22, 27) Cranidium, GSC 142581: (14) dorsal view; (22) anterior view; (27) right lateral view (TCM 2). (15, 18, 23) Cranidium, GSC 142582: (15) dorsal view; (18) anterior view; (23) right lateral view (TCM 13). (16, 19, 24) Cranidium, GSC 142583: (16) dorsal view; (19) anterior view; (24) right lateral view (TCM 2). (17, 20, 25) Cranidium, GSC 142584: (17) dorsal view; (20) anterior view; (25) right lateral view (TCM 2). (28, 32, 33) Cranidium, GSC 142585: (28) right lateral view; (32) anterior view; (33) dorsal view (TCM 2). (29, 31, 35) Cranidium, GSC 142586,: (29) left lateral view; (31) anterior view; (35) dorsal view (TCM 2). (30, 36, 37) Cranidium, GSC 142587: (30) left lateral view; (36) dorsal view; (37) anterior view (TCM 18). (34, 41, 42) Cranidium, GSC 142588: (34) dorsal view; (41) anterior view; (42) right lateral view (TCM 2). (38–40) Cranidium, GSC 142589: (38) anterior view; (39) dorsal view; (40) left lateral view (TCM 2). Scale bar = 0.5 mm.
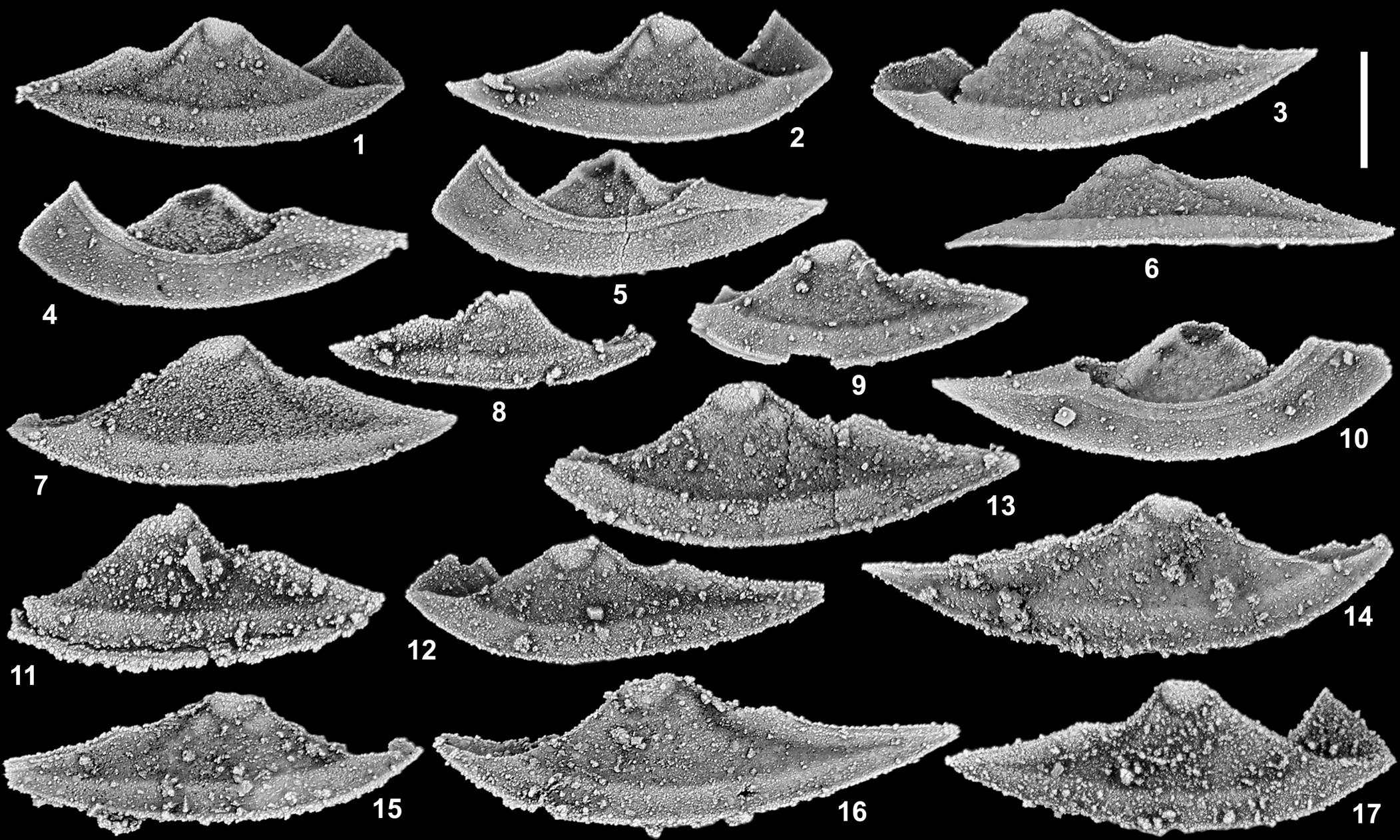
Figure 5. Oenonella wasisnamei n. sp. from horizons TCM 2, TCM 13, and TCM 18, Table Cove Formation (Darriwilian), near Marechal Island, Hare Bay, western Newfoundland, Canada. (1, 4) Right librigena, GSC 142590: (1) external view; (4) internal view (TCM 2). (2, 5) Right librigena, GSC 142591: (2) external view; (5) internal view (TCM 2). (3, 6, 10) Left librigena, GSC 142592: (3) external view; (6) lateral view; (10) internal view (TCM 18). (7) Left librigena, GSC 142593, external view (TCM 2). (8) Right librigena, GSC 142594, external view (TCM 13). (9) Left librigena, GSC 142595, external view (TCM 2). (11) Left librigena, GSC 142596, external view (TCM 13). (12) Left librigena, GSC 142597, external view (TCM 18). (13) Left librigena, GSC 142598, external view (TCM 2). (14) Right librigena, GSC 142599, external view (TCM 18). (15) Right librigena, GSC 142600, external view (TCM 18). (16) Left librigena, GSC 142601, external view (TCM 18). (17) Right librigena, GSC 142602, external view (TCM 2). Scale bar = 0.5 mm.
Diagnosis
Anterior border, anterior sections of facial sutures, preglabellar field, and glabella all very long; palpebral lobes tiny and set posteriorly with rear edge opposite the front of L1; anterior border with single raised line along anterior margin; frontal area and librigenal field with mottled and faint caecal sculpture and pitting; librigena with anterior and posterior small swollen eye socle lobes, posterior border with a single raised line near margin; librigenal doublure with one raised line on inner margin and another near it; pygidium nearly semicircular in outline, with raised ridges at rear of posterior pleural band usually visible on all six pleural segments;
Occurrence
Horizons TCM 2, 13, 18, and BIT 1.2, Table Cove Formation (Darriwilian), near Marechal Island, Hare Bay, western Newfoundland, Canada.
Description
Cranidium elongate, sagittal length 112.7% (102.5%–128.0%) maximum width across posterior fixigenal projections and 166.9% (152.5%–187.6%) width across palpebral lobes; width across maximum point of divergence of anterior sections of facial sutures 107.3% (98.6%–114.5%) width across palpebral lobes; anterior border long (sag.; exsag.), of similar length (sag.; exsag.) in medial region between points where cut anteriorly by anterior sections of facial sutures, shorter (exsag.) laterally in region obliquely cut by suture; border with mostly smooth dorsal aspect, gentle dorsal inflation, subtly swollen posteriorly to form a gentle rim in front of anterior border furrow, with sculpture of a single raised line, subparallel to and set just behind anterior margin; anterior margin describing moderate anterior arc; anterior border furrow very weakly impressed, mainly a break in slope between slightly swollen posterior aspect of border and preglabellar field, anteriorly arcuate, subparallel to anterior margin of border; preglabellar field elongate, sagittal length 31.4% (28.8%–37.3%) sagittal length of cranidium, field and frontal areas slightly dorsally concave, with faint caecal sculpture and pitting visible in some specimens (Fig. 4.1, 4.5); anterior sections of facial sutures slightly divergent in front of palpebral lobes, nearly subparallel in some specimens (e.g., Fig. 4.16), with course varying from nearly straight to more commonly slightly laterally bowed (e.g., Fig. 4.5); interocular fixigena lacking sculpture, nearly flat, with only slight transverse dorsal inflation (e.g., Fig. 4.7, 4.19, 4.20), about twice as wide as palpebral lobe, more strongly inflated posteriorly in front of posterior border furrow (best seen ventrally, e.g., Fig. 4.2); palpebral lobe slightly shorter (exsag.) than occipital ring (sag.), very narrow, lateral margin describing shallow lateral arc, elevated at a slight angle from interocular fixigena (Fig. 4.7); palpebral furrow only impressed posteriorly near rear of lobe, where it is shallow (e.g., Fig. 4.1, 4.5), over anterior course only present as a break in slope from the interocular fixigena; posterior fixigenal projection relatively narrow, with slim triangular strip of fixigena between posterior section of facial suture and posterior border furrow, posterior suture set 40°–50° from transverse behind palpebral lobe, deflected strongly posteriorly opposite anterior edge of posterior border furrow; posterior border furrow relatively shallow but distinctly impressed proximally, adaxial to fulcrum, shallower behind interocular fixigena and extended distally as only a very shallow impression (e.g., Fig. 4.17, 4.36) or entirely effaced distally (e.g., Fig. 4.1, 4.5); posterior border distinct and dorsally convex proximally, where border furrow is defined, but flattened distally and partially or wholly merged with fixigena; posterior margin of posterior border nearly transverse, only slightly posterolaterally directed adaxial to fulcrum, turned strongly posteriorly distally, lateral extent forming sharp posteriorly directed point; fulcrum fairly steeply declined, distal posterior fixigenal projection set about 50° from horizontal; glabella excluding LO with maximum width across base, 72.7% (69.1%–77.1%) sagittal length; axial furrows nearly straight, with only slight deflection around L1 and L2, anteriorly convergent, narrow and firmly inscribed, slightly wider posteriorly; preglabellar furrow with more or less even anterior arc, varying in width from moderate to narrow, so that glabella in plan view ranges from subtrapezoidal (e.g., Fig. 4.1, 4.5, 4.16) to subtriangular (e.g., Fig. 4.3, 4.17, 4.39) depending upon its anterior width; preglabellar furrow of similar depth and width to anterior part of axial furrow; S1 visible on all specimens as a slight lateral notch, with a distinct but shallow furrow running posteromedially for a short distance, to partially outline a small but independently inflated L1; S2 visible only as a faint lateral indentation, L2 weakly defined but with slight independent inflation, around which the axial furrow slightly deflects in most specimens; S3 and L3 not obviously defined; SO transverse, short (sag.; exsag.), firmly impressed; LO very slightly longer sagittally than exsagittally, sagittal length 12.1% (10.1%–13.2%) cranidial sagittal length, posterior margin describing gentle posterior arc; very faint median node present at about half midlength of occipital ring in some specimens (e.g., Fig. 4.1, 4.5, 4.13), obscure in others (e.g., Fig. 4.3, 4.14, 4.16, 4.33); doublure forming articulating surface under LO (Fig. 4.4), featureless, anterior margin with shallow anterior arc, slightly ventrally concave; doublure beneath posterior fixigena projection limited to a very short (exsag.) strip along rear, slightly longer distally.
Librigena with maximum width at rear of eye, 31.9% (29.3%–34.3%) length of lateral margin; lateral border broad, 28.0% (25.9%–30.5%) maximum librigenal width, of similar width anteriorly and posteriorly, lacking sculpture, with slight dorsal inflation (Fig. 5.6), extended posteriorly to form abaxial part of genal spine, very slightly narrowed in this region; posterior facial suture with slight, narrow sutural ridge, turned posteriorly to cut posterior border at about one-third lateral distance on field; posterior border slightly narrower than lateral border, with a single raised line near margin, running subparallel with margin; posterior border broad near facial suture, progressively more narrow posteriorly, where it forms the adaxial portion of the genal spine; eye small, narrow; eye socle of two weakly inflated anterior and posterior lobes, posterior larger and slightly more inflated; between lobes, visual surface separated from field by a weak furrow; field with mottled sculpture and weak caecal pitting, extended posteriorly onto anterior part of genal spine, pinched out as narrow furrow running posteriorly to union of posterior and lateral borders; genal spine broad, subtriangular, with slight posterior curvature and sharp distal tip; anterior facial suture nearly straight opposite field, deflected anterior as it cuts across lateral border; doublure broad and flat, outer part slightly ventrally concave and lacking sculpture; inner portion with elongate ridge along inner edge, of similar width anteriorly, broader posteriorly underneath anterior part of genal spine, tapered posteriorly under adaxial edge of distal portion of genal spine; ridge separated from abaxial flattened part of doublure by narrow furrow; single prominent raised line set on ridge at adaxial margin (best seen on Fig. 5.10); inner edge of doublure set adaxial to lateral border furrow, underlying part of field; connective suture nearly straight, with slight lateral change in course abaxially near contact with anterior section of facial suture.
Rostral plate not found, but intact anterior doublural projections (Fig. 5.1, 5.5) show that it was large, flat, and trapezoidal, broader anteriorly than posteriorly.
Hypostome and thorax not found.
Pygidium with maximum width across first segment, 143.0% (129.0%–171.0%) sagittal length; axis with maximum anterior width 63.8% (58.2%–69.2%) sagittal length and sagittal length 66.4% (62.4%–75.3%) sagittal length of pygidium; pygidium approximately semicircular in plan view, arc of margin becoming somewhat more pronounced posteriorly; pygidium with strong dorsal vaulting, distal pleural regions steeply turned down around fulcrum, axis dorsally produced in transverse view; bottom margin of pygidium completely flat in both transverse and posterior views; pygidium composed of six segments; axis with six axial rings, all fully visible in some specimens (Fig. 6.1, 6.12), posterior rings effaced in others (Fig. 6.5, 6.31, 6.33); articulating half-ring very large, ellipsoid in shape, slightly more than twice as long sagittally as first axial ring; first ring longer (exsag.) near axial furrow than sagittally; all rings short (sag., exsag.), lacking sculpture, relatively weakly inflated; ring furrows shallow but complete in most specimens; axial furrows posteriorly convergent, ranging from nearly straight (Fig. 6.5, 6.10) to slightly laterally bowed (Fig. 6.1), meeting posteriorly to fully circumscribe rounded rear of axis; proximal portions of pleurae flat; portions distal to pleurae steeply inclined, in some specimens nearly vertical (e.g., Fig. 6.28, 6.29), ranging from slightly outwardly convex (Fig. 6.9) to slightly outwardly concave (Fig. 6.6); all six pleural segments expressed on most specimens; first segment with very faint pleural furrow proximally, anterior and posterior pleural bands of about the same length (exsag.); segment longer (exsag.) distal to fulcrum, rear margin bowed slightly posteriorly; rear of posterior pleural band inflated into prominent ridge, which runs from axial furrow all the way to the lateral margin, slightly posteriorly overhanging first interpleural furrow; subsequent posterior segments with very similar morphology; first four segments nearly completely expressed, fifth and sixth marked mainly by expression of posterior ridge, sixth segment nearly effaced in some specimens (Fig. 6.5, 6.31); posteromedian region of pleurae behind tiny axial terminal piece smooth; no distinct border developed, but adaxial extent of doublure is reflected dorsally on some specimens as a subtly raised arc more or less around the fulcrum (e.g., Fig. 6.12); scattered and wavy raised line sculpture is developed across the entire length of the first segment around the fulcrum on most specimens (best seen on Fig. 6.1)—where it appears absent it may be a matter of indifferent preservation; similar sculpture is present but more weakly expressed on the second segment in some specimens (Fig. 6.12, 6.31 left side); doublure broad, narrower anteriorly, and becoming progressively broad posterior, longest sagittally, with slight ventral concavity, set more or less plane-parallel to steeply inclined distal dorsal regions of pleurae, thus steeply inclined from the plane of the base of the pygidial margin (Fig. 6.7); sculpture of three fairly weak raised lines on anterior half of surface area, running subparallel to anterior margin (Fig. 6.2).

Figure 6. Oenonella wasisnamei n. sp. from horizons TCM 2, TCM 13, TCM 18, and BIT 1.2, Table Cove Formation (Darriwilian), near Marechal Island, Hare Bay, western Newfoundland, Canada. (1–3, 6, 7) Pygidium, GSC 142603,: (1) dorsal view; (2) ventral view; (3) right lateral view; (6) posterior view; (7) anterior view (TCM 18). (4, 5, 9, 13) Pygidium, GSC 142604: (4) right lateral view; (5) dorsal view; (9) posterior view; (13) ventral view (TCM 2). (8, 12, 16) Pygidium, GSC 142605: (8) right lateral view; (12) dorsal view; (16) posterior view (TCM 18). (10, 14, 17) Pygidium, GSC 142606: (10) dorsal view; (14) posterior view; (17) right lateral view (TCM 2). (11, 15, 18) Pygidium, GSC 142607: (11) dorsal view; (15) posterior view; (18) left lateral view (TCM 18). (19, 21, 26) Pygidium, GSC 142608: (19) dorsal view; (21) right lateral view; (26) posterior view (TCM 18). (20, 22, 28) Pygidium, GSC 142609: (20) right lateral view; (22) dorsal view; (28) posterior view (TCM 13). (23, 24, 29) Pygidium, GSC 142610: (23) dorsal view; (24) left lateral view; (29) posterior view (TCM 18). (25, 30, 31) Pygidium, GSC 142611: (25) left lateral, view; (30) posterior view; (31) dorsal view (TCM 2). (27, 32, 33) Pygidium, GSC 142612: (27) left lateral view; (32) posterior view; (33) dorsal view (BIT 1.2). Scale bars = 0.25 mm.
Etymology
For Buddy Wasisname.
Materials
In addition to the holotype, assigned specimens GSC 142578–142612.
Remarks
While there are two species of Oenonella occurring together at some horizons in the Table Cove Formation, there is no question of misassociation due to both strong morphological correspondence of exoskeletal parts and relative abundance of the species. In particular, O. wasisnamei occurs to the exclusion of O. otherfellersorum at horizon TCM 18, which confirms beyond any doubt the correct association of cranidia, librigenae, and pygidia.
Oenonella wasisnamei was compared with the type species, O. paulula, in the preceding and is compared with O. otherfellersorum under remarks on that species in the following. Its closest morphologic comparison is with Oenonella sp. from the Floian of the Shallow Bay Formation, which is known only from a single pygidium (Fig. 3.19). The species share the prominent expression of the raised transverse ridges along the rear of each pygidial pleural segment. They differ in that the course of these ridges in specimens of O. wasisnamei is slightly posteriorly bowed in the distal region abaxial to the fulcrum and atop the doublure, and they are sharply deflected at the fulcrum. In Oenonella sp. the ridges are evenly arcuate, with no abrupt fulcral deflection and no posterior bowing in their distal regions. In addition, the sixth pair of ridges are separate in specimens of O. wasisnamei but are merged into a single post-axial ridge in Oenonella sp.
Oenonella otherfellersorum new species
Figure 7
Holotype
Cranidium, GSC 142613 (Fig. 7.1, 7.2, 7.7, 7.11), from horizon TCM 14*, Table Cove Formation (Darriwilian), near Marechal Island, Hare Bay, western Newfoundland, Canada.
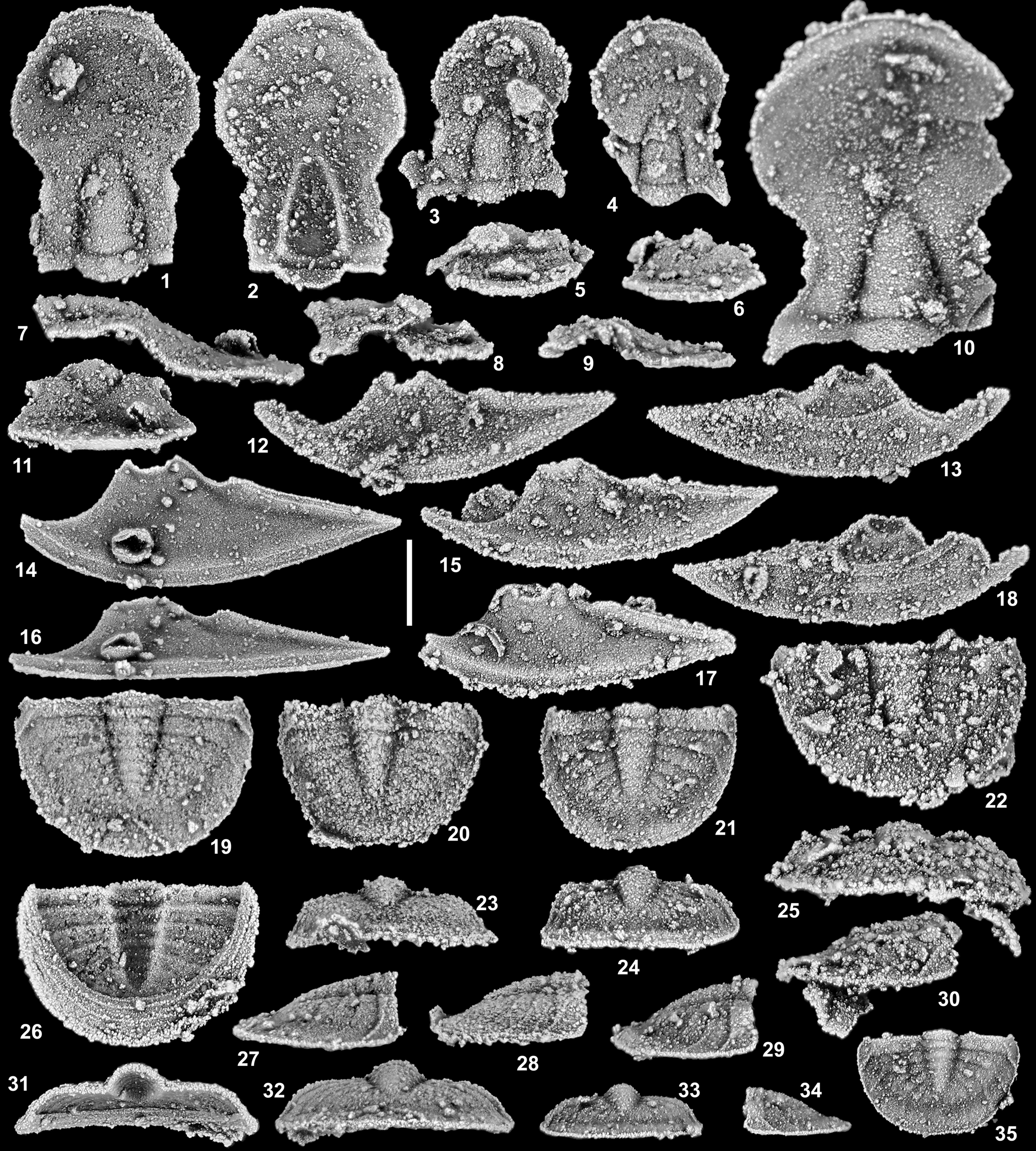
Figure 7. Oenonella otherfellersorum n. sp. from horizons TCM 2, TCM 13, and TCM 14*, Table Cove Formation (Darriwilian), near Marechal Island, Hare Bay, western Newfoundland, Canada. (1, 2, 7, 11) Cranidium, holotype, GSC 142613: (1) dorsal view; (2) ventral view; (7) right lateral view; (11) anterior view (TCM 14*). (3, 5, 8) Cranidium, GSC 142614: (3) dorsal view; (5) anterior view; (8) right lateral view (TCM 14*). (4, 6, 9) Cranidium, GSC 142615: (4) dorsal view; (6) anterior view; (9) right lateral view (TCM 13). (10) Cranidium, GSC 142616, dorsal view (TCM 13). (12, 13) Left librigena, GSC 142617: (12) external view; (13) internal view (TCM 14*). (14, 16) Left librigena, GSC 142618: (14) external view; (16) lateral view (TCM 13). (15, 18) Left librigena, GSC 142619: (15) external view; (18) internal view (TCM 14*). (17) Left librigena, GSC 142620, external view (TCM 14*). (19, 26, 27, 31, 32) Pygidium, GSC 142621: (19) dorsal view; (26) ventral view; (27) right lateral view; (31) anterior view; (32) posterior view (TCM 13). (20, 23, 28) Pygidium, GSC 142622: (20) dorsal view; (23) posterior view; (28) right lateral view (TCM 2). (21, 24, 29) Pygidium, GSC 142623: (21) dorsal view; (24) posterior view; (29) right lateral view (TCM 13). (22, 25, 30) Pygidium, GSC 142624: (22) dorsal view; (25) posterior view; (30) right lateral view (TCM 14*). (33–35) Pygidium, GSC 142625: (33) posterior view; (34) left lateral view; (35) dorsal view (TCM 13). Scale bar = 0.5 mm.
Diagnosis
Preglabellar field and frontal areas very elongate; anterior border short (sag., exsag.), with raised line at anterior margin and second smaller raised line near posterior margin; anterior sections of facial sutures strongly laterally bowed; palpebral furrow shallow but fully expressed; glabella small and narrow; librigena with raised line on lateral margin of lateral border and posterior margin of posterior border, and second raised line near lateral border furrow and at about half width of posterior border; pygidium with margins nearly straight and subparallel anteriorly, rounded and evenly arcuate posteriorly; transverse ridges at rear of posterior pygidial pleural bands relatively subdued on second and more posterior segments; distal pleural regions of pygidium above doublure distinctly outwardly concave.
Occurrence
Horizons TCM 2, 13, and 14*, Table Cove Formation (Darriwilian), near Marechal Island, Hare Bay, western Newfoundland, Canada.
Description
Cranidium very elongate, width across maximum divergence of anterior sections of facial sutures 71.5% (67.6%–74.8%) sagittal length and 123.2% (116.4%–132.0%) width across midpoint of palpebral lobes; anterior margin of anterior border cut by anterior section of facial suture at a position just adaxial to anterior edge of palpebral lobe, describing anteriorly directed arc; anterior border relatively short (sag., exsag.), nearly flat dorsally, with sculpture of one raised line running along anterior margin and a second, finer, raised line just in front of posterior margin, both running subparallel to anterior arc of margin; anterior border furrow very shallow, mainly marking a slight change in slope at rear of border; anterior sections of facial sutures very long (exsag.), strongly laterally bowed, parentheses-like; preglabellar field exceptionally long, sagittal length 40.6% (38.2%–43.6%) that of cranidium; preglabellar field and frontal area broad, slightly dorsally concave, lacking sculpture; interocular fixigena gradational with frontal area, width 19.2% (14.6%–21.3%) width of cranidium across midlength of palpebral lobes, with slight dorsal convexity, lacking sculpture; narrow and shallow palpebral furrow fully expressed along length of palpebral lobe; palpebral lobe confluent with slight sutural ridges both anteriorly on rear part of anterior section of facial suture and posteriorly on anterior part of posterior section of facial suture, about as long (exsag.) as sagittal length of LO, very narrow, held almost horizontally (Fig. 7.11); posterior fixigena dorsally somewhat inflated adaxially along rear part of axial furrow (see particularly Fig. 7.10, left side), lacking sculpture; posterior fixigenal projections narrow, extended laterally only about as far as lateral extent of anterior sections of facial suture, or slightly less; fixigena forming small triangular region on posterior projection, slightly dorsally concave, lacking sculpture; posterior border furrow short (exsag.), shallow, but impressed adaxial to fulcrum; very shallow to obsolete distal to fulcrum; posterior border with slight dorsal inflation, proximally slightly shorter (exsag.) than LO, distal to fulcrum becoming elongate, to form sharp nearly posteriorly directed tip where cut by posterior section of facial suture, lacking sculpture; glabella short, weakly inflated, and narrow, sagittal length excluding LO 36.2% (34.9%–37.2%) that of cranidium, maximum width across L1 63.0% (62.0%–64.3%) sagittal length; axial furrows deep, slightly shallower and narrower anteriorly versus posteriorly, in smaller specimens (Fig. 7.1, 7.3, 7.4) subparallel posterior and convergent anterior, in single larger specimen (Fig. 7.10) more or less convergent along entire length; shape of glabella consequently varies from of even width posteriorly with triangular anterior region to subtriangular in plan view (Fig. 7.10); L1 about as long as LO, with slight lateral inflation; S1 expressed only as a shallow lateral depression; L2 with very slight independent lateral inflation; S2 and anterior lobes/furrows not obviously expressed; glabella lacking dorsal sculpture; SO transverse, very short (sag., exsag.), firmly impressed; LO longest sagittally, significantly shorter (exsag.) near axial furrow, lacking dorsal sculpture, with very faint median node set on anterior half (Fig. 7.10); articulating surface of doublure beneath LO with transverse anterior margin (Fig. 7.2), slightly ventrally concave, lacking sculpture; doublure forming a narrow strip beneath posterior margin of posterior border.
Rostral plate, hypostome, and thorax not found.
Librigena with width at midlength of eye, excluding visual surface, 27.2% (25.2%–30.4%) length from contact of anterior facial suture with adaxial edge of lateral border to tip of genal spine; eye narrow but relatively elongate; socle of a single faint posterior swollen region (best seen on Fig. 7.17), visual surface separated from field by shallow furrow; field smooth, completely lacking sculpture, extended posteriorly in elongate triangle to tip of genal spine; lateral border furrow very shallow, reflecting only gradational change in slope from field to border; posterior border furrow shallow but clearly expressed from near eye to along adaxial part of genal spine, separated from posterior section of facial suture near eye by small sutural ridge; posterior section of facial suture short, cutting posterior border posteriorly just abaxial to bottom of eye; posterior border flat and narrow, with prominent raised line running entire length at about midwidth and a second raised line running along the entire adaxial margin; border and lines extended distally to tip of genal spine; lateral border wider than posterior border, with similar raised line along length, but set near to adaxial margin and a second raised line along margin; lateral border and raised lines extended posterior to tip of genal spine; adaxial margin of posterior border nearly straight; lateral margin of lateral border strongly laterally bowed; doublure very broad, underlying lateral border, entirely of genal spine, and most of the area of the field, with sculpture of a raised line along adaxial margin and two more raised lines on ventral aspect, curved to run subparallel to inner margin (Fig. 7.13, 7.18), doublure with very slight ventral concavity.
Pygidium with maximum width across first segment, 137.7% (133.7%–142.2%) sagittal length; pygidium composed of six segments; axis with maximum anterior width 25.0% (23.5%–27.7%) overall pygidial width and 52.2% (47.0%–61.0%) sagittal axial length; sagittal length of axis 66.2% (64.3%–69.6%) length of pygidium; anterior margin of pleural regions of pygidium nearly transversely straight; axial furrows narrow, well impressed over much of their length, distinctly shallower and more weakly impressed in posterior region, but still meeting posteriorly to fully define rear of axis; ring furrows all short (sag., exsag.), shallow, transversely straight, but distinctly impressed; articulating half-ring large, subequal in sagittal length (Fig. 7.19) to slightly longer (Fig. 7.21, 7.35) than first ring; six axial rings of similar length sagittally and exsagittally, progressively narrower posteriorly, each with independent inflation though somewhat effaced in some specimens (e.g., Fig. 7.21); axis with slight to moderate dorsal vaulting above surrounding proximal pleural regions (Fig. 7.23, 7.24); fulcrum set about two-thirds distance across pleural region, areas distal to fulcrum strongly downturned (Fig. 7.23, 7.24); pleural furrow extremely weakly impressed on first and second segment, dividing segment into anterior and posterior pleural bands of subequal length, pleural furrow only intermittently visible on posterior segments of some specimens (e.g., Fig. 7.21); linear ridge developed along rear of posterior pleural band, mainly in distal region, where it is turned sharply posterior and slightly overhangs interpleural furrow; ridges obvious on anterior three segments, more variably and weakly expressed posteriorly; interpleural furrows short (exsag.), well impressed, first furrow transverse, posterior furrows turned increasingly posteriorly; all six segments with pleural bands and interpleural furrows expressed; pleural features extended uninterrupted to margin, marked most prominently by posterior pleural ridges of first three segments; no true border developed, but there is a distinct, flattened, dorsally concave region surrounding the pygidial margin where the pleural structures, while extending to the margin, are somewhat effaced compared with higher on the pygidium; doublure relatively narrow anteriorly, progressively broadened posteriorly, inner margin semicircular, with sculpture of three or four more or less evenly spaced raised lines set subparallel to inner and outer margins; doublure sagittally reaching nearly to rear of axis.
Etymology
For the Other Fellers.
Materials
In addition to the holotype, GSC 142614–142625.
Remarks
Oenonella otherfellersorum differs from O. wasisnamei, with which it occurs at some horizons, in a cranidial anterior border with raised lines both at the anterior margin and in front of the posterior margin versus at the anterior margin only. The anterior border is significantly shorter, and the anterior sections of the facial sutures are much more laterally bowed. The preglabellar field and frontal area show no sign of the subdued caecal sculpture and pitting characteristic of this region in O. wasisnamei. The glabella is relatively smaller, and posteriorly is more parallel-sided in most specimens, versus anteriorly tapering along its entire length. The librigena lacks caecal sculpture and pitting on the field, has only a posterior swollen socle lobe, and has narrower posterior and lateral borders. The librigenal lateral border has a prominent raised line on its adaxial part that is not found in O. wasisnamei. The librigenal doublure is somewhat broader and has three, versus two, subparallel raised lines. The pygidium is more parallel-sided anteriorly versus generally semicircular in outline. The pygidial axis is slightly narrower and the axial rings more subject to partial effacement. The raised ridges along the posterior pygidial pleural bands are less prominent, particularly posteriorly; the outwardly concave distal part of the pleural region is more prominent. Finally, the pygidial doublure is not as long sagittally but bears more-prominent raised lines.
Oenonella otherfellersorum differs from both O. paulula and O. platylimbata in having much more laterally bowed anterior sections of the facial sutures. The anterior border is shorter and has a posteriorly set raised line. The glabella is relatively much smaller and the preglabellar field much longer. The interocular fixigena is narrower, and there is no sign of the faint eye ridge seen in O. paulula. L1 and L2 are not as strongly inflated, and S1 is much less prominent. The pygidial axis is far smaller and the pleural regions concomitantly larger.
Oenonella otherfellersorum differs from Oenonella sp. 1 from the Shallow Bay Formation in having subdued versus robust transverse ridges along the rear of the pygidial pleural segments. The ridges are much more transverse in O. otherfellersorum, deflected sharply posterolaterally only near the pygidial lateral margin. They are nearly evenly bowed posteriorly in Oenonella sp. In addition, the axis comes to a narrower posterior point in O. otherfellersorum, and it is comparatively somewhat longer (sag.). The pleural region in O. otherfellersorum is slightly dorsally concave in the area atop the doublure, whereas it is dorsally convex in this area in Oenonella sp.
Oenonella sp. 1
Figure 3.19
Occurrence
Talus boulder 8E/8, Shallow Bay Formation (Floian), Back Cove, Cow Head, western Newfoundland, Canada.
Materials
Assigned specimen GSC 142626.
Remarks
A single specimen from the conglomerates of the Shallow Bay Formation in western Newfoundland shows a subsemicircular pygidium, six axial rings, and raised ridges at the rear of the posterior pleural bands extended to the pygidial margin, all typical of Oenonella. The specimen was compared with younger species in the preceding. Its main unique feature is that while the anterior five pleural segments feature paired posterior ridges, those of the sixth are apparently fused into a single posteromedian ridge, although the tiny sixth axial ring is well developed.
Oenonella sp. 2
Figure 8
- Reference Fortey1980
Oenonella sp.; Fortey, p. 63.

Figure 8. Oenonella sp. 2 from the Antelope Valley Formation (Dapingian), flanking beds of Meiklejohn bioherm, Meiklejohn Peak, Nye County, southern Nevada, USA. (1–4) Cranidium, NHM It 9810: (1) dorsal view; (2) oblique view; (3) left dorsolateral view; (4) anterodorsal view. Scale bar = 0.5 mm.
Occurrence
Flanking beds of Meiklejohn bioherm (Ross, Reference Ross1972), Antelope Valley Formation (Dapingian), Meiklejohn Peak, Nye County, southern Nevada, USA.
Materials
Assigned specimen NHM It 9810.
Remarks
A single cranidium from the Dapingian of southern Nevada most closely resembles those of the other known Dapingian species, O. paulula. The species share comparable cranidial dimensions, including the length of the preglabellar field, the degree of anterior divergence of the anterior sections of the facial sutures, the substantial width of the interocular fixigenae, and the length and width of the glabella. The specimen has a fairly prominent median node on LO (preserved on the internal mold) that seems slightly larger than that of O. paulula, but both species have more-prominent nodes than either of the new Darriwilian taxa. Also comparable between the Nevada specimen and O. paulula is the marked subspherical inflation of L1. An eye ridge is partially preserved on the right side of the specimen, although it is obscured near the glabella, and this seems somewhat more prominent than in O. paulula. The right side also partially preserves a palpebral furrow that seems narrower and more linear than in O. paulula (e.g., Fig. 3.3). The Nevada species differs from all others in the extended length of its anterior border and the presence of prominent caecal pits aligned in an arc along its anterior border furrow.
Genus Amechilus Ross, Reference Ross1951
Type species
Amechilus palaora Ross, Reference Ross1951, from the Garden City Formation (mid-Tremadocian; Stairsian), southeastern Idaho, USA.
Other species
Hyperbolochilus ? forteyi Peng, Reference Peng1990, Shallow Bay Formation (lower Tremadocian), western Newfoundland, Canada (Laurentia).
Diagnosis
Anterior sections of facial suture with moderate lateral arc; anterior border furrow shallow but independently impressed (as opposed to a simple break in slope between anterior border and preglabellar field); glabella relatively short, subtrapezoidal; palpebral lobes narrow but elongate; posterior fixigenal projections short (exsag.), longer abaxially, very wide, running slightly posterolaterally.
Remarks
Amechilus has been known only from a single specimen, the unique holotype of its type species. Kobayashi (Reference Kobayashi1955, p. 459) proposed Amechilus tuberculatus from the Lower Ordovician of the McKay Group in southeastern British Columbia, Canada. Dean (Reference Dean1988, p. 24) considered this species to be a junior subjective synonym of the hillyardinine Metabowmania latilimbata Kobayashi, Reference Kobayashi1955. Adrain and Westrop (Reference Adrain and Westrop2007, p. 233) tentatively accepted this but pointed out that there were numerous apparent morphological differences and that new collections from the McKay Group were likely necessary to meaningfully evaluate the problem. There is no question, however, that the unique holotype of A. tuberculatus is a hillyardinine, unrelated to Amechilus palaora.
Fortey (Reference Fortey, Briggs and Lane1983, p. 193, pl. 25, figs. 10, 11) illustrated a single cranidium from the “Symphysurina Zone” of the Shallow Bay Formation at Broom Point, western Newfoundland, as “Hyperbolochilus? sp. nov.” Hyperbolochilus Ross, Reference Ross1951 is a hillyardinine (see., e.g., Adrain et al., Reference Adrain, Karim, Westrop and Landing2014, fig. 13C, D, G, H, fig. 15O, S) restricted to the younger mid-Tremadocian Stairsian Stage. Fortey's comparison was reasonable, especially as all that was known then of Hyperbolochilus was a few small photographs published by Ross (Reference Ross1951) and Hintze (Reference Hintze1953). Work in progress on hillyardinines, including multiple species of Hyperbolochilus, indicates that none is similar in detail to the Shallow Bay Formation specimen. In particular, the latter shows an elongate preglabellar field and expansive frontal area and a glabella that is relatively small and tapers strongly forward. Species of Hyperbolochilus have shorter preglabellar fields, generally less-divergent anterior facial sutures, and relatively larger glabellae with nearly parallel sides.
When he proposed Proscharyia, Peng (Reference Peng1990) assigned Fortey's specimen to his new taxon. Curiously, he listed it in his “Other species” section as “Hyperbolochilus ? forteyi.” In his remarks on the type species of Proscharyia, P. sinensis, Peng (Reference Peng1990, p. 109) said “The close similarity of Hyperbolochilus ? forteyi, described originally as H.? sp. nov., …” The form “Hyperbolochilus ? forteyi” is Peng's own invention, and apparently he sought to name a new species for Fortey's (Reference Fortey, Briggs and Lane1983) single specimen. He did so, however, without any formal procedure. He did not explain that he was naming a new species, did not explain the basis for the name (evidently for Richard Fortey), did not designate a holotype, and gave no formal diagnosis. There is hence some question whether the name is available under the International Code of Zoological Nomenclature. It does appear that it is. The fourth edition of the Code (International Commission on Zoological Nomenclature, 1999) explicitly required designation of a holotype, but this was not in effect at the time of Peng's publication. As Fortey (Reference Fortey, Briggs and Lane1983) listed and illustrated a single specimen, it would become the holotype by monotypy. Peng (Reference Peng1990, p. 109) did distinguish the species as follows: “Hyperbolochilus ? forteyi differs from the present species [i.e., Proscharyia sinensis] only in the absence of the lateral glabellar furrows, and the relatively broader (sag.) proportion of the preglabellar field to the anterior border.” While not a formal diagnosis, this statement does serve a similar purpose as it purports to distinguish the species from a putatively related taxon. Hence, the species apparently represents an available name. Despite obviously treating it as a species of Proscharyia, nowhere in his paper did Peng (Reference Peng1990) refer to it as “Proscharyia forteyi n. sp.,” only as “Hyperbolochilus ? forteyi,” and the latter is the binomen with which the species was established.
Hyperbolochilus forteyi resembles both P. sinensis and A. palaora in general dimensions but seems to share more features with the latter. In particular, it lacks incised glabellar furrows and has distinct eye ridges. Much more information would obviously be necessary to evaluate it properly, but for the time being, it is tentatively assigned to Amechilus rather than Proscharyia.
Amechilus palaora Ross, Reference Ross1951
Figure 9
- Reference Ross1951
Amechilus palaora Ross, p. 112, pl. 28, fig. 15.
- Reference Hupé1953
Amechilus palaora; Hupé, p. 67, fig. 5.1.
- Reference Kobayashi1955
Amechilus palaora; Kobayashi, p. 459.
- Reference Erben1961
Amechilus palaora; Erben, p. 88, text-fig. 2a.
- Reference Dean1988
Amechilus palaora; Dean, p. 11.
- Reference Ross, Hintze, Ethington, Miller, Taylor and Repetski1997
Amechilus palaora; Ross et al., p. 18.
- Reference White and Lieberman1998
Amechilus palaora; White and Lieberman, p. 6.
- Reference Jell and Adrain2003
Amechilus palaora; Jell and Adrain, p. 340.
- Reference Adrain, Karim, Westrop and Landing2014
Amechilus palaora; Adrain et al., p. 179.
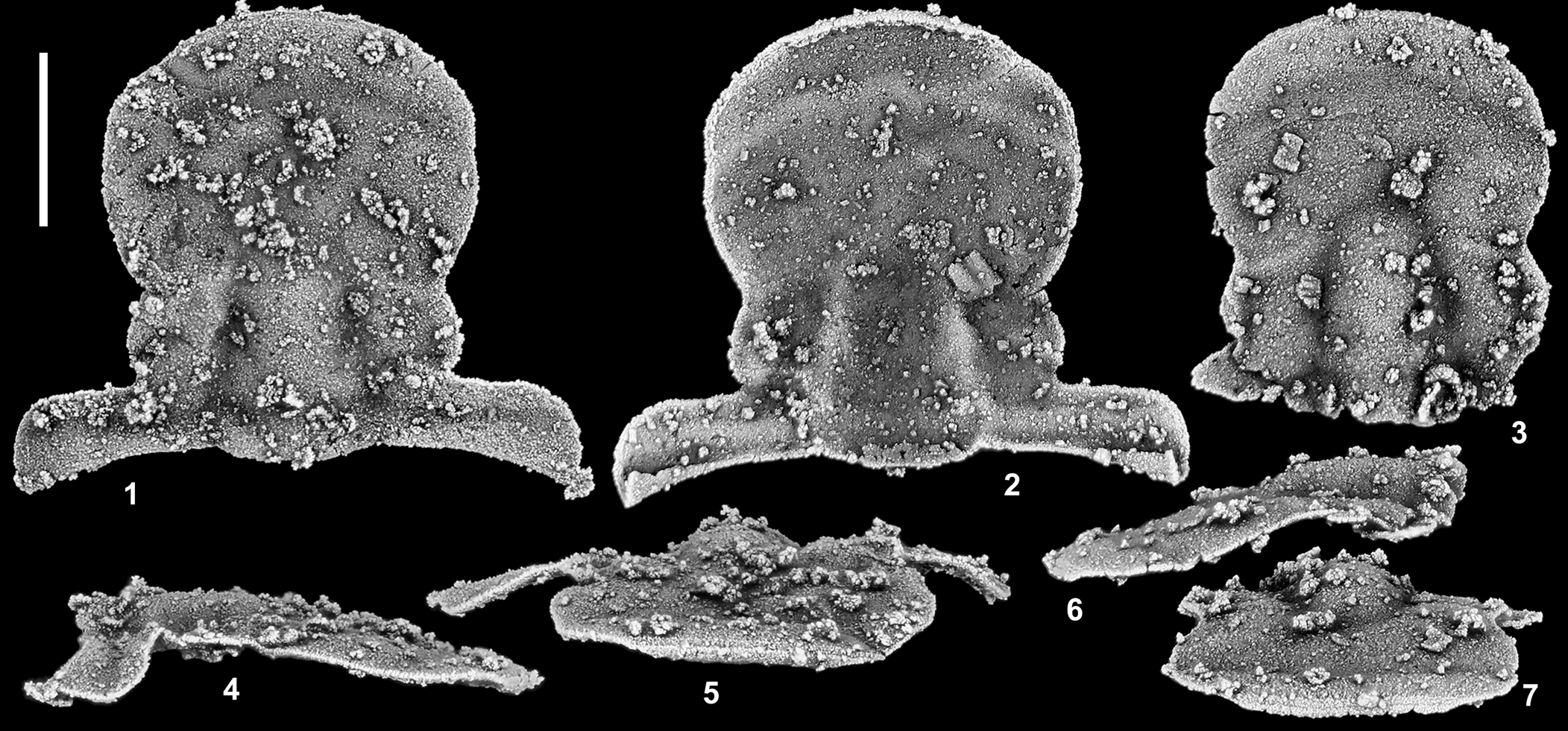
Figure 9. Amechilus palaora Ross, Reference Ross1951 from Section MME 84.0 m, Fillmore Formation (mid-Tremadocian; Stairsian; Bearriverops deltaensis Zone), Middle Mountain, Ibex area, Millard County, western Utah, USA. (1, 2, 4, 5) cranidium, SUI 148413: (1) dorsal, view; (2) ventral view; (4) right lateral view; (5) anterior view. (3, 6, 7) Cranidium, SUI 134142: (3) dorsal view; (6) left lateral view; (7) anterior view. Scale bar = 1 mm.
Holotype
Cranidium, YPM 18182 (Ross, Reference Ross1951, pl. 28, fig. 15), from the Garden City Formation (mid-Tremadocian; Stairsian), Locality 5, east side of Hillyard Canyon, Bear River Range, Franklin County, southeastern Idaho, USA.
Diagnosis
As for genus.
Occurrence
New material is from Section MME 84.0 m, Fillmore Formation (mid-Tremadocian; Stairsian; Bearriverops deltaensis Zone), Middle Mountain, Ibex area, Millard County, western Utah, USA.
Materials
SUI 134142, 148413.
Remarks
Amechilus palaora has not been encountered in the Garden City Formation despite extensive sampling. Two cranidia, however, have been recovered from the Bearriverops deltaensis Zone in the Fillmore Formation, western Utah. They are nearly identical to Ross's (Reference Ross1951, pl. 28, fig. 15) single illustrated cranidium and unquestionably belong to the same species. Unfortunately, the species is extremely rare, and no librigenae or pygidia that could be associated with it have been recovered.
Genus Proscharyia Peng, Reference Peng1990
Type species
Proscharyia sinensis Peng, Reference Peng1990 from the Madaoyu Formation (upper Tremadocian), Panjiazui, Taoyuan, northwest Hunan, China (South China).
Other species
Protarchaeganus [sic] sanduensis Zhou, Reference Zhou1981, Guotang Formation (lower Tremadocian), Guizhou, China (South China).
Diagnosis
See Peng (Reference Peng1990, p. 108).
Remarks
Protarchaeganus [sic] sanduensis Zhou, Reference Zhou1981, was not mentioned when Proscharyia was proposed, but it is exceedingly similar to Proscharyia sinensis. The only substantial differences are an anterior border that appears to be slightly longer in general, and in particular to be of similar length, versus shorter, exsagittally versus sagittally, a relatively larger glabella, and a pygidium that is shorter (sag.) and wider than some of those assigned to Proscharyia sinensis, although it is more comparable to others (e.g., Peng, Reference Peng1990, pl. 19, figs. 14, 15). As Proscharyia sanduensis is known from only two internal mold specimens and neither species is known from librigenae, detailed comparison is necessarily limited.
Zhou et al. (Reference Zhou, Zhou and Yin2016, p. 322, fig. 21L) illustrated a pygidium from the Darriwilian Zhuozishan Formation of Inner Mongolia as “Proscharyia sp.” The morphology of this specimen certainly seems consistent with an oenonellid as it has five axial rings and well-expressed pleural bands and furrows that extend to the margin. Other affinities (e.g., proetid, rorringtoniid) are difficult to rule out, however. In the absence of an associated cranidium, the identification of the specimen as an oenonellid must be tentative.
Acknowledgments
S. Stouge provided guidance and assistance in the field in Newfoundland in 1996 and 2001, including measuring and logging the section and drafting the column upon which Figure 2 is based. T. Adrain assisted in the field in Newfoundland in 2001 and 2004 and managed loans and curation. M. Coyne provided GSC specimen numbers. B. Beck assisted in the field in Newfoundland in 2004. T. Karim assisted in the field in 2004 and permitted the inclusion of the Shallow Bay Formation specimen, which she photographed. A. Bradley assisted in the field in Utah in 2018 and processed critical samples. R. Ng assisted in the field in Utah in 2018. S. Morrison assisted with literature. F.-J. Lindemann arranged loan of material from the Natural History Museum, University of Oslo. J. Loch and an anonymous referee provided comments that improved the paper.










 Open access
Open access