



Introduction
Qilianotryma Xu in Jin et al., Reference Jin, Ye, Xu and Sun1979 is a mid-Katian (Ka2–3, Late Ordovician) atrypide brachiopod of the subfamily Spirigerininae first reported from Qilian Mountains of Qaidam terrane that is now part of Northwest China. Qilianotryma features a biconvex to dorsibiconvex profile and uniplicate anterior commissure typical of many early atrypides in this region (e.g., Eospirigerina, Rongatrypa, and Sulcatospira) but is distinctive in featuring an elliptical transverse outline and fine multicostellate ornamentation unlike the subpentagonal to subtriangular outline and coarse costae of most other atrypides from this region in the Upper Ordovician. The internal shell morphology of Qilianotryma mirabile Xu in Jin et al., Reference Jin, Ye, Xu and Sun1979, the type species, is poorly known although the internal morphologies of other species (e.g., Qilianotryma suspectum) have been described (Popov et al., Reference Popov, Nikitin and Sokiran1999).
Qilianotryma has previously been reported from the Koumenzi Formation (Katian of the Upper Ordovician) in Qilian Mountains, Qinghai Province, Northwest China. Qilianotryma suspectum from Henan Province represents the first occurrence of this species in South China, however, and is illustrated herein. The holotype of the type species Qilianotryma mirabile from the Koumenzi Formation of Qilian Mountains was rephotographed to illustrate diagnostic features of the species and genus more clearly and restudied to emend the diagnosis for this genus.
Cocks and Modzalevskaya (Reference Cocks and Modzalevskaya1997) first reported Qilianotryma outside of China on the basis of fossils initially described as Euroatrypa tajmyrica Nikiforova in Nikiforova et al., Reference Nikiforova, Oradovskaya and Popov1982 collected from the Katian of Russian central Taimyr (Popov in Nikiforova et al., Reference Nikiforova, Oradovskaya and Popov1982). Since then, there has been some debate as to whether Euroatrypa is a valid genus or should be accepted as a junior synonym of Qilianotryma (e.g., Cocks and Modzalevskaya, Reference Cocks and Modzalevskaya1997; Popov et al., Reference Popov, Nikitin and Sokiran1999; Nikitin et al., Reference Nikitin, Popov and Bassett2006; Rong et al., Reference Rong, Zhan, Huang, Xu, Fu, Li, Rong, Jin, Shen and Zhan2017). Species assigned to Euroatrypa have been differentiated from Qilianotryma on the basis of a more globose outline, a shorter hinge line, and a fold and sulcus not originating from the beak (Nikitin et al., Reference Nikitin, Popov and Bassett2006). Figured specimens reveal considerable morphological variation in Euroatrypa through its ontogeny (Nikiforova et al., Reference Nikiforova, Oradovskaya and Popov1982), however, so there is still some uncertainty as to the validity of Euroatrypa.
By more clearly defining the characteristics and range of this important early atrypide brachiopod genus, this study will contribute to an ongoing effort to better understand brachiopod evolution and biogeography in the plates that now comprise China and central Asia during the Early Paleozoic as new lineages were evolving during the Ordovician Radiation. A better understanding of the evolution and distribution of Qilianotryma and other atrypide lineages will be key to understanding the evolution and dispersal of brachiopods in this region. Given the poor paleomagnetic coverage in these regions, fossils are a particularly useful tool available to plot the location of these plates and terranes through time.
Geological setting
The study area is located in Xichuan County, southwestern Henan Province, central China (Fig. 1). This region was part of a shallow-water platform on the north margin (present orientation) of South China paleoplate during the Late Ordovician (e.g., Ma et al., Reference Ma, Chen and Wang2009; Dong et al., Reference Dong, Zhang, Neubauer, Liu, Genser and Hauzenberger2011; Rong et al., Reference Rong, Zhan, Wang, Huang, Tang and Luan2015; Jing et al., Reference Jing, Stouge, Ding, Wang and Zhou2017). South China is thought to have been located in the tropical latitudes to the west of Gondwana at this time (Popov and Cocks, Reference Popov and Cocks2017; Torsvik and Cocks, Reference Torsvik and Cocks2017; Jin et al., Reference Jin, Zhan and Wu2018). Several paleoplates (e.g., North China, Tarim) and terranes (e.g., Chu-Ili, Chingiz-Tarbagatai, Atasu-Zhamshi) were also located in the tropical region and adjacent to South China west to Gondwana in the Late Ordovician.

Figure 1. Simplified geographical maps of central China and the study area in Xichuan County, southwestern Henan Province, central China, modified from Zhang et al., Reference Zhang, Zhan, Sproat and Huang2019b.
The Shiyanhe Formation is composed mainly of a series of gray thick-bedded grainstones interbedded with yellow thin-bedded siltstone and greenish mudstones. Macrofossils (e.g., brachiopods, corals, trilobites, bryozoans, and bivalves) are diverse and abundant (e.g., Yan et al., Reference Yan, Xi and Wang1983; Deng, Reference Deng1987; Liu et al., Reference Liu, Wang, Zhang and Du1991; Zeng et al., Reference Zeng, Liu, Wan, Pei, Yan, Zhang and Du1993; Xu, Reference Xu1996; Zhang et al., Reference Zhang, Zhan, Sproat and Huang2019b). The coral fauna is dominated by Agetolites and Agetolitella (Deng, Reference Deng1987), two genera that are found in shallow-water settings across South China and North China during the Late Ordovician. Conodonts collected from the Shiyanhe Formation belong to, ascendingly, the Oulodus ulrichi, Oulodus robustus, Aphelognathus grandis, and Aphelognathus divergens biozones, indicating a Katian (Late Ordovician) age (Ka2–Ka4) (Jing et al., Reference Jing, Stouge, Ding, Wang and Zhou2017). The Shiyanhe Formation unconformably underlies the Zhangwan Formation (Llandovery, Silurian).
Brachiopod shells are abundant in grainstones and siltstone interbeds at the locality, and loose specimens can be easily collected from weathered surfaces. Internal molds are also well preserved in siltstone and silty mudstone. This brachiopod fauna consists of 18 species within 18 genera. The strophomenide Sowerbyella (Sowerbyella) sinensis Wang, Jin, and Fang, Reference Wang, Jin and Fang1964 and atrypide Rongatrypa xichuanensis (Xu, Reference Xu1996) are the most abundant species in this fauna, while spiriferide, lingulide, and protorthide brachiopods are very rare from our collection.
Materials and methods
Specimens examined in this study were collected from the Shiyanhe Formation at the Yaziwo section (33°07′50″N, 111°18′32″E, WGS84) in southwestern Xichuan County, China. More than 1,000 brachiopod specimens were collected from this formation; however, only two specimens have been confidently identified as Qilianotryma. Both valves were collected from the reddish massive carbonate rocks in the upper part of the formation.
Fossil materials were prepared, measured with digital calipers, coated with ammonium chloride powder (Teichert, Reference Teichert1948), and photographed under an optical microscope. One conjoined shell was serially sectioned with a refined Croft parallel grinder (Zhang et al., Reference Zhang, Sproat, Zhan, Zhang, Luan and Huang2019a). Serial sections were produced on acetate peels, photographed under an optical microscope, and manually traced to illustrate the internal morphology of the shell. The software PAST 3.20 (Hammer et al., Reference Hammer, Harper and Ryan2001) was used to plot scatter charts and run discriminant analysis on the dataset of measurements.
Repository and institutional abbreviation
Two conjoined valves and 80 acetate peel replications of transverse sections examined in this study are deposited at Nanjing Institute of Geology and Palaeontology (NIGP), Chinese Academy of Sciences, Nanjing, China (NIGP 173727–173728). The holotype of Qilianotryma mirabile was collected by Hankui Xu in the Koumenzi Formation at the upstream of Tianbao river, Qilian Mountains, Qaidam terrane, Northwest China. The remains of the serial sectioned holotype specimen and a complete epoxy replica are deposited at Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences (NIGP 42574).
Systematic paleontology
Order Atrypida Rzhonsnitskaia, Reference Rzhonsnitskaia1960
Suborder Atrypidina Rzhonsnitskaia, Reference Rzhonsnitskaia1960
Superfamily Atrypoidea Gill, Reference Gill1871
Family Atrypinidae McEwan, Reference McEwan1939
Subfamily Spirigerininae Rzhonsnitskaia, Reference Rzhonsnitskaia1974
Genus Qilianotryma Xu in Jin et al., Reference Jin, Ye, Xu and Sun1979
Type species
Qilianotryma mirabile Xu in Jin et al., Reference Jin, Ye, Xu and Sun1979 (p. 97, 98, in-text fig. 40, pl. 20, figs. 23, 24); collected from the Koumenzi Formation (Katian, Upper Ordovician), Qilian Mountains, Northwest China.
Other species
Qilianotryma suspectum (Popov in Nikiforova et al., Reference Nikiforova, Oradovskaya and Popov1982) (p. 57, pl. 6, figs. 9–12); type specimens initially identified as Euroatrypa suspecta in Nikiforova et al. (Reference Nikiforova, Oradovskaya and Popov1982); collected from the Katian of the Chu-Ili terrane, Kazakhstan. Qilianotryma cf. Q. suspectum Popov in Nikiforova et al., Reference Nikiforova, Oradovskaya and Popov1982 (p. 67, pl. 6, figs. 9–12) from the Degeres Member of the Dulankara Formation, Chu Ili terrane, Kazakhstan; see also Popov and Cocks (Reference Popov and Cocks2021, p. 94–95, pl. 10, fig. 19) from an unnamed formation in southern Kazakhstan. Qilianotryma sp. Popov and Cocks, Reference Popov and Cocks2014 (p. 749, fig. 25X, Y), collected from the Akdombak Formation of the Chingiz Terrane, Kazakhstan.
Diagnosis
See Rong et al., Reference Rong, Zhan, Huang, Xu, Fu, Li, Rong, Jin, Shen and Zhan2017.
Occurrence
Koumenzi Formation (Katian, Upper Ordovician) of the Qilian Mountains, Qaidam terrane, Northwest China; Shiyanhe Formation (Katian, Upper Ordovician) of Xichuan County, Henan Province, South China; Degeres and Akkol Members, Dulankara Formation (Katian, Upper Ordovician) of the Dulankara Mountains and an unnamed formation (Katian, Upper Ordovician) of the Ergenekty Mountains, southern Chu-Ili Range, Kazakhstan; Odak Beds of the Angrensor Formation (Katian, Upper Ordovician), central Kazakhstan; Akdombak Formation (Katian, Upper Ordovician), Mount Akdombak area, Chingiz, Kazakhstan.
Remarks
Qilianotryma and its type species Qilianotryma mirabile was established on the basis of a single specimen found in the Koumenzi Formation (Katian, Upper Ordovician) of Qilian Mountains, Northwest China (Xu in Jin et al., Reference Jin, Ye, Xu and Sun1979). This genus was previously assigned to the family Ancistrorhynchidae within the Rhynchonellida because of its similarity to Ancistrorhyncha Ulrich and Cooper, Reference Ulrich and Cooper1942 in terms of external morphology (Xu in Jin et al., Reference Jin, Ye, Xu and Sun1979). However, the presence of jugal processes in the mantle cavity (Xu in Jin et al., Reference Jin, Ye, Xu and Sun1979, in-text fig. 40) indicates this genus should instead be assigned to the Atrypida and with affinities to the Atrypidae, although spiralia are not illustrated in the initial description of the species and are not visible in the holotype specimen. It also represents a major difference compared with other Late Ordovician early atrypide lineages such as the Anazygidae that have a solid jugum rather than jugal processes (see Copper, Reference Copper1977; Sproat and McLeod, Reference Sproat and McLeod2023) although the function of these structures remains poorly understood.
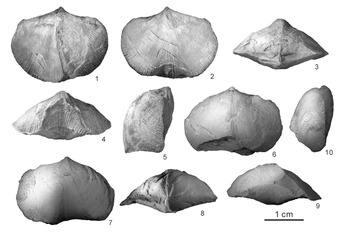
The type species Qilianotryma mirabile is poorly known due to the ambiguity of the original figure and the hard access to the initial fossil locality in northern Tibet (Jin et al., Reference Jin, Ye, Xu and Sun1979). The holotype is rephotographed in this study (Fig. 2), indicating a less globose and longer hinge line compared with Euroatrypa type species (i.e., Euroatrypa tajmyrica, Nikiforova et al., Reference Nikiforova, Oradovskaya and Popov1982, pl. 6, figs. 3, 4) and other species (e.g., Euroatrypa aitenensis, Nikitin et al., Reference Nikitin, Popov and Bassett2006, fig. 35.1, 35.3).

Figure 2. Holotype of Qilianotryma mirabile Xu in Jin et al., Reference Jin, Ye, Xu and Sun1979 from the Koumenzi Formation at Tianbaohe upper stream section, Qilian County, Qinghai Province, northwestern China. (1–4) Holotype, NIGP 42575: (1) dorsal view; (2) ventral view; (3) lateral view; (4) anterior view. (5–9) Epoxy cast representing the complete holotype, NIGP 42575: (5) dorsal view; (6) ventral view; (7) lateral view; (8) anterior view; (9) posterior view. Dashed lines represent the surface that the shell was ground down to. Scale bar = 5 mm.
A parasulcate commissure and lateral furrows (and plications) on either side of the dorsal fold and ventral sulcus are known in some species of Qilianotryma (e.g., Qilianotryma suspectum, Popov et al., Reference Popov, Nikitin and Sokiran1999), but these characteristics are not common in species of the genus. Other species have a more subtle division between the fold and sulcus and the flanks of each valve (e.g., Qilianotryma mirabile).
Some shells can be only questionably assigned to the genus. Qilianotryma cf. Q. suspectum Popov and Cocks, Reference Popov and Cocks2021 resemble Q. suspectum in overall character, but only a single dorsal valve interior mold was available for study, and thus the shell can be assigned only tentatively to the genus. Similarly, Qilianotryma sp. Popov and Cocks, Reference Popov and Cocks2014 is similar to the type species, Q. mirabile, in the exterior features of the shell, such as the fine ribs and uniplicate anterior. Given that no shell interiors or serial sections have been made to study the internal morphology of the shells and that only a few deformed shells are known from the locality, its definitive assignment to the genus will be possible only with study of additional material.
Qilianotryma differs from Spirigerina and Eospirigerina in its finely, evenly multicostellate radial ornamentation and transverse outline. Qilianotryma also has fewer spiralia whorls (usually fewer than seven) than typically found in Eospirigerina (fewer than 12 whorls). Qilianotryma differs from Anabaria and Rongatrypa in its fine radial ornamentation, thin dental plates in the ventral interior, and lack of a vertical myophragm and a cardinal process at the end of notothyrial cavity.
Euroatrypa Oradovskaya in Nikiforova et al., Reference Nikiforova, Oradovskaya and Popov1982 and its type species, Euroatrypa tajmyrica, are broadly similar to Qilianotryma and were established on the basis of materials from the Korotkaya Formation (Katian), central Taimyr of northern Siberia, Russia (Nikiforova et al., Reference Nikiforova, Oradovskaya and Popov1982). Additional specimens of Euroatrypa have been also found in the Tirekhtyakh horizon (Katian), Northeast Russia (Nikiforova et al., Reference Nikiforova, Oradovskaya and Popov1982) and the Odak Beds (Katian), central Kazakhstan (Nikitin et al., Reference Nikitin, Popov and Bassett2006). Euroatrypa was considered a subjective synonym of Qilianotryma by Cocks and Modzalevskaya (Reference Cocks and Modzalevskaya1997) and Copper (Reference Copper and Kaesler2002).
Nikitin et al. (Reference Nikitin, Popov and Bassett2006) maintained Euroatrypa as a separate taxon based on examination of new material of both Qilianotryma and Euroatrypa from the Angrensor Formation (Katian) from further collections at the Odak Beds in central Kazakhstan, suggesting these two genera should be treated as different taxa since Euroatrypa has a more globose shell, a shorter hinge line, and a ventral sulcus and dorsal fold that do not originate from the beak.
Available measurable specimens of Euroatrypa (Table 1) appear to be broadly similar to those of Qilianotryma in terms of length, width, and thickness (Fig. 3). Discriminant analysis, however, shows that the taxa can be readily differentiated, with a 77.3% correct classification based on shell dimensions alone (Fig. 4). With some hesitation, we accept these two genera are different in shell outline. The length of the hinge line and development of the dorsal fold and ventral sulcus are also essential in discriminating Qilianotryma from Euroatrypa. Euroatrypa is herein able to be differentiated from Qilianotryma as a valid genus.

Figure 3. (1, 2) Linear regression plots of width versus length and width versus depth of measurable specimens of Qilianotryma and previously identified Euroatrypa: (1) k = 0.64 and 0.75, r2 = 0.74 and 0.91, respectively; (2) k = 0.50 and 0.42, r2 = 0.84 and 0.65, respectively. Points in dark green and light green represent specimens previously described Qilianotryma and Euroatrypa, respectively. The data point with a red outline represents the holotype of the type species, Qilianotryma mirabile Xu in Jin et al., Reference Jin, Ye, Xu and Sun1979. Each point corresponds with specimens presented in Table 1.

Figure 4. Histogram of discriminant projection values for reported measurable specimens of Qilianotryma and Euroatrypa (Table 1). The cutoff point is set to zero. Blue, pink, and red zones represent Qilianotryma, Euroatrypa, and the mixture of two genera, respectively.
Table 1. Dimensions of measurable specimens of Qilianotryma and Euroatrypa from China and central Asia. L/W = length divided by width; D/W = depth divided by width. Specimens are measured in millimeters.

Qilianotryma suspectum (Popov in Nikiforova et al., Reference Nikiforova, Oradovskaya and Popov1982)
Figures 5, 6
- Reference Rukavishnikova1956
Cliftonia ex gr. spirigeroides (M'Coy, Reference M'Coy1851); Rukavishnikova, p. 151, pl. 4, figs. 14, 15.
- Reference Nikiforova, Oradovskaya and Popov1982
Euroatrypa suspecta Popov in Nikiforova et al., p. 57, pl. 6, figs. 9–12.
- Reference Popov, Nikitin and Sokiran1999
Qilianotryma suspectum; Popov et al., p. 634, pl. 1, figs. 11–15.
- Reference Nikitin, Popov and Bassett2006
Qilianotryma suspectum; Nikitin et al., p. 270, figs. 35 12a–e, 38 1–3, 39.
- Reference Popov and Cocks2006
Qilianotryma suspectum; Popov and Cocks, p. 279, fig. 6G–H, J.

Figure 5. Two specimens of Qilianotryma suspectum (Popov in Nikiforova et al., Reference Nikiforova, Oradovskaya and Popov1982) from the upper part of the Shiyanhe Formation at Yaziwo section, Xichuan County, southwestern Henan Province, central China. (1–5) Specimen NIGP 173727: (1) dorsal view; (2) ventral view; (3) posterior view; (4) anterior view; (5) lateral view. (6–10) Specimen NIGP 173728: (6) dorsal view; (7) ventral view; (8) posterior view; (9) anterior view; (10) lateral view. Scale bar = 1 cm.

Figure 6. Sketches of internal structures of brachiopod Qilianotryma suspectum (Popov in Nikiforova et al., Reference Nikiforova, Oradovskaya and Popov1982) from the upper part of the Shiyanhe Formation at Yaziwo section, Xichuan County, southwestern Henan Province, central China. Numbers below each sketch indicate the distance, in millimeters, of each section from the posterior of the shell. One spiralia on the right is broken and likely displaced from its position during life inside the shell. The catalog number of the studied cellulose acetate peels is NIGP 173728. Scale bar = 2 mm.
Holotype
A conjoined valve (CNIGR 25/11943) from Akkol Beds, upper Dulankara Formation (Katian) of the Dulankara Mountains, southern Chu-Ili Range, Kazakhstan (Nikiforova et al., Reference Nikiforova, Oradovskaya and Popov1982, p. 57, pl. 6, figs. 9–12).
Occurrence
Shiyanhe Formation (mid–upper Katian) in the Yaziwo section, Xichuan County, Henan Province, South China.
Description
Shell moderate in size, dorsibiconvex, elliptical transverse to subpentagonal in outline, about 70–80% as long as wide, and 45–67% as thick as wide (Fig. 5; Table 1). Anterior commissure uniplicate. Hinge line wide and astrophic, about four-fifths as long as shell width. Cardinal extremities rounded. Radial ornament finely and evenly multicostellate with 20–24 costellae on fold and sulcus and 31–44 costellae on lateral flanks; 5–6 costellae per 3 mm in anterior commissure of adult specimens (Fig. 5.1, 5.2).
Ventral valve slightly convex, with the maximum height at one-third length from beak (Fig. 5.5, 5.10). Beak incurved and pointed, apsacline interarea with open delthyrium. Ventral sulcus originating from the beak, becoming prominent and wide anteriorly (Fig. 5.2, 5.7). Semioval tongue-shaped sulcus taking up one-half to three-fifths of the shell width.
Dorsal valve moderately to strongly convex, with maximum height at two-thirds length from the beak (Fig. 5.5, 5.10). Median fold originating from the beak and forming a uniplicate commissure at anterior. Lateral furrows very weak, originating from the beak (Fig. 5.8).
Teeth simple and strong in ventral valve (Fig. 6, 1.2–1.9). Dental plates thin (Fig. 6, 1.2–2.05), slightly diverging dorsally, and thinning toward the anterior (Fig. 6, 1.2–2.05). Dental cavities subtriangular (Fig. 6, 1.5–1.8) to ellipsoid (Fig. 6, 2.05) in transverse sections. Delthyrial cavity wide, taking up one-sixth of the shell width and one-third of shell depth.
Socket plates strong in dorsal valve. Crural bases long and weak (Fig. 6, 1.8–2.0). Ventral median ridge with a pair of weak oblique submedian ridges present in dorsal valve. Spiralia delicate, with up to seven whorls (Fig. 6, 5.1–9.1). Jugal processes short, located ventroposteriorly (Fig. 6, 2.7). Muscle scars poorly impressed.
Materials
Two conjoined shells. Specimens NIGP 173727 and NIGP 173728 are figured in Figure 5. Internal morphology of specimen NIGP 173728 is illustrated in Figure 6.
Remarks
This species differs from the type species, Qilianotryma mirabile Xu in Jin et al., Reference Jin, Ye, Xu and Sun1979, in its larger size, less-elliptical outline, and more-dorsibiconvex lateral profile with lateral furrows on the dorsal valve. Qilianotryma suspectum from Chu-Ili, Kazakhstan, differs from this species in having 16–20 costellae on fold and sulcus and more-prominent lateral plications on ventral valves (Popov et al., Reference Popov, Nikitin and Sokiran1999). Euroatrypa kolymica Oradovskaya in Nikiforova et al., Reference Nikiforova, Oradovskaya and Popov1982 is smaller, equally biconvex in lateral profile, and has a stronger rectangular anterior commissure, taking up two-thirds of shell length and width. Euroatrypa tajmyrica Nikiforova in Nikiforova et al., Reference Nikiforova, Oradovskaya and Popov1982 is more ventribiconvex in some junior shells, with a prominent carination in the ventral umbonal region. The dorsal fold and ventral sulcus are weaker without any lateral furrows or plications and take up four-fifths of the width and two-thirds of the depth of the shell. Qilianotryma cf. Q. suspectum Popov and Cocks, Reference Popov and Cocks2021 has a less-elevated but wider dorsal fold compared with the specimens in this study.
Paleobiogeographic implications
Qilianotryma was first described from Qilian Mountains, Northwest China (Xu in Jin et al., Reference Jin, Ye, Xu and Sun1979) located on the northern Tibet Plateau and regarded as a part of the Qaidam terrane in the Late Ordovician. The Qaidam terrane was inferred to have been close to the Lhasa (or Qiangtang) terrane in the midlatitudes off the margin of western Gondwana and the neighboring South China paleoplate and Kazakh terranes during the Late Ordovician. However, the precise paleogeographic location of the Qaidam terrane is still uncertain due to poor paleomagnetic coverage. Recent studies reported the discoveries of Middle–Late Ordovician volcanic rocks in the Halahu of this region, perhaps indicating an active margin (Li et al., Reference Li, Zhang, Wang, Liu, Qiao, Kui, Li, Zhao, Xue and Li2019, Reference Li, Zhang, Cao, Wang, Wang, Wang, Qiao, Chen and Chen2020).
Qilianotryma is known from a brachiopod fauna in the Koumenzi Formation of this region. This fauna consists of Altaethyrella, Bokotorthis, Eospirigerina, Liostrophia, ?Mimella, Porambonites, Ptychoglyptus, Qilianotryma, Sowerbyella, Strophomena, Sulcatospira, and Triplesia (Xu in Jin et al., Reference Jin, Ye, Xu and Sun1979; Rong et al., Reference Rong, Zhan, Huang, Xu, Fu, Li, Rong, Jin, Shen and Zhan2017), indicating a shallow-water, midshelf environmental setting (Rong et al., Reference Rong, Zhan, Huang, Xu, Fu, Li, Rong, Jin, Shen and Zhan2017).
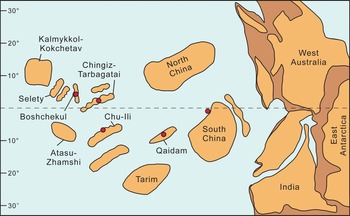
Outside China, Qilianotryma is known from the Chu-Ili, Boshchekul, and Chingiz-Tarbagatai terranes of Kazakhstan (Fig. 7; Nikiforova et al., Reference Nikiforova, Oradovskaya and Popov1982; Popov et al., Reference Popov, Nikitin and Sokiran1999; Nikitin et al., Reference Nikitin, Popov and Bassett2006; Popov and Cocks, Reference Popov and Cocks2006, Reference Popov and Cocks2014, Reference Popov and Cocks2021). Qilianotryma sp. was noted from Akdombak Formation (upper Katian) of Chingiz, Kazakhstan (Popov and Cocks, Reference Popov and Cocks2014). Qilianotryma cf. Q. suspectum (Popov in Nikiforova et al., Reference Nikiforova, Oradovskaya and Popov1982) is known from an unnamed formation (early to mid-Katian) of Chu-Ili, Kaakhstan (Popov and Cocks, Reference Popov and Cocks2021). Qilianotryma suspectum has a wide distribution compared with other species. It has been noted in the Platymena–Strophomena, Strophomena–Christiania, Metambonites–Plectorthis, and Parastrophina–Kellerella associations in the Akkol Member, Dulankara Formation, Chu-Ili of Kazakhstan (Popov and Cocks, Reference Popov and Cocks2006) and Odak of Kazakhstan (Nikitin et al., Reference Nikitin, Popov and Bassett2006). Brachiopod fauna in the Shiyanhe Formation in this study is also characteristic of Altaethyrella and Schachriomonia, showing a faunal similarity with those in Kazakhstan.

Figure 7. The distribution of Qilianotryma in the tropical region west to Gondwana during the Late Ordovician. Paleogeographical map is modified from Popov and Cocks, Reference Popov and Cocks2017. The individual localities of Qilianotryma are from Rukavishnikova, Reference Rukavishnikova1956, Jin et al., Reference Jin, Ye, Xu and Sun1979, Nikiforova et al., Reference Nikiforova, Oradovskaya and Popov1982, Popov et al., Reference Popov, Nikitin and Sokiran1999, Nikitin et al., Reference Nikitin, Popov and Bassett2006, and Popov and Cocks, Reference Popov and Cocks2006.
Altaethyrella is the most widely spread genus in the faunas where Qilianotryma was found. These faunas were named Altaethyrella Fauna (Rong and Zhan, Reference Rong, Zhan, Rong and Fang2004), which flourished mainly in the well-oxygenated shallow-water marine setting with high faunal richness and diversity. Altaethyrella Fauna has been known to distribute in different paleoplates or terranes of the Russian Altai Mountains, Kyrgyzstan, Turkmenistan, Kazakhstan, South China, North China, and Northwest China (e.g., Severgina, Reference Severgina1978; Zhan and Li, Reference Zhan and Li1998; Popov and Cocks, Reference Popov and Cocks2017; Rong et al., Reference Rong, Zhan, Huang, Xu, Fu, Li, Rong, Jin, Shen and Zhan2017; Sproat and Zhan, Reference Sproat and Zhan2018). These localities are widely accepted to have been located in the tropical region west to Gondwana during the Late Ordovician (e.g., Chen et al., Reference Chen, Bergström, Zhang and Wang2013; Harper et al., Reference Harper, Rasmussen, Liljeroth, Blodgett, Candela, Jin, Percival, Rong, Villas and Zhan2013; Popov and Cocks, Reference Popov and Cocks2017; Torsvik and Cocks, Reference Torsvik and Cocks2017).
The limited geographic range of Qilianotryma reaffirms that Chu-Ili, Boshchekul, and Chingiz-Tarbagatai terranes of Kazakhstan, Qaidam terrane of Northwest China, and South China Paleoplate were located close to each other. The co-occurrence of Qilianotryma suspectum in Chu-Ili terrane and South China matches the recent paleogeographic reconstruction (Popov and Cocks, Reference Popov and Cocks2017), supporting the interpretation that Chu-Illi and South China were adjacent to each other during the Late Ordovician. The relationship to these plates will become increasingly clear as more of the shallow-water brachiopod fauna of these plates is documented and described.
Evolutionary significance
Qilianotryma, like many atrypides in these regions, belongs to the subfamily Spirigerininae Rzhonsnitskaia, Reference Rzhonsnitskaia1974. Brachiopods in this subfamily lack frills, have fewer whorls in the spiralia, and are typically more equally biconvex than later atrypides in the Silurian and Devonian (e.g., Copper, Reference Copper1967, Reference Copper1977, Reference Copper1978); they also possess jugal processes rather than a solid jugum or jugal plates. Almost all Qilianotryma are Late Ordovician in age, with only one species questionably assigned to Qilianotryma from the Silurian (Baarli, Reference Baarli2021). Qilianotryma? sigridi Baarli, Reference Baarli2021 was collected from Padda and Leangen members of the Solvik Formation (Aeronian, Silurian) in Norway (Baltica; Baarli, Reference Baarli2021) and is similar to the type species of Qilianotryma in external morphology but distinct in some key diagnostic characters (e.g., medium-coarse ribs, fibrous crura, and the presence of a cardinal process). No definitive Qilianotryma are known to have survived the first pulse of the End-Ordovician Mass Extinction.
Beitaia Rong, Xu, and Yang, Reference Rong, Xu and Yang1974 from the upper Rhuddanian and Aeronian of Llandovery, Silurian in South China, and western Tarim (Rong et al., Reference Rong, Zhan, Huang, Xu, Fu, Li, Rong, Jin, Shen and Zhan2017) may represent the successor to Qilianotryma. This genus is similar to Qilianotryma in having fine and continuous costellate ornamentation, dorsibiconvex lateral profile in adult shells, and the presence of jugal process in the transverse section. Beitaia differs from Qilianotryma in its subquadrate outline, shorter hinge line, ventral sulcus originated from half of the shell, and distinct concentric filae on the full shell.
Qilianotryma is very different from the atrypides that dominated other paleoequatorial plates during the Late Ordovician. The most common and well known of Late Ordovician atrypides from Laurentia are the Anazygidina. Anazyga and Zygospira appear to look similar to their counterparts from Asia, but their external morphologies differ in terms of their reversed fold and sulcus and the structure of the spiralia compared with most atrypides from mid- and East Asia (Copper, Reference Copper1977; Sproat and McLeod, Reference Sproat and McLeod2023). Most shallow-marine North American atrypides possess a jugum rather than separate jugal processes typical of Asian lineages, indicating that they likely evolved along different evolutionary pathways (see Popov et al., Reference Popov, Nikitin and Sokiran1999; Baarli et al., Reference Baarli, Huang and Maroja2022). The evolutionary pressures driving the divergence of these forms remain uncertain, but these differences hint at different evolutionary pressures acting on brachiopods in each of these equatorial zones.
Conclusions
This first report of Qilianotryma suspectum (Popov in Nikiforova et al., Reference Nikiforova, Oradovskaya and Popov1982) from the Shiyanhe Formation of Henan Province, central China, significantly expands the geographical range of the genus. The rephotograph of type material and discriminant analysis support that Qilianotryma and broadly similar Euroatrypa can be differentiated in terms of external morphology, the length of the hinge line, and the development of the median fold and sulcus. The presence of Qilianotryma in shallow-marine communities across the South China paleoplate, Qaidam terrane, and several Kazakh terranes (i.e., Chu-Ili, Boshchekul, and Chingiz-Tarbagatai) suggests that the low-latitude brachiopod province once thought to stretch across these plates and North America may not be as homogeneous as once thought although further analysis of other lineages is needed to confirm this. Perhaps this provincialism was a consequence of either underlying paleogeographic factors or the evolution of different novel characteristics in brachiopods during the Ordovician Radiation.
Acknowledgments
We are grateful to T. Shcherbanenko for translating Russian references, H. Zhang for laboratory support, J. Rong, Y. Wang, B. Huang, P. Tang, and X. Luan for support in the field. We thank L.E. Popov and R. Freeman for their constructive reviews and editor S. Zamora and associate editor Z. Zhang for their editorial comments on the manuscript. Financial support was provided to Y.Z. and R.Z. by the National Natural Science Foundation of China (grant nos. 42202021, 41972011) and the Strategic Priority Research Program of the Chinese Academy of Sciences (grant no. XDB26000000). Funding to C.D.S. was provided by a Natural Sciences and Engineering Research Council of Canada Discovery Grant. This paper is also a contribution to IGCP project 735 “Rocks and the rise of Ordovician life.”
Declaration of competing interests
The authors declare none.







 Open access
Open access