


The Neotropical genus Atelopus (Anura: Bufonidae) has declined dramatically; 70% of 113 species have suffered population crashes (Lötters et al., Reference Lötters, La Marca, Stuart, Gagliardo and Veith2004; La Marca et al., Reference La Marca, Lips, Lötters, Puschendorf, Ibañez and Ron2005; Pounds et al., Reference Pounds, Bustamante, Coloma, Consuegra, Fogden and Foster2006; IUCN, 2007). There are 10 endemic Atelopus species in Venezuela (La Marca & Reinthaler, Reference La Marca and Reinthaler1991). Nine of these are categorized as Critically Endangered and one is possibly extinct (Rodríguez & Rojas-Suárez, Reference Rodríguez and Rojas-Suárez1995; IUCN, 2007). Despite intensive sampling in their former habitats, most species had not been observed since the 1980s (La Marca & Lötters, Reference La Marca, Lötters, Böhme, Bischoff and Ziegler1997; Manzanilla & La Marca, Reference Manzanilla and La Marca2004). Although the causes of many amphibian population declines are not yet well understood, chytridiomycosis, a disease caused by the pathogenic fungus Batrachochytrium dendrobatidis, has been frequently associated with Atelopus declines (Berger et al., Reference Berger, Speare, Daszak, Green, Cunningham and Goggin1998; Lips, Reference Lips1998; Bonaccorso et al., Reference Bonaccorso, Guayasamin, Méndez and Speare2003; Lampo et al., Reference Lampo, Rodríguez, La Marca and Daszak2006).
Once common, Atelopus cruciger disappeared for almost 20 years and was believed to be extinct (Manzanilla & La Marca, Reference Manzanilla and La Marca2004). However, recent sightings of A. cruciger at the Cordillera de La Costa in the Venezuelan coastal range and A. mucubajiensis at the Cordillera de Mérida in the Venezuelan Andes indicated they may yet be extant (Eliot, Reference Eliot2003; Barrio-Amorós, Reference Barrio-Amorós2004). We therefore evaluated the current distribution of A. cruciger and the presence of B. dendrobatidis in its remnant populations.
To construct a historical distribution map of the species we examined data from five national (Museo Estación Biológica de Rancho Grande, MEBRG; Museo de Biología de la Universidad Central de Venezuela, MBUCV; Museo de Historia Natural La Salle, MHNLS; Museo del Instituto de Zoología Agrícola de la Universidad Central de Venezuela, MIZA; Dr. Manuel González Sponga's private collection) and eight US museum collections (California Academy of Sciences, CAS; Carnegie Museum of Natural History, CM; Field Museum of Natural History, Chicago, FMNH; Museum of Natural History, University of Kansas, KU; Museum of Comparative Zoology, Harvard University, Cambridge, Massachussets, MCZ; Museum of Vertebrate Zoology, University of California, Berkeley, MVZ; University of Michigan Museum of Zoology, UMMZ; National Museum of Natural History, Washington, DC, USNM), and reviewed literature on sightings and collections (Sexton, Reference Sexton1958; Bonaccorso et al., Reference Bonaccorso, Guayasamin, Méndez and Speare2003; Manzanilla & La Marca, Reference Manzanilla and La Marca2004). From the historical distribution map we selected 15 locations for field exploration, based on accessibility, safety, and likely persistence of the original habitats (Fig. 1, Table 1).
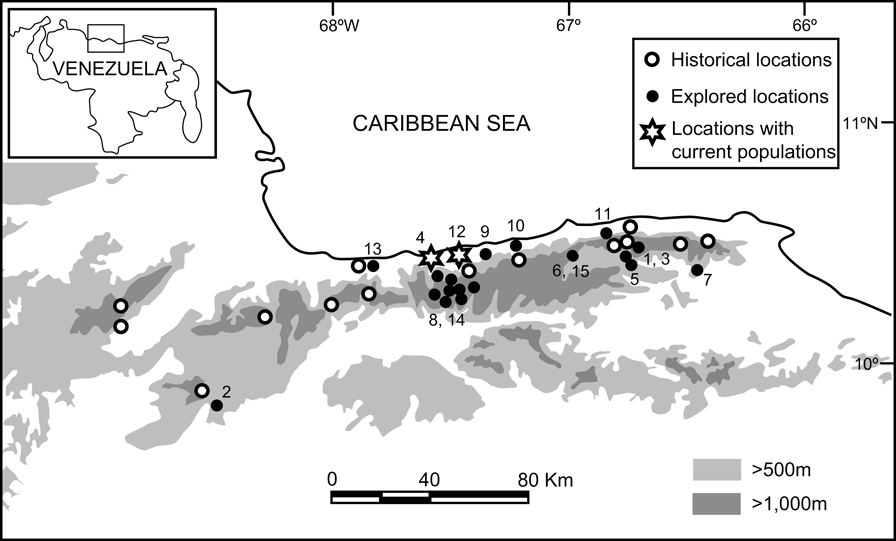
Fig. 1 Known historical distribution of Atelopus cruciger based on museum records, with the locations that we explored and the current distribution of the species (for further details see Table 1). Inset shows the location of the main map in Venezuela.
Table 1 Locations explored in the search for Atelopus cruciger (see numbered locations on Fig. 1).
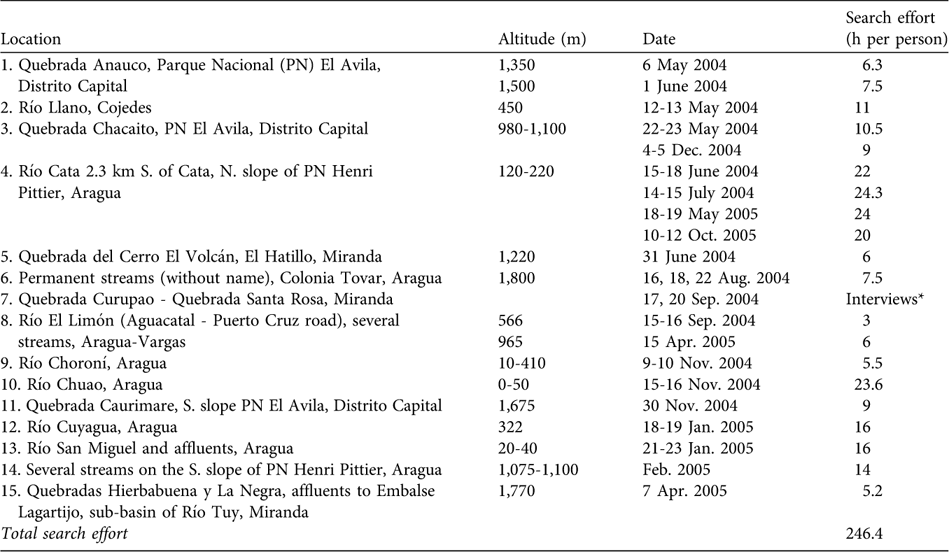
* We did not explore this location because it is unsafe but interviews with local people suggested the species is not present.
We found 627 museum records of A. cruciger from 33 locations distributed over most of the Cordillera de La Costa (Fig. 1). The data indicated that A. cruciger was once abundant, especially in streams along the road between Maracay and Choroní (mostly inside the Parque Nacional Henri Pittier), and on the southern slope of the Parque Nacional El Avila, next to Caracas. The altitudinal distribution of the species was 0-2,400 m but most records were over 500-2,000 m in gallery, cloud and semi-deciduous forests and their ecotones with littoral vegetation and agricultural areas.
We searched for A. cruciger using visual encounter survey methods (Heyer et al., Reference Heyer, Donelly, McDiarmid, Hayek and Foster1994; Lips et al., Reference Lips, Mendelson, Muñoz-Alonso, Canseco-Márquez and Mulcahy2004) between 08.00 and 16.00 to maximize the probability of sighting this diurnal species (Sexton, Reference Sexton1958). Each individual encountered was photographed. We sampled 23 individuals for B. dendrobatidis using swabs and then released them (Kriger et al., Reference Kriger, Hines, Hyatt, Boyle and Hero2006). Biosecurity protocols were followed to avoid pathogen dispersal and sample contamination (Aguirre & Lampo, Reference Aguirre, Lampo, Angulo, Rueda-Almonacid, Rodríguez-Mahecha and La Marca2006). For detection of B. dendrobatidis and zoospore quantification we extracted nucleic acids from swabs with PrepMan Ultra and conducted real time Taqman PCR assays (Boyle et al., Reference Boyle, Boyle, Olsen, Morgan and Hyatt2004). Standard curves were constructed by using 100, 10, 1 and 0.1 B. dendrobatidis zoospore quantification standards provided by the Australian Animal Health Laboratory.
After a total of 246 person hours of intensive searching (Table 1) we found A. cruciger at only two locations (4 and 12; Table 1, Fig. 1) at altitudes of 220 and 322 m, respectively, on the northern slope of the Parque Nacional Henri Pittier. This suggests that A. cruciger persists on the northern slope of the Cordillera de La Costa but in a more restricted area than formerly. We encountered 83 individuals including juveniles and adult males and females (Table 2; Plate 1). The presence of juveniles suggests these two populations may be reproducing and viable, although larval stages were not detected. One adult male was found morbid, at Río Cata (location 4 in Fig. 1). A DNA analysis of tissue samples from this male revealed high levels of infection (244,484 zoospores in 1-3 mg of tissue). Excessive skin sloughing indicated that chytridiomycosis was probably implicated (Berger et al., Reference Berger, Speare and Kent1999). Infection was also detected in three out of 23 apparent healthy adults (from location 4), although zoospore loads were very low (1.0-3.9 zoospores in 1-3 mg of tissue).

Plate 1 Atelopus cruciger adult male sighted at Río Cata (location 4 on Fig. 1) on 11 October 2005. Photo: F. Rojas.
Table 2 Atelopus cruciger individuals found in two locations at the Parque Nacional Henry Pittier (see numbered locations on Fig. 1).

The apparent synchrony of rediscovery of a few Atelopus species in different regions (Eliot, Reference Eliot2003; Barrio-Amorós, Reference Barrio-Amorós2004; Black, Reference Black2006) suggests that recent climatic conditions could be favouring population recoveries. It is also possible that this synchronized reappearance is a consequence of the relative synchrony in the disappearance of species phylogenetically related and with similar life history traits. A. cruciger was rediscovered within a year of the detection of one adult female A. mucubajiensis in the Andes (Barrio-Amorós, Reference Barrio-Amorós2004). Furthermore, a third population of A. cruciger was rediscovered in February 2006 in Río El Duro, Edo, Aragua (J. Valera & M. Frontado, pers. comm.), also on the northern slope of the Parque Nacional Henri Pittier. According to the chytrid-thermal-optimum hypothesis, a decrease in the maximum daily temperatures over the last decade promoted the growth of B. dendrobatidis in amphibians, which led to high mortalities (Pounds et al., Reference Pounds, Bustamante, Coloma, Consuegra, Fogden and Foster2006). The fact that all A. cruciger populations detected are at <350 m is consistent with this hypothesis. Maximum temperatures at these sites are greater than at highland altitudes, although other non-exclusive explanations are also possible. Dry conditions, for instance, could have also favoured pathogen transmission rates triggering epidemic events that led to population crashes (Lampo et al., Reference Lampo, Rodríguez, La Marca and Daszak2006). It is possible that A. cruciger populations are now experiencing a post epidemic recovery, with low prevalence and infection loads of B. dendrobatidis. However, the discovery of a severely infected morbid frog also suggests that other infected frogs could develop the disease and die. The climate-linked hypothesis predicts that, unless regional climatic trends are reversed, new epidemics are likely to occur where the pathogen persists endemically.
To ensure management in any case of new epidemics that could jeopardize the long-term survival of A. cruciger, we need to monitor the prevalence of B. dendrobatidis infection and the sizes of remnant populations. As long as treatments cannot be effectively applied in the field, ex situ management will remain the only available conservation strategy. However, in situ conservation could be a feasible alternative in the short-term. Close monitoring of climatic, ecological and epidemiological variables, which we are now conducting, are fundamental for the identification of threshold indicators that could help us decide when and where to apply management strategies.
Acknowledgements
This project was funded by the Iniciativa de Especies Amenazadas, the Museo de Historia Natural La Salle and the Instituto Venezolano de Investigaciones Científicas. Permits were provided by The Ministerio del Ambiente y de los Recursos Naturales. We thank Fernando Rojas, Juan Carlos Rodríguez, Gilson Rivas, Arnaldo Ferrer, Oscar Lasso, Guillermo García, Julian Mora, Maigualida Malavé and Alex Malavé for their support during fieldwork, and Fabiola León for DNA detection of B. dendrobatidis. The comments of Kathryn M. Rodríguez-Clark and Dinora Sánchez-Hernández greatly improved this manuscript.
Biographical sketches
Argelia Rodríguez-Contreras is interested in the conservation of threatened amphibian species and the link between amphibian declines, chytridiomycosis and climate change. J. Celsa Señaris, the Director of the Museo de Historia Natural La Salle, works on the systematics and taxonomy of the Venezuelan herpetofauna. Margarita Lampo has a broad interest in population ecology and epidemiology, specializing in amphibians. Ramón Rivero is the curator of the herpetological section of the Museo Estación Biológica de Rancho Grande, and researches the taxonomy of Venezuelan amphibians.




