



Introduction
Cattle, particularly indigenous breeds, are indispensable in alleviating rural poverty, and employment generation in developing countries. Bos indicus cattle are well adapted to hot and humid climatic conditions prevailing in the tropics, especially in Southeast Asia (Sodhi et al., Reference Sodhi, Mukesh, Kishore, Mishra, Kataria and Joshi2013),
Efforts to maximize the superior indigenous cattle were made possible by incorporating advanced reproductive biotechnologies. In this endeavour, ovum pick-up and in-vitro embryo production (OPU-IVEP) has been added as the second biotechnological intervention for improving genetic gain and production. Exogenous use of hormones and proper knowledge of ovarian follicular dynamics have been considered two important pillars for maximizing the number of quality oocytes and progeny from a genetically superior cow. The gonadotropins stimulation before OPU proved to be an efficient method to harvest a large number of follicles for aspiration (Fry, Reference Fry2020).
For in vivo recovery of the oocytes, USG-guided ovum pick-up has proved to be an efficient, repeatable, less-invasive technique over the years and sets the foundation for the successful implementation of assisted reproductive technology (Qi et al., Reference Qi, Yao, Ma, Wang, Zhao, Liu and Sun2013; Landeo et al., Reference Landeo, Zuñiga, Gastelu, Artica, Ruiz, Silva and Ratto2022). Oocytes recovered from slaughterhouse-derived ovaries are of unknown genetics and heterogeneous in quality with varying developmental competence (Bessa et al., Reference Bessa, Nishimura, Franco and Dode2013). Whereas, IVEP success depends mainly on the proper growth of follicles and subsequent the quality of oocytes, therefore, oocyte quality is of prime importance for determining the competence. Oocyte developmental competence is a complex process that translates the ability of the oocyte to complete maturation, undergo successful fertilization, reach the blastocyst stage and yield viable and healthy progeny following embryo transfer (Orozco-Lucero & Sirard, Reference Orozco-Lucero and Sirard2018).
Morphological assessment is being used for evaluation of the oocytes and its selection for OPU-IVEP varies among the users and based on the experience of the evaluator which is largely misinterpreted. Many times, these morphological assessments are not reliable enough to act as the sole criteria for the evaluation of oocyte competence which stresses to have an alternate selection criterion for selecting the viable oocytes for IVEP. Hence, an alternative mechanism by identifying molecular markers to predict oocyte competence appears a major researchable issue (Khurchabilig et al., Reference Khurchabilig, Sato, Ashibe, Hara, Fukumori and Nagao2020). Therefore, the focus centred on the identification of molecular markers to select the most potential oocytes for embryo production and embedded with higher implantation efficiency (Ruvolo et al., Reference Ruvolo, Fattouh, Bosco, Brucculeri and Cittadini2013) in Bos indicus were lacking. The present study is framed on the above facts and attempts to examine the effect of FSH stimulation on oocyte developmental competence parameters of OPU-derived Bos indicus (Tharparkar) oocytes by evaluating the various morphological parameters and expression profiles of genes viz., GDF9, BMP15, PCNA, BCL-2 and BAX related to folliculogenesis, cumulus expansion, oocyte maturation and apoptosis.
Materials and methods
Location and climatic parameters
The experiment was conducted during the summer (April to July) of 2023 at Cattle and Buffalo Farm, ICAR-IVRI, Izatnagar, located at an altitude of 564 metres above mean sea level, at latitude and longitude of 28° N and 79°E, respectively. The climate of the region is humid and tropical with the mean temperature during summer was 30.7°C (min 15.1- max 40.9°C), and relative humidity was 53.2.
Animals and feed management
A total of ten parous, clinically normal, cyclic and non-lactating Tharparkar cows (aged 6.80 ± 0.81 years; body weight 395.60 ±6.50 kg) were selected from the Cattle and Buffalo farm for oocyte aspiration. All cows were fed 1.5– 2.5 kg of concentrate feed containing 20% digestible crude protein and 70% total digestible nutrients, had ad-lib access to wheat straw, green fodder two times a day, and ad-lib access to drinking water. All the animals were maintained under hygienic and optimal management conditions in a semi-intensive system with access to a large, open paddock for free movement. Heat detection in the herd was carried out twice daily at 07:00 and 16:00 using a vasectomized bull (teaser). Health and vaccination protocols were followed as per the standard schedule of farm management.
Experimental design
Before the start of the experiment, all the cows were screened to rule out any uterine infections/or any reproductive abnormalities by per-rectal as well as ultrasound examinations. All ten cyclic, dry, Tharparkar cows were synchronized with a single dose of PGF2α (EstrumateTM, MSD Animal Health), and all oestrus cows were randomly divided into two groups comprising five cows in each group. Further, five oestrus cows underwent ovum pick-up (OPU) on the 2nd day of oestrus and the remaining five cows were subjected to FSH stimulation (Stimufol®, Reprobiol, Belgium) at the dose of 130 µg divided into four sub-doses (40, 40, 25 and 25 µg morning and evening, I/M) before OPU. OPU was performed 48 h (coasting period) after the last FSH injection. All Tharparkar cows were given the rest of one and half cycles (33.20 ± 0.71 days) and again synchronized using single dose PGF2α and grouped in such a way that non-stimulated animals of the previous set of experiments were subjected to FSH stimulation and the remaining five animals were subjected to OPU on 2nd day of oestrus cycle without stimulation. In summary, a total of 20 OPU sessions were performed which included 10 sessions each in FSH-stimulated (n = 10) as well as non-stimulated (n = 10) Tharparkar cows during the early luteal phase (day-2) of the oestrus cycle in the absence of dominant follicle and corpus luteum (Figure 1).
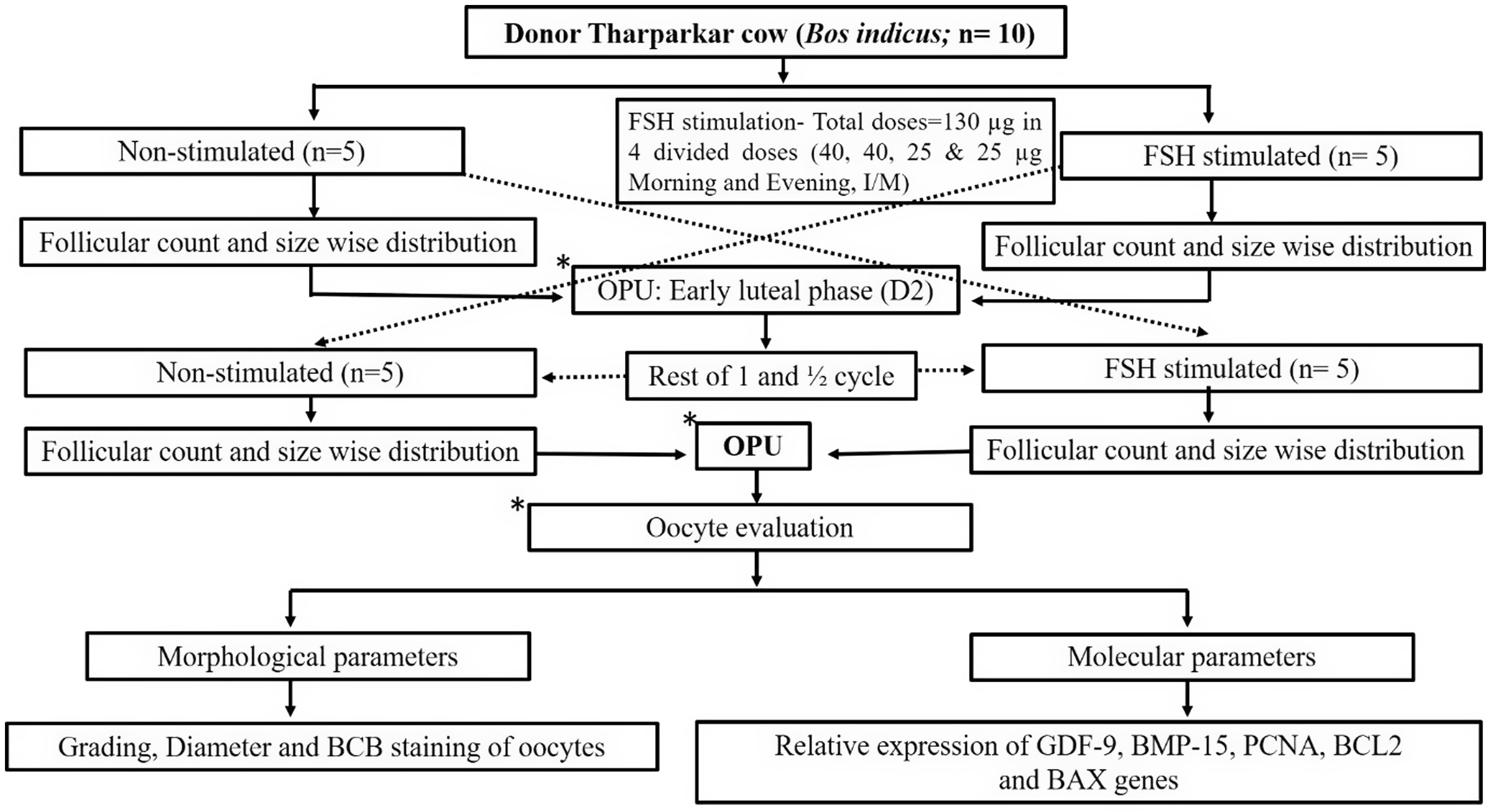
Figure 1. Experimental design.
Preparation of donor and ovum pick-up
The donors were properly restrained, and rectal evacuation of dung was performed with a gloved hand., followed by cleaning of the perineal region with 70% ethanol. Before each OPU session, cows were given 4 ml of Xylocain® 2% (Lignocain Hydrochloride, Zydus Cadila, India) epidurally to prevent abdominal straining and facilitate ovarian handling and smooth follicle aspiration.
A lubricated transvaginal probe was inserted into the anterior vagina and positioned against the ovary for viewing and aspirating follicles. Data on follicle number and diameter were recorded using a caliper on a monitor image and categorized into groups i.e. follicles ≥6 mm and <6 mm. Once the ovary and target follicle were stabilized, follicular aspiration was performed by a single trained technician using a real-time B-mode ultrasound scanner (Draminski 4 Vet-slim, Poland) equipped with a multi-frequency (2–14 MHz) micro convex probe. During the entire OPU session, the needle and aspiration line were thoroughly rinsed with pre-warmed (37 °C) OPU recovery medium (IVF Bioscience, UK) to prevent blood clotting or oocytes from sticking to the tubing and collecting conical tube (50 ml). A negative pressure of 85 to 90 mm Hg was generated using a vacuum aspiration (Wireless) pump with a thermostat (WTA, Brazil) to create a vacuum and generate a fluid flow of 15–20 ml per min. for both groups. Transvaginal ultrasonographic images of an FSH-stimulated and non-stimulated donors ovaries before (Figure 2 A & 2 C) and after Ovum Pick up (Figure 2 B & 2 D) respectively. Oocyte recovery rate (%) was calculated by using the following formula:
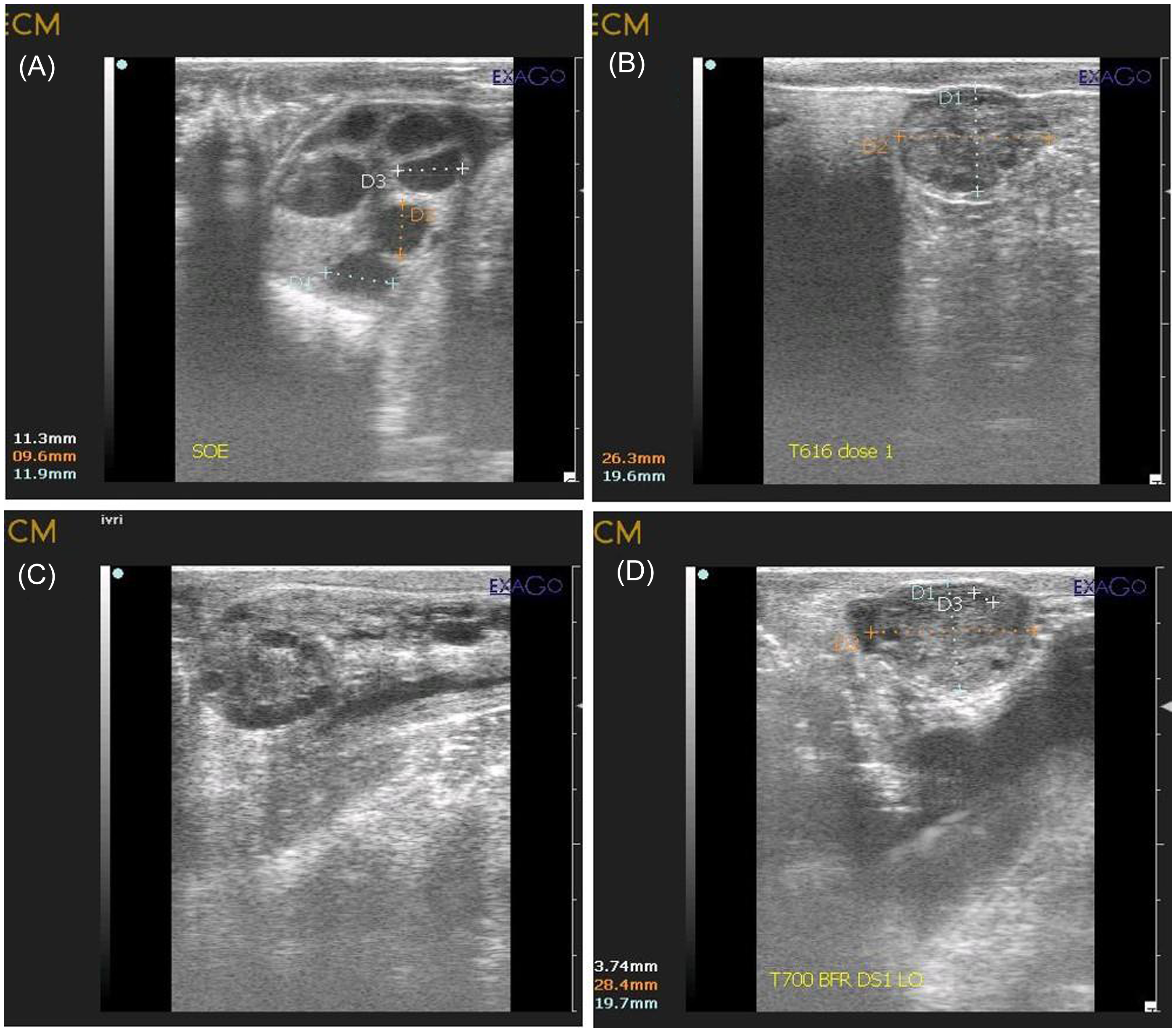
Figure 2. Transvaginal ultrasonographic images of stimulated and non-stimulated ovaries. (A) Partially stimulated ovary prior to OPU showing numerous follicles of ≥ 6mm. (B) Partially stimulated ovary after transvaginal oocytes aspiration showing very few left over small size follicles. (C) Non-stimulated ovary on day −2 of oestrous cycle prior to OPU showing numerous growing follicles of < 6mm. (D) Non-stimulated ovary on day −2 of oestrous cycle after transvaginal oocytes aspiration showing very few left over follicles.
• Oocyte recovery rate = number of oocytes recovered/number of follicles aspirated.
Evaluation of morphological parameters of oocytes
Washed and filtered follicular aspirate was transferred to a square grid Petri dish (90 × 15 mm, Tarsons, Chennai) and examined under a thermal stage-fitted zoom stereo microscope (SMZ 1000, Nikon, Japan) at 20x magnification to identify cumulus-oocyte complexes (COCs). COCs were transferred to a 35 mm Petri dish containing BO wash medium (IVF Bioscience, UK) and examined under a zoom stereomicroscope at 63 x magnification. The collected oocytes of both groups were graded primarily based on the layer of cumulus cells under a stereozoom microscope as per the procedure described by Demetrio et al. (Reference Demetrio, Hasler, Oliveira, Demetrio, Fonseca and Santos2021). Oocyte quality deterioration is expressed using alphabetical coding in ascending order: A>B>C>D The sum of A, B and C is considered good-quality oocytes. Further, oocytes of both the groups were measured for their diameter as the mean length of two perpendicular axes using a ZEISS Axio inverted microscope (ZEISS Axio, Germany) and grouped based on the diameter i.e. ≥120 µm and <120 µm (Fair et al., Reference Fair, Hyttel and Greve1995).
The Brilliant Cresyl Blue staining (Sigma Chemicals Co.) was performed following the protocol of Manjunatha et al. (Reference Manjunatha, Gupta, Devaraj, Ravindra and Nandi2007). First, the Cumulus-Oocyte Complexes (COCs) were washed thrice in Dulbecco’s Phosphate-Buffered Saline (D-PBS) containing 0.4% Bovine Serum Albumin. Then, the COCs were exposed to 26 µM BCB solution for 90 minutes at 38.5°C temperature and 5% CO2 in a humidified air atmosphere. After staining, the COCs were washed twice with D-PBS and observed under a stereozoom microscope after 5 minutes. The percentage of BCB+VE and BCB-VE oocytes was recorded accordingly in both the groups. The oocytes were classified into two groups based on the cytoplasm colour i.e. oocytes having any degree of blue cytoplasm were assigned as BCB+VE and oocytes without any blue cytoplasm coloration were designated as BCB-VE.
Evaluation of relative expression profile of important genes in COCs of oocytes
Expression levels of GDF-9, BMP-15, PCNA, BCL-2 and BAX were identified in the oocytes retrieved through OPU by qPCR.
Total RNA extraction
Extraction of total RNA for the relative quantification of important gene transcripts from the COCs surrounded by oocytes retrieved through the OPU technique was carried out by trizol method and stored at −80ºC until used. The purity and concentration of total RNA were checked using the Nanodrop Spectrophotometer reading at OD 260 and 280. Samples with OD 260/280 values between 1.8 and 2.0 were used for further study.
Reverse transcription (RT)
A 20 ng of the total RNA (extracted from 15 to 20 COCs) was used for cDNA preparation with Prime Script™ 1st strand cDNA Synthesis Kit (Cat. #6110A, TAKARA BIO INC., USA), using oligodT primers following manufacturer’s instructions. cDNA formation was confirmed by PCR reactions set up in a final volume of 25 μL having 10× PCR buffer, 10 mM dNTPs, 1.0 U of Dream Taq DNA polymerase, and 10 pmol/μl each of forward and reverse primers for Glyceraldehyde-3-phosphate dehydrogenase (GAPDH), at 56°C annealing temperature. The cDNA was properly labelled and stored at −20 ºC for later use. The quality of cDNA was assessed by an amplification reaction for a housekeeping gene GAPDH. The PCR reactions for other gene-specific primers were standardized using gradient PCR to find the most suitable annealing temperature for the corresponding primers.
Primer designing
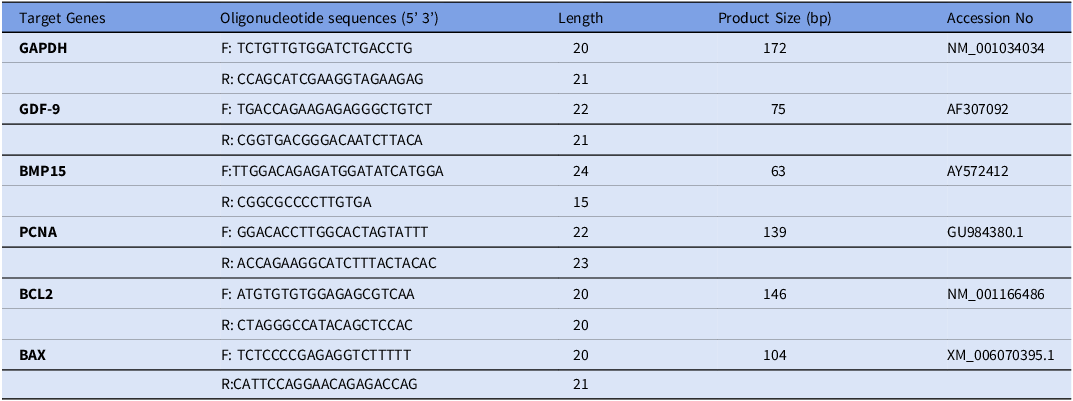
Initially, checked the published complete or partial gene sequence of the target gene in the National Center for Biotechnology (NCBI) database https://www.ncbi.nlm.nih.gov. Then, we analyzed the accuracy of forward and reverse primers using the oligo analyzer tool (https://www.idtdna.com/ pages/tools/oligo analyzer), followed by the NCBI blast analysis. Table 1 contains the sequence of the primers for the targeted genes and the housekeeping gene (GAPDH).
Table 1. List of primers used in qPCR for amplification of different target genes in COC’s of oocytes and its relation to oocyte quality
Real-time PCR
The qPCR reaction was carried out in a Smart Cycler Real Time qPCR (Cepheid, USA) spectrofluorometer using Platinum® SYBR® Green qPCR SuperMix-UDG (Cat#: 11733-038, Invitrogen Corporation, USA) to amplify all the target and housekeeping genes. The annealing temperature of each determined factor was standardized in qPCR using cDNA synthesized from COCs extracted total RNA. For the experiment, an opaque qPCR strip (Bio-rad) with optically clear flat caps was used. The reaction mixture was prepared which comprised of 5µL SYBR Green master mix, 0.25 µL each of forward and reverse primers, 20 ng of template cDNA, and nuclease-free water to make a total volume of 10 µL. GAPDH was taken as the housekeeping gene. A No-Template Control reaction was also run alongside to check for non-specific amplification.
After preparing the master mix, the strips underwent centrifugation or a short spin. Smart cycler protocol of real-time PCR was kept as: 2-min hold at 50°C for UDG incubation followed by a 2-min hold at 95°C to initiate the cycling conditions. The cycling conditions consisted of denaturation at 95°C for 15 seconds, annealing at 56°C for 30 s, and extension at 72°C for 30 s, which were repeated for 40 cycles. The fluorescent gain reading was taken at the end of each cycle to record the threshold values (ct). These values were used to calculate the fold change expression of the targeted gene. Subsequently, a melt curve was added, which involved gradual heating from 65–95°C at a rate of 1°C per second. The fluorescent data was collected during the temperature change. Finally, the obtained dissociation curve is analyzed for specific amplification of the gene, and to check for primer dimer formation resulting from non-specific amplification. After the run had ended, cycle threshold values, amplification plot and dissociation curve for all the transcripts were acquired and recorded. The real-time PCR specificity was further confirmed by the melt curve analysis. The results are the average 2ddCt values ± SEM of qPCR assays.
Statistical analysis
Data obtained in all the experiments were analyzed using the statistical software GraphPad Prism 8.0.2. The effect of FSH stimulation on the follicle size, oocyte diameter, BCB staining, total follicles aspirated, total oocytes recovered, oocytes recovery rate (%), and morphology of cumulus-oocyte complexes (COCs) was compared using unpaired ‘t’-test between the groups. The relative gene expression after FSH stimulation in both the groups (FSH-stimulated and non-stimulated) was measured by the ratio of Etarget and Eref, where Etarget is the efficiency of the target gene transcript and Eref is the efficiency of the housekeeping gene transcript. The difference in mean values for all data analyzed with P < 0.05 was considered significant.
Results
The oestrous cycle was monitored using B-mode USG regularly and the recorded number of follicles (22.6 ± 1.84) per cow indicates the good status of follicular pool in indigenous Tharparkar cows even in the absence of FSH stimulation. The FSH stimulation of Tharparkar cows leads to a significantly higher number of ≥6 mm size follicles, oocyte recovery rate and recovered oocytes per session as presented in Table 2. The classification of oocytes based on COC’s grades showed that there was a significant difference (P value <0.05) between the non-stimulated and stimulated Tharparkar cows, except in Grade D. The stimulated cows had a significantly higher number of Grade A oocytes as compared to the non-stimulated cows. Conversely, the non-stimulated cows had a higher number as well as percentage of Grade B and C oocytes than the FSH-stimulated Tharparkar cows. On the other hand, we could not notice any significant difference for Grade D oocytes in either of the groups. as shown in Table 3a. There was a significantly higher number as well as percentage of oocytes having a diameter ≥120 µm and BCB+VE oocytes in the FSH stimulated as compared to the non-stimulated Tharparkar cows respectively as shown in Table 3b, 3c and Figure 3. The relative mRNA expression profile of GDF9, BMP15, PCNA and BCL-2 (Anti-apoptotic) genes was 1.53 ± 0.15, 1.23 ± 0.04, 0.88 ± 0.05 and 1.37 ± 0.02 folds higher in the FSH-stimulated cows, on the other hand, BAX (Pro-apoptotic gene) was lower by 0.59 ± 0.09 fold in the FSH-stimulated Tharparkar cows as shown in Figure 4.
Table 2. Follicular aspiration, oocyte yields and recovery rates in FSH-stimulated and non-stimulated Tharparkar cows (Bos indicus) subjected to transvaginal ovum pick-up

Table 3. Oocyte developmental competence based on various morphological attributes retrieved through transvaginal ovum pick-up in Tharparkar cows (Bos indicus)
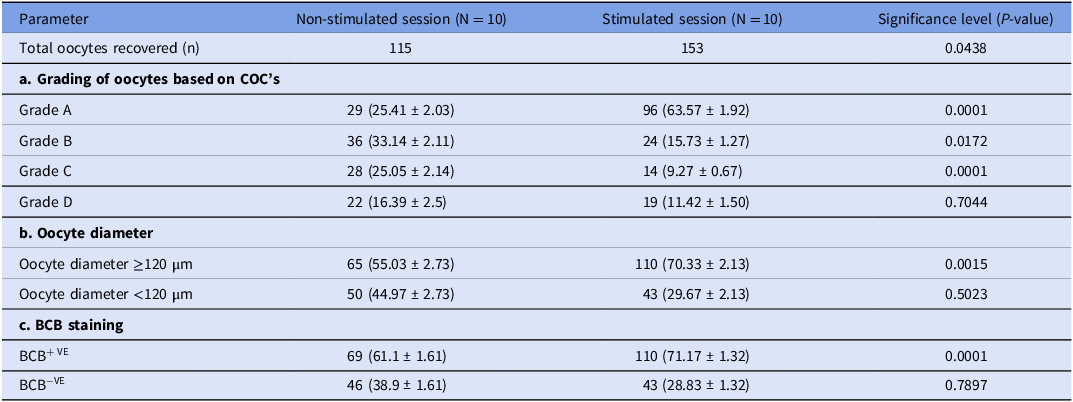
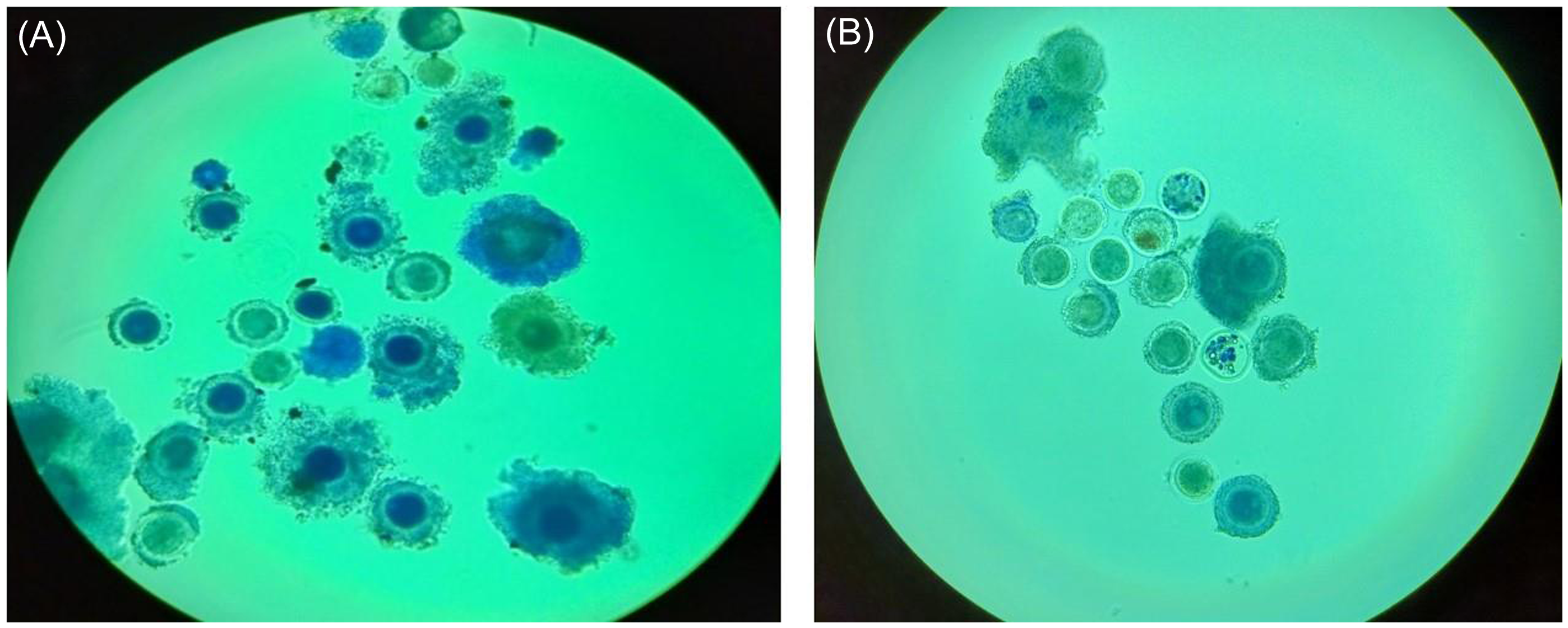
Figure 3. Brilliant Cresyl Blue (BCB) staining of oocytes retrieved through Transvaginal Ovum pick-up. (A) Numerous BCB+VE oocytes (showing cytoplasm blue coloured) recovered from stimulated Donor. (B) Very few BCB+VE recovered from non-stimulated Donor.
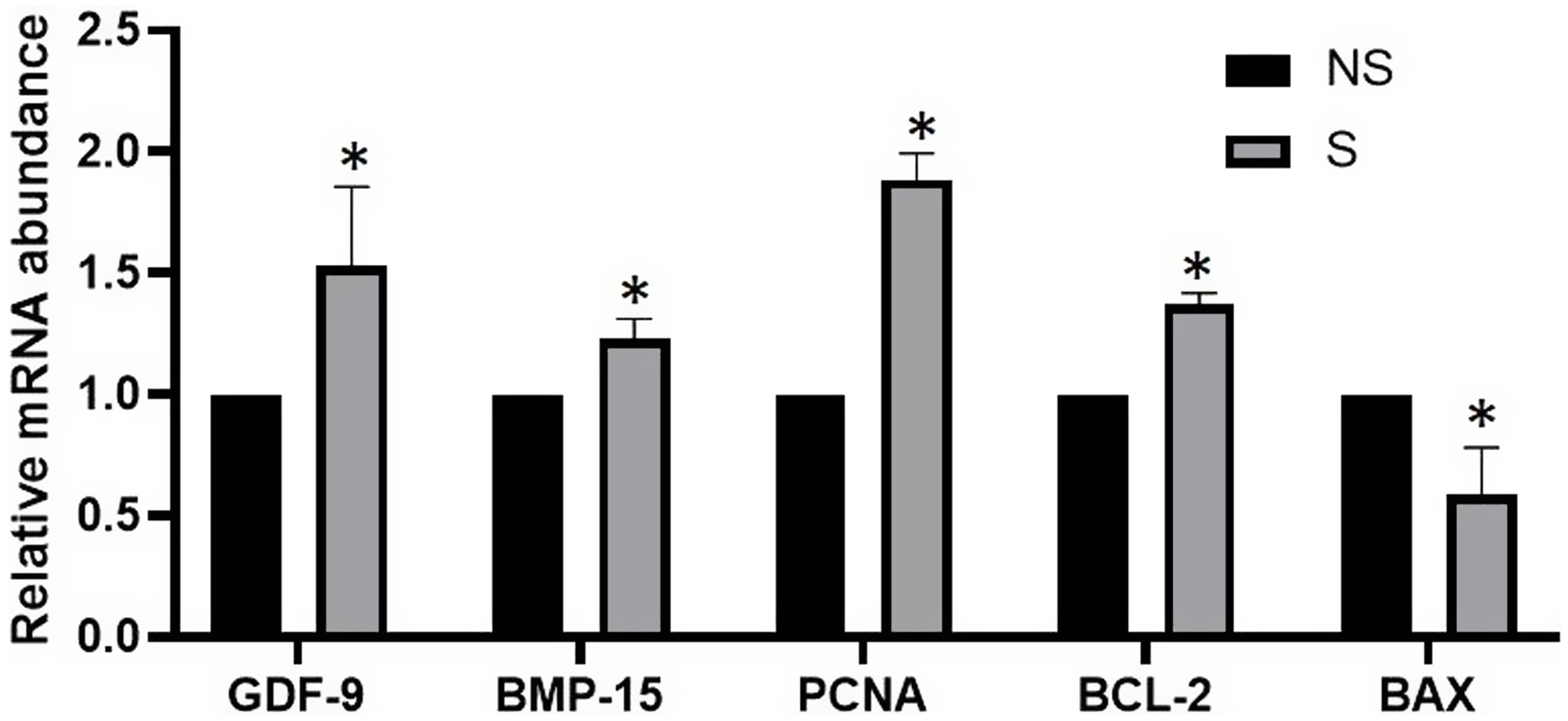
Figure 4. Relative expression of GDF9, BMP15, PCNA, BCL-2 and BAX gene transcripts on cumulus-oocyte complex retrieved through transvaginal ovum pick-up in FSH-stimulated Tharparkar cows. The values represent the mean ± s.e.m. of separate qPCR experiments. Bars with (*) superscripts differ significantly (P < 0.05). (NS = Non-stimulated, S = Stimulated).
Discussion
A higher mean number of total follicles available for aspiration (24.00±0.94) per OPU session was recorded in FSH-stimulated cows though statistically non-significant as compared to non-stimulated cows (22.6 ± 1.84) and many other reports (Chaubal et al., Reference Chaubal, Ferre, Molina, Faber, Bols, Rezamand and Yang2007; Sendag et al., Reference Sendag, Cetin, Alan, Hadeler and Niemann2008) recorded similar observation. The possible explanation may be that FSH increases the size of the follicles (>6 mm) rather than the number of follicles. Earlier reports also indicated that ovarian stimulation had no significant effect on the number of follicles available for aspiration (Patil et al., Reference Patil, Hadiya, Layek, Gorani, Raj, Karuppanasamy and Gupta2022). The number of follicles aspirated per cow per OPU session was higher in Tharparkar cows irrespective of FSH stimulation than the reports of Kang et al. (Reference Kang, Kim, Lee, Lee, Han and Cho2019) and Hadimani et al. (Reference Hadimani, Khillare, Amle, Meshram, Rangnekar, Mali and Nimbalkar2022) and almost corroborated with the observation recorded by Patil et al. (Reference Patil, Hadiya, Layek, Gorani, Raj, Karuppanasamy and Gupta2022) and Krishna et al. (Reference Krishna, Rao, Veerabramhaiah, Kumar, Srikanth and Yasaswini2023) this may be due to breed differences viz. Bos indicus has a higher number of follicular pools than Bos taurus.
The size of the follicles from which the oocytes are retrieved is a key factor in determining IVEP rates (Caixeta et al., Reference Caixeta, Ripamonte, Franco, Junior and Dode2009). In the present study, there were significantly higher numbers as well as mean percentage of ≥6 mm size and total aspirated follicles in FSH-stimulated cows than in the non-stimulated Tharparkar cows may be due to follicles of ≥6 mm suitable for follicular aspiration in OPU. Different researchers observed that the follicles ranging from 6–10 mm showed good developmental competence (Ongaratto et al., Reference Ongaratto, Cedeño, Rodriguez-Villamil, Tríbulo and Bó2020; Hayden et al., Reference Hayden, Sala, Absalón-Medina, Motta, Pereira, Moreno and García-Guerra2022). Findings by Alves et al. (Reference Alves, Cordeiro, de Lima, Annes, Dos Santos, Ispada and Milazzotto2019) reported that the follicular fluid microenvironment of large follicles (≥6 mm) has higher levels of electrolytes, glucose, reactive oxygen species, glutathione superoxide dismutase activity, lipids, cholesterol, pyruvate and oestradiol resulting in higher oocyte developmental competence, and thus better embryonic development. Further, FSH after binding with the FSH receptor modifies follicular cells and increases the size of the follicles (>6 mm) leading to an increase in oocyte competency and cumulus expansion. Stimulated cows produced a significantly higher number of oocytes and oocyte yield per cow than the non-stimulated ones. A similar observation on the oocyte yield was reported by Patil et al. (Reference Patil, Hadiya, Layek, Gorani, Raj, Karuppanasamy and Gupta2022).
Quality of USG and probe characteristics have a significant impact on the recovery of oocytes (Bols et al., Reference Bols, Vanholder, Leroy, Aerts and Van Soom2003). The higher oocyte recovery may be due to the fact that the OPU was performed at the follicular wave emergence on day 2 after the oestrus and that too in the absence of the dominant follicle and corpus luteum during the early luteal phase. Similar findings were also reported by Viana et al. (Reference Viana, Palhao, Siqueira, Fonseca and Camargo2010) and Hadimani et al. (Reference Hadimani, Khillare, Amle, Meshram, Rangnekar, Mali and Nimbalkar2022). Further, OPU was performed at 85–90 mmHg vacuum pressure resulting in a higher recovery rate, also corroborated with the reports of Manik et al. (Reference Manik, Singla and Palta2003) in cattle, used vacuum pressure of 75–100 mmHg. However, Harkal et al. (Reference Harkal, Pawshe, Ingawale, Deshmukh, Ambalkar and Kale2019) recorded a higher oocyte recovery rate when follicular aspiration was done at 100 mm Hg vacuum pressure. The present findings on oocyte recovery are in agreement with the reports of Li et al. (Reference Li, Chen, Pi, Liu and Shi2007) and Hadimani et al. (Reference Hadimani, Khillare, Amle, Meshram, Rangnekar, Mali and Nimbalkar2022) in the non-stimulated cows. However, many workers reported a higher recovery rate than the present study (Nogueira et al., Reference Nogueira, de Souza, Puelker, Giometti, Firetti, Dias and Castilho2021; Krishna et al., Reference Krishna, Rao, Veerabramhaiah, Kumar, Srikanth and Yasaswini2023).
In FSH-stimulated cows, we recorded higher oocyte recovery rate (%) than the reports of Ongaratto et al. (Reference Ongaratto, Cedeño, Rodriguez-Villamil, Tríbulo and Bó2020). However, similar observations were recorded by Oliveira et al. (Reference Oliveira, Sanches, Seddon, Veras, Lima, Monteiro and Sartori2016) and de Morae s et al. (Reference de Moraes, Adona, Guemra, De Bem and dos Santos Miranda2019). Vieira et al. (Reference Vieira, Rodrigues, Netto, Guerreiro, Silveira, Moreira and Baruselli2014, Reference Vieira, Rodrigues, Netto, Guerreiro, Silveira, Freitas and Baruselli2016) reported that treatment of gonadotropin hormone for 2 days results higher number of COCs and embryos and FSH deprivation for about 36–48 h prior to OPU is recommended to recover more oocytes possessing optimal developmental capacity.
The finding indicated that FSH stimulation of Tharparkar cows resulted in a higher number of oocytes recovery with a concomitant increase in the number of oocytes with a diameter ≥120 µm. Some reports marked the importance of oocyte diameter with its developmental competence (Bhardwaj et al., Reference Bhardwaj, Ansari, Pandey, Parmar, Chandra, Kumar and Sharma2016; Fair, Reference Fair2018). Oocytes retrieved from follicles with larger diameters show better developmental competence as compared to oocytes recovered from smaller follicles and vice-versa (Bagg et al., Reference Bagg, Nottle, Armstrong and Grupen2007). Further, studies also identified the differences between competent and non-competent oocytes based on oocyte diameter (Bessa et al., Reference Bessa, Nishimura, Franco and Dode2013). Yang et al. (Reference Yang, Kanno, Huang, Kang, Yanagawa and Nagano2016) observed better maturation rate and developmental competence for oocytes having >120 µm size and on the other side, Aguila et al. (Reference Aguila, Treulen, Therrien, Felmer, Valdivia and Smith2020) reported that the selection of follicles between 6–10 mm with oocyte diameters ranging between 115–130 µm has the greater potential to optimize developmental competence. In another study, Otoi et al. (Reference Otoi, Yamamoto, Koyama, Tachikawa and Suzuki1997) reported that there is a clear relationship between oocyte diameter and developmental competence in terms of meiotic maturation and embryo yield following IVF procedures. Fair et al. (Reference Fair, Hyttel and Greve1995) reported that the rates of resumption of meiosis I to M-II were higher for oocytes having a size >110 µm.
The results suggested that ovarian stimulation with FSH had a significant impact on oocyte quality as well as quantity including a higher percentage of Grade A oocytes. Gilchrist et al. (Reference Gilchrist, Ritter, Cranfield, Jeffery, Amato, Scott and Ritvos2004) stated that COCs are essential for oocyte development and maturation. Further, Matoba et al. (Reference Matoba, Bender, Fahey, Mamo, Brennan, Lonergan and Fair2014) reported morphological evaluation of COCs for identifying good-quality oocytes as a non-invasive technique for selecting the most competent oocytes. The percentage of quality oocytes in the present study is higher than that of most of the reports (Saini et al., Reference Saini, Singh, Shah, Singh, Kaushik, Manik and Chauhan2015; Landeo et al., Reference Landeo, Zuñiga, Gastelu, Artica, Ruiz, Silva and Ratto2022). However, Sendag et al. (Reference Sendag, Cetin, Alan, Hadeler and Niemann2008) reported higher quality of oocytes in Holstein cows collected through OPU. The higher number of quality oocytes in the present study was due to FSH treatment before OPU which produces more Grade A oocytes as FSH after binding with the FSH receptor modifies follicular cells and increases the number of large (>6 mm) size follicles, oocyte competency and cumulus expansion. The present findings are in agreement with Cavalieri et al. (Reference Cavalieri, Morotti, Seneda, Colombo, Andreazzi, Emanuelli and Rigolon2018) and Ongaratto et al. (Reference Ongaratto, Cedeño, Rodriguez-Villamil, Tríbulo and Bó2020) who also reported a similar trend in the quality of oocytes collected from Bos indicus and Angus cows, respectively.
The study suggests that FSH stimulation had a significant impact notably on the total number of oocytes recovered but also on the percentages of BCB+VE in Tharparkar cows (P <0.05). The BCB staining test is a proven non-invasive and simple method for selecting developmentally competent oocytes and it evaluates glucose-6-phosphate dehydrogenase activity (Bhardwaj et al., Reference Bhardwaj, Ansari, Pandey, Parmar, Chandra, Kumar and Sharma2016) and is employed successfully in cattle (Bhojwani et al., Reference Bhojwani, Alm, Torner, Kanitz and Poehland2007). BCB+ve oocytes showed greater developmental potential, maturation and more blastocyst formation (Silva-Santos et al., Reference Silva-Santos, Santos, Koetz Júnior, Morotti, Siloto, Marcantonio and Seneda2014).
A higher percentage of BCB+ve oocytes in the present study may be due to coasting period of 36–48 h which further helped in the proper growth of the oocytes. On the other hand, Silva-Santos et al. (Reference Silva-Santos, Santos, Koetz Júnior, Morotti, Siloto, Marcantonio and Seneda2014) and Castaneda et al. (Reference Castaneda, Kaye, Pantaleon, Phillips, Norman, Fry and Michael2013) have reported almost comparable quality BCB+ve oocytes in bovine. Castaneda et al. (Reference Castaneda, Kaye, Pantaleon, Phillips, Norman, Fry and Michael2013) observed that the higher lipid content of BCB+ve bovine oocytes might be associated with their better developmental competence. Also, it can be used efficiently to enhance the outcome of IVEP before the maturation of oocytes (Alm et al., Reference Alm, Torner, Lohrke, Viergutz, Ghoneim and Kanitz2005). It is worth to mention that we could not found any literature on BCB staining of oocytes recovered through OPU. To date, the oocyte quality assessment is mostly based on morphology (Wang and Sun, Reference Wang and Sun2006).
GDF9 and BMP-15 also known as oocyte secreted factors, belong to the transforming growth factor-β family and have a potential value as non-invasive markers for determining oocyte developmental potential (Yoshino et al., Reference Yoshino, McMahon, Sharma and Shimasaki2006; Su et al., Reference Su, Sugiura, Li, Wigglesworth, Matzuk and Eppig2010). Several studies in cattle reported higher expression of GDF9 and BMP15 in the ovary which play pivotal roles during folliculogenesis, cumulus expansion, decreasing apoptosis of COCs, oocyte maturation, oocyte developmental competence and thus good quality oocytes (Gode et al., Reference Gode, Gulekli, Dogan, Korhan, Dogan, Bige and Atabey2011; Kussano et al., Reference Kussano, Leme, Guimarães, Franco and Dode2016).
The higher expression of GDF9 and BMP15 genes in FSH-stimulated Tharparkar cows suggests the positive effect of FSH stimulation on the transcriptional activity of these genes. These findings imply a potential role of FSH in enhancing ovarian function and follicular development as both GDF9 and BMP15 are the key genes involved in oocyte development including maturation (Gilchrist et al., Reference Gilchrist, Ritter, Cranfield, Jeffery, Amato, Scott and Ritvos2004). The results of the present study provide valuable insights into the molecular mechanisms underlying the response to FSH stimulation in the context of ovarian function suggesting that FSH may play a key role in modulating the expression of GDF9 and BMP15 genes and thus influencing oocyte quality and reproductive outcomes (Pfeffer et al., Reference Pfeffer, Sisco, Donnison, Somers and Smith2007; Sirait et al., Reference Sirait, Wiweko, Jusuf, Iftitah and Muharam2021). Further, Hussein et al. (Reference Hussein, Thompson and Gilchrist2006) reported that BMP15 and GDF9 enhance oocyte developmental competence by regulating the COCs microenvironment and oocyte developmental programming. Increased BMP15 and GDF9 expression stimulates FSH receptor expression in pre-antral follicles and the level of FSH receptors in granulosa cells which in turn leads to better maturation, subsequent embryonic development and establish a viable full-term pregnancy in mammals (Rajput et al., Reference Rajput, Lee, Zhenhua, Di, Folger and Smith2014). Sugimura et al. (Reference Sugimura, Kobayashi, Okae, Yamanouchi, Matsuda, Kojima and Gilchrist2017) highlighted that BMP15 and GDF9 stimulate gene expression that enables cumulus expansion via epidermal growth factor signalling and further reported that in the event of FSH-priming, there is better cell-to-cell communication, enhanced follicular growth which allows rescuing some of the follicles from atresia and maintain healthy status by activating anti-inflammatory cascade, transport of nutritional molecules from cumulus cells to oocytes towards final oocyte maturation and establishing proper ovulation cascade. The higher mRNA expression of BMP-15 and GDF9 in the present study might be responsible for the higher follicular growth, oocyte diameter, recovery as well as quality. These findings have been supported by the findings of Chu et al. (Reference Chu, Dufort and Sirard2012) and Nivet et al. (Reference Nivet, Bunel, Labrecque, Belanger, Vigneault, Blondin and Sirard2012). Saini et al. (Reference Saini, Singh, Shah, Singh, Kaushik, Manik and Chauhan2015) reported higher expression of GDF9 and BMP15 and concomitantly greater cleavage rate of Grade A and B than C and D Grade COCs.
PCNA protein is an amazing biomarker of cellular proliferation expressed at higher levels in oocytes recovered from more mature follicles ranging between 6–10 mm (Robker & Richards, Reference Robker and Richards1998). Various researchers explained the role of the PCNA gene during folliculogenesis in the oocytes (Oktay et al., Reference Oktay, Schenken and Nelson1995; Wood and Shivji, Reference Wood and Shivji1997).
The possible explanation could be that due to more number of cumulus cell layers results into more mRNA expression in good-quality oocytes. In the present study, we also observed relatively higher expression of the PCNA gene in the oocytes of FSH-stimulated cows, as comparatively larger-sized follicles were recorded (≥6 mm) than the non-stimulated group. However, the significance of the PCNA expression in the oocyte is still unknown and yet to be elucidated. Further, it has been speculated that though no new DNA synthesis takes place in the growing oocyte, it is possible that DNA polymerases are activated to repair potential damage to the genetic material in the oocytes selected to grow (Oktay et al., Reference Oktay, Schenken and Nelson1995). The presence of relatively higher expression of the PCNA gene in oocytes of FSH stimulated group suggests a role for this protein in folliculogenesis.
BCL-2 and BAX are the molecular markers used to assess the developmental competence of oocytes recovered from cattle (Opiela et al., Reference Opiela, Kątska-Książkiewicz, Lipiński, Słomski, Bzowska and Ryńska2008). The different expression levels of BCL-2 and BAX genes in the FSH-stimulated group have yielded interesting results as both pivotal genes are involved in the regulation of apoptosis.
The increase in the expression of BCL-2 and a concomitant decrease in BAX gene transcript expression suggests that FSH stimulation plays an important role in up-regulating the anti-apoptotic pathway. It has been found that the BCL-2 protein prevents apoptosis induced by a variety of stimuli and maintains cell survival by influencing the release of cytochrome C from mitochondria rather than by altering proliferation (Yang et al., Reference Yang, Liu, Bhalla, Kim, Ibrado, Cai and Wang1997). On the other hand, BAX identified by co-immunoprecipitation with the BCL-2 protein, is the first pro-apoptotic homolog. When BAX is overexpressed in cells, apoptotic death is accelerated.
The finding of the present study is supported by Saini et al. (Reference Saini, Singh, Shah, Singh, Kaushik, Manik and Chauhan2015), Melo et al. (Reference Melo, Cordeiro, Pellegrino, Wei, Daye, Nishimura and Dode2017) and Khurchabilig et al. (Reference Khurchabilig, Sato, Ashibe, Hara, Fukumori and Nagao2020) that BCL-2 and BAX are the molecular markers used to assess the developmental competence of oocytes and showed upregulation in good quality oocytes and downregulation in poor quality oocytes in cattle. However, the decrease in BAX expression indicates that FSH stimulation may exert an inhibitory effect on the pro-apoptotic pathway by rescuing some of the follicles from atresia and progressing towards growth phase. BAX gene is involved in promoting cell death and its lower expression in the FSH-stimulated group implies a potential mechanism by which FSH may suppress apoptosis and support cell survival. In the present study, we observed that the non-stimulated Tharparkar cows oocytes had relatively lower expression of GDF9, BMP15 and BCL-2 genes which may be due to less proportion of good quality oocytes i.e. Grade A and Grade B type of oocytes. Irrespective of stimulation, several researchers (Saini et al., Reference Saini, Singh, Shah, Singh, Kaushik, Manik and Chauhan2015, Boldura et al., Reference Boldura, Nica, Milovanov, Popescu, Marc Zarcula, Bonca and Tulcan2016) reported that in the event of higher proportion in good quality oocytes (more Grade A and Grade B), there was a relatively higher expression of BCL-2 having greater developmental competence, however, expression of BAX being pro-apoptotic gene was more in poor quality oocytes (Grade C and D) with poor developmental competence. Saini et al. (Reference Saini, Singh, Shah, Singh, Kaushik, Manik and Chauhan2015) reported Grade A and B COCs showed greater expression of BCL-2 and higher developmental competence of oocytes than Grade C and D COCs during, before and after maturation stages. However, De Bem et al. (Reference De Bem, Adona, Bressan, Mesquita, Chiaratti, Meirelles and Leal2014) reported that the developmental competence of an oocyte is dependent on numerous variables and multiple factors must be considered to determine the quality of an oocyte.
In conclusion, from this study, significant changes in morphological and molecular parameters associated with oocyte competence have been recorded. The partial stimulation of ovaries resulted in higher number of follicles ≥6 mm, more quality oocytes recovery. Stimulated group yielded more grade A oocytes, higher percentage of oocytes with diameter of ≥120 µm and more BCB+VE. Further, the relative mRNA expression profile of oocyte-secreted factors viz. GDF9, BMP15, PCNA (Proliferation indicator) and cells apoptotic regulators like BCL-2 was higher and BAX was lower in the FSH-stimulated cow. These results indicated that FSH stimulation before OPU in Bos indicus cows has a significant impact on follicle size, quality oocyte recovery which helps in proper embryo development and successful pregnancy.
Acknowledgements
The authors would like to thank the Director and In charge, Cattle and Buffalo Farm, ICAR-IVRI, Izatnagar, India for facilitating the study.
Funding
This research was supported by Institute-funded project entitled ‘Quality embryo and superior calves production using multiple ovulation and embryo transfer and OPU-IVF-ET platform in Indigenus cattle’ (Project No. 7-31/AR/2020-21).
Competing interests
The authors declare that there is no conflict of interest in publishing the article.
Ethical standards
All experimental procedures were approved by the Institute Animal Ethics Committee of ICAR-IVRI, Izatnagar, U.P. India (26-3/2020-21/JD(R)/IAEC).







