Early hominins were not limited to particular sites or localities in a paleontological or archeological sense, but lived and died in complex and dynamic landscapes and ecosystems of which we have partial, incomplete records. The fossil evidence of early hominin paleoenvironments is always limited, sometimes providing brief snapshots of small areas, other times affording very coarse chronological and spatial resolution over large distances. Taphonomic conditions typically vary within any one locality over time, and from one locality to another. And yet, it is these partial and biased records that we use to build an understanding of the forces that have shaped our evolution.
We think that to gain a better understanding of our evolution and its ecological context it is necessary to take a broad view. As in the pioneering volume by Clark Howell and François Bourlière, African Ecology and Human Evolution (1963), here we aim to consider the whole continent of Africa during the time when our lineage emerged and dispersed across the continent. The focus of this volume is on the paleoenvironmental conditions and paleoecological relationships that are likely to have shaped the evolution of our ancestors as well as closely related species during the past 7 million years. We aim to discuss regions with a rich fossil record as well as regions with a sparse record, from north to south and from east to west (Figure 1.1).
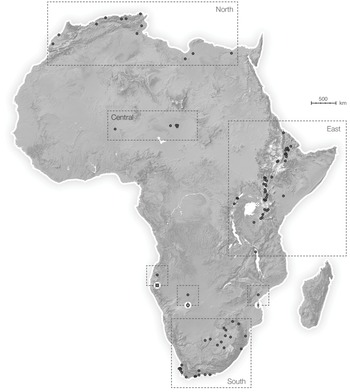
Figure 1.1 Continental map of Africa, with dashed boxes showing the geographic extent of the smaller regional maps. Individual sites (or site regions) are indicated with open circles and are labelled in the regional maps and discussed in the text.
This volume bears witness to decades of painstaking work by numerous researchers at hundreds of different sites. The research presented in the following chapters is necessarily interdisciplinary and builds on the efforts of scholars like Jean de Heinzelin, Bill Bishop, Maurice Taieb, Frank Brown, and Richard Hay, who, among others, laid a solid geological foundation for studies of hominin paleoenvironments. Likewise, the taphonomic approaches of Kay Behrensmeyer in East Africa, among others, continue to shape paleoecological studies across the continent.
This volume is organized into four main parts. In Part I, we present a review of key methods of paleoecological reconstruction, discuss biases in the hominin fossil record, and provide two synthetic chapters on the paleoenvironments of the continent. Part II is focused on southern Africa (Figure 1.2), with an overview chapter followed by site-specific contributions on Makapansgat, Kromdraai, Cooper’s Cave, Sterkfontein, Swartkrans, Cornelia-Uitzoek, Taung, and Wonderwerk Cave.
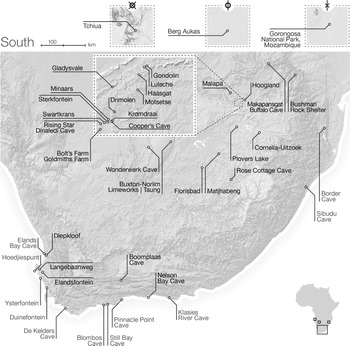
Figure 1.2 Regional map of southern Africa, including site regions in Angola, Namibia, and Mozambique. The very dense cluster of sites in the Cradle of Humankind World Heritage Site are indicated in the dashed inset box.
Part III begins with an overview of eastern, central, and western African paleoenvironments during the late Miocene to the early Pleistocene, followed by site-specific chapters on Buia, Gona, Hadar, Dikika, Galili, Melka Kunture, Konso, the lower Omo Valley, West Turkana, East Turkana, Tugen Hills, the Homa Peninsula, Olorgesailie, Peninj, Olduvai, Laetoli, and the Karonga Basin (Figure 1.3a, 1.3b). Part IV provides a discussion of North African paleoenvironments in the Pliocene and Pleistocene, with site-specific contributions on Ahl al Oughlam, Tighenif, Thomas Quarry, and Oulad Hamida Quarry (Figure 1.4).
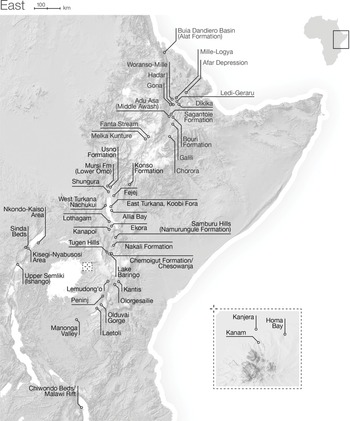
Figure 1.3a The dense clustering of sites in eastern Africa is closely associated with the East African Rift System, which contains two main rift branches: the Eastern (Gregory) Branch, which extends from the Afar region in Ethiopia through Kenya and Tanzania, and the Western (Albertine) Branch from Uganda and the DRC to Malawi and Mozambique.

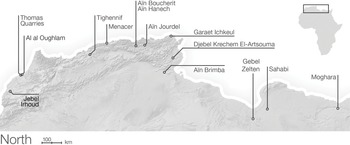
Figure 1.4 The important sites of northern Africa, including Jebel Irhoud, which has yielded the oldest remains of Homo sapiens.
As currently understood, hominin evolution spans the period from nearly 7 Ma (million years) to the present (Figure 1.5). This time frame is based on the earliest records of possible hominins, including the species Ardipithecus kadabba (Reference WoldeGabriel, Haile-Selassie and RenneHaile-Selassie, 2001; Reference Haile-Selassie, Suwa and WhiteHaile-Selassie et al., 2004a, Reference Haile-Selassie, White, Bernor, Rook, Vrba, Haile-Selassie and WoldeGabriel2009a), Orrorin tugenensis (Reference Senut, Pickford and GommerySenut et al., 2001), and Sahelanthropus tchadensis (Reference Brunet, Guy and PilbeamBrunet et al., 2002). Thus, current evidence indicates that the earliest hominins lived in eastern and central Africa. The earliest evidence of the genus Australopithecus dates to about 4.2 Ma, and by about 3.4 Ma there is some evidence of phylogenetic diversification with a few species of Australopithecus occurring in eastern, central, and possibly southern Africa (Reference Haile-Selassie, Mellilo and SuHaile-Selassie et al., 2016a). Homo and Paranthropus appear in the fossil records of eastern Africa toward the end of the Pliocene, about 2.8–2.7 Ma (Reference Wood and LeakeyWood and Leakey, 2011; Reference Villmoare, Kimbel and SeyoumVillmoare et al., 2015), and diversify during the early Pleistocene (Reference Antón, Potts and AielloAntón et al., 2014). By the end of the early Pleistocene, ~0.8 Ma, only species of the genus Homo remain in Africa and elsewhere (Reference RightmireRightmire, 2013; Reference Schroeder, Scott and GarvinSchroeder et al., 2017).
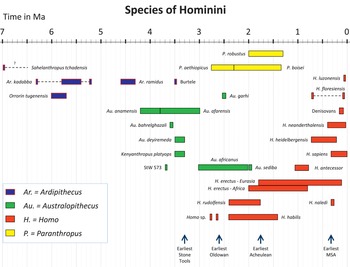
Figure 1.5 Chronology of hominin species. There are alternative interpretations to the hominin taxa depicted here.
Hominid evolution takes place in the late Cenozoic during a time of important climatic, tectonic, and environmental changes. After the Mid-Miocene Climatic Optimum, 17–15 Ma, high-latitude global temperatures as measured by oxygen isotopes in marine Foraminifera began to decline (Figure 1.6; Reference Zachos, Pagani, Sloan, Thomas and BillupsZachos et al., 2001). This cooling trend resulted in the establishment of the East Antarctica ice sheets in the middle Miocene (Reference Lewis, Marchant, Ashworth, Hemming and MachlusLewis et al., 2007) and the West Antarctic ice sheets in the late Miocene (Reference Zachos, Pagani, Sloan, Thomas and BillupsZachos et al., 2001). The late Miocene climatic cooling culminated in ephemeral glaciation in the northern hemisphere between 6 and 5.5 Ma (Reference Holbourn, Kuhnt and ClemensHolbourn et al., 2018) coincident with the Messinian salinity crisis (Reference Krijgsman, Hilgen, Raffi, Sierro and WilsonKrijgsman et al., 1999; Reference Garcia-Castellanos and VillaseñorGarcia-Castellanos and Villaseñor, 2011). The early Pliocene (4.5–3.0 Ma) saw a return to warmer temperatures (Reference Wara, Ravelo and DelaneyWara et al., 2005; Reference Ravelo, Dekens and McCarthyRavelo et al., 2006; Reference Fedorov, Brierley and LawrenceFedorov et al., 2013) with generally wetter conditions in much of Africa (Reference Feakins, deMenocal, Werdelin and SandersFeakins and deMenocal, 2010; Reference Salzmann, Williams and HaywoodSalzmann et al., 2011). The late Pliocene and Pleistocene was a time of further global cooling, with the onset of Northern Hemisphere glaciation between 3 and 2.6 Ma (Reference Ravelo, Andreasen, Lyle, Olivarez Lyle and WaraRavelo et al., 2004), and increasing amplitude of climatic oscillations during the Pleistocene ice ages (Reference Raymo and HuybersRaymo and Huybers, 2008). The mid-Pleistocene transition at ~0.95 Ma was marked by increased ice volume in northern latitudes and an increase in the amplitude of climatic oscillations (Reference Larrasoaña, Roberts, Rohling, Winklhofer and WehausenLarrasoaña et al., 2003). Dust records off the coast of Africa indicate that there were peaks of aridity at 2.8, 1.7, and 1.0 Ma (Reference deMenocaldeMenocal, 1995). Alongside these major trends, climate cycles resulting from orbital forcing, i.e., changes in the Earth’s orbital precession, axial obliquity, and orbital eccentricity (Milankovitch cycles; Reference Hays, Imbrie and ShackletonHays et al., 1976; Reference ShackletonShackleton, 2000; Reference Philander and FedorovPhilander and Fedorov, 2003; Reference Lisiecki and RaymoLisiecki and Raymo, 2007; Reference Bajo, Drysdale and WoodheadBajo et al., 2020), may have played an important role in environmental and ecological changes in Africa (Reference Kingston, Deino, Edgar and HillKingston et al., 2007; Reference Lupien, Russell and YostLupien et al., 2019). Cycles of ~21 ky (orbital precession) were dominant during the Miocene and most of the Pliocene, shifting to 41 ky (obliquity) dominant cyclicity at ~2.8 Ma, and 100–400 ky cycles (eccentricity modulating orbital precession) after 1 Ma (Reference Tiedemann, Sarnthein and ShackletonTiedemann et al., 1994; Reference deMenocaldeMenocal, 1995). Orbital forcing interacts with monsoon cycles to modulate levels and distribution of precipitation across eastern Africa, where the Intertropical Convergence Zone plays a key role (Reference Feakins, deMenocal, Werdelin and SandersFeakins and deMenocal, 2010). Some of the high-eccentricity intervals coincided with periods of high precipitation in eastern Africa and expansion of rift valley lakes (Reference Trauth, Maslin, Deino and StreckerTrauth et al., 2005, Reference Trauth, Larrasoaña and Mudelsee2009). However, it remains a major challenge to relate the fine-scale resolution of many climatic records from the oceans to the coarser and more discontinuous records of hominins and their environments on land (Reference BehrensmeyerBehrensmeyer, 2006).
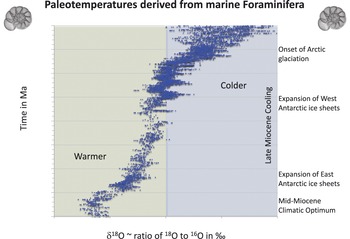
Figure 1.6 Paleotemperatures derived from benthic Foraminifera with δ18O values in ‰ on the horizontal axis and time in Ma (million years) on the vertical axis. Graph drawn with data from the Deep Sea Drilling Project and the Ocean Drilling Project summarized in Reference Zachos, Pagani, Sloan, Thomas and BillupsZachos et al. (2001). This graph provides a view of global temperatures during the past 16 million years as a general background to understanding evolution in Africa.
The climatic changes discussed above are manifested in Africa through their influence on precipitation and vegetation. In modern African savannas, the proportion of woody vs. grassy vegetation is primarily determined by mean annual precipitation. Regions which receive <650 mm of rain per year tend to have constrained woody cover but with significant grasses (Reference Sankaran, Hanan and ScholesSankaran et al., 2005). Regions with more than 650 mm of rainfall have woody cover that tends to become dominant, unless constrained by fires or herbivores, with megaherbivores playing an important role in opening up wooded areas (Reference WesternWestern, 1989). It is not only the amount of rainfall that’s important in structuring vegetation: the seasonal distribution of rainfall and the severity and length of dry seasons also play a key role (Reference ShorrocksShorrocks, 2007). Furthermore, topography and soil types modify the effects of rainfall.
In our environmental descriptions, we rely on Frank White’s African main vegetation types (Reference White and HannusWhite, 1983). Forests consist of continuous stands of trees with interdigitating crowns in multiple layers, frequently with lianas. Canopy height ranges from 10 to 50 m or more. The ground layer may include shrubs, bryophytes, and localized grasses. Rainforests are most widespread in the Guineo-Congolian region, with drier forests in this region classified as semi-evergreen. Dry forests, which may experience several months of dry season, are most common in the Zambezian and Sudanian regions. Riparian or gallery forests bordering fluvial channels and floodplains provide shade, refuge, and arboreal resources in areas that may otherwise have sparse arboreal vegetation. Afromontane forests typically occur at elevations above 1500 m and are composed of shorter trees than in lowland rainforests. Woodlands consist of widely spaced stands of trees with canopies covering at least 40 percent of the ground surface. Canopy height ranges from 8 to 20 m, and heliophilous grasses dominate the ground layer. Deciduous and semideciduous woodlands are common in the Sudanian and Zambezian regions. Bushland consists of areas dominated by woody plants usually 3–7 m in height, and covering at least 40 percent of the ground surface. Bushland is common in the Somalia–Masai region. Wooded grasslands are areas in which woody plants cover from 10 percent to 40 percent of the surface area, with grasses predominating. Grasslands are areas covered primarily by grasses with up to 10 percent cover of woody plants. Sedges (Cyperaceae) may be very common in edaphic grasslands associated with waterlogged conditions, which are widespread in the Sudanian, Zambezian, Somalia–Masai, and coastal regions. The term savanna refers to tropical and subtropical grasslands with scattered bushes and trees where the dominant grasses tend to be C4 plants (Reference ShorrocksShorrocks, 2007). These vegetation types usually grade into one another and may be interdigitated in complex ways depending on various factors (topography, hydrology, and soils).
We use the term environment to refer broadly to the supporting matrices of life that surround living organisms (Reference CalowCalow, 1999). An environment may be more or less heterogeneous depending on the organism sensing it (Reference Begon, Townsend and HarperBegon et al., 2006). Paleoenvironments are the inferred matrices of life surrounding organisms in the past. The term habitat refers to the place where a species or population lives. Species favor some habitats over others, evidenced by their abundance and distribution. An assemblage of species that interact with one another in a given area is called a community (Reference Levin, Carpenter and GodfrayLevin et al., 2009). Fossil assemblages rarely constitute past communities in an ecological sense. Time-averaging, bone durability, and transport tend to mix individuals from different communities at different times (Reference Behrensmeyer, Kidwell and GastaldoBehrensmeyer et al., 2000). On a broader scale, the term ecosystem refers to the integrated biological and physical components of a landscape with a range of community types (Reference Levin, Carpenter and GodfrayLevin et al., 2009).
The chapters that follow provide a rich source of primary and synthetic data on hominin paleoenvironments and paleoecology in Africa, with most contributions focusing on the Pliocene and early Pleistocene, but also with contributions spanning from the late Miocene to the late Pleistocene and Holocene. Several chapters present primary data on the mammalian fauna, but also discuss other available paleoenvironmental and paleoecological sources of evidence. We hope that these contributions will generate further synthetic work, exploration of unknown areas, and generate new ideas and approaches to the study of our African origins.